
Arbovirus Detection in Aedes aegypti Mosquitoes in Manabí, Ecuador
 , and
, and
Abstract
1. Introduction
2. Material and Methods
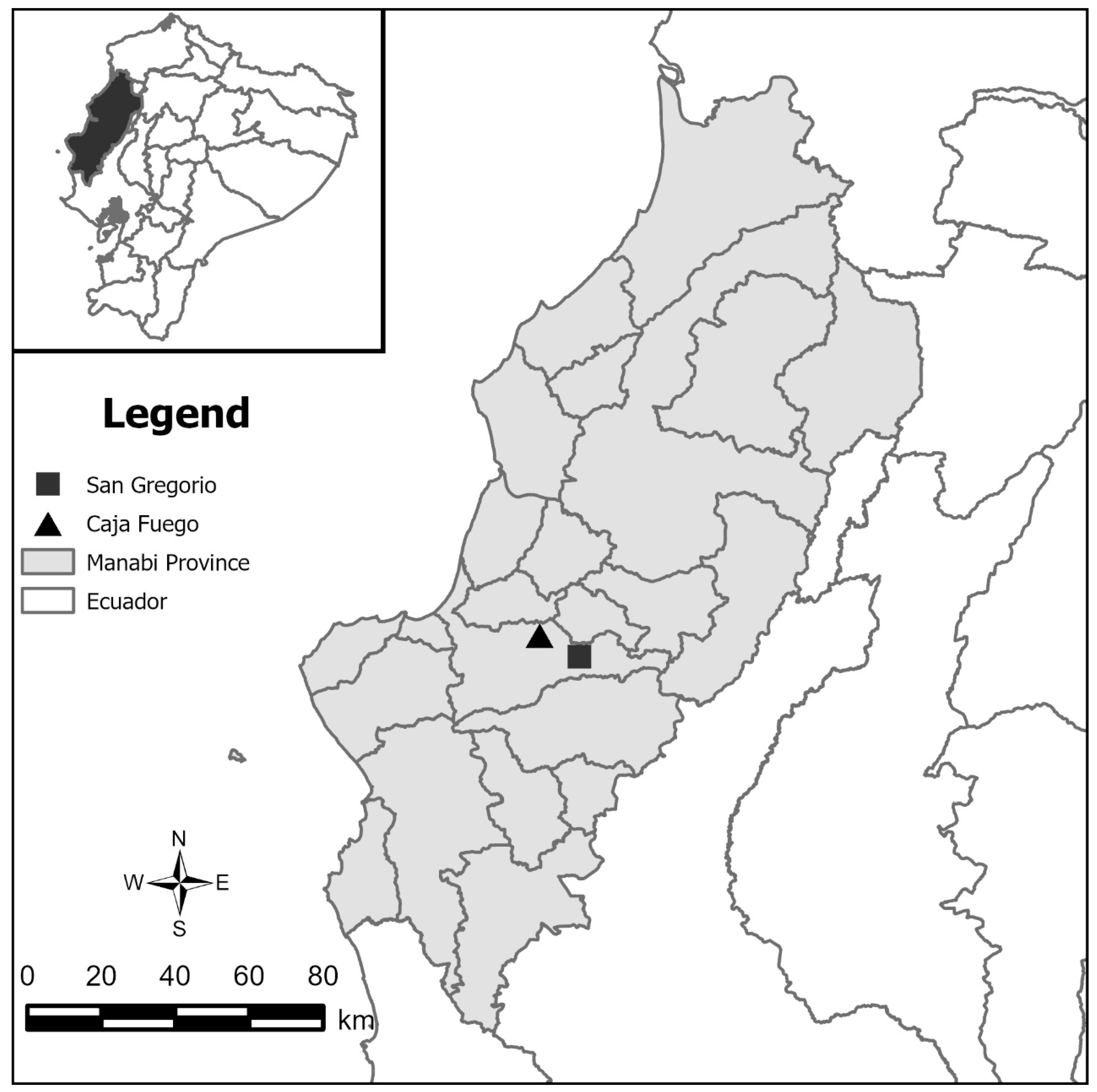
2.1. Study Area
2.2. Mosquito Collection
2.3. RNA Extraction in Pool
2.4. cDNA Synthesis
2.5. Positive and Negative Controls
2.6. End-Point PCR
2.7. Real-Time PCR and Sanger Sequencing
2.8. Bioinformatic Analysis
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hernandez, R.; Brown, D.T.; Paredes, A. Structural differences observed in arboviruses of the alphavirus and flavivirus genera. Adv. Virol. 2014, 2014, 259382. [Google Scholar] [CrossRef] [PubMed]
- Soni, S.; Gill, V.J.S.; Singh, J.; Chhabra, J.; Gill, G.J.S.; Bakshi, R. Dengue, chikungunya, and Zika: The causes and threats of emerging and re-emerging arboviral diseases. Cureus 2023, 15, e41717. [Google Scholar] [CrossRef]
- Velandia-Romero, M.; Olano, V.; Coronel-Ruiz, C.; Cabezas, L.; Calderón-Peláez, M.; Castellanos, J.; Matiz, M. Detección del virus del dengue en larvas y pupas de Aedes aegypti recolectadas en áreas rurales del municipio de Anapoima, Cundinamarca, Colombia. Biomédica 2017, 37, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Ryan, S.J.; Carlson, C.J.; Mordecai, E.A.; Johnson, L.R. Global expansion and redistribution of Aedes-borne virus transmission risk with climate change. PLoS Negl. Trop. Dis. 2019, 13, e0007213. [Google Scholar] [CrossRef] [PubMed]
- Campbell-Lendrum, D.; Manga, L.; Bagayoko, M.; Sommerfeld, J. Climate change and vector-borne diseases: What are the implications for public health research and policy? Philos. Trans. R. Soc. Biol. Sci. 2015, 370, 20130552. [Google Scholar] [CrossRef]
- Santiago, G.A.; Vázquez, J.; Courtney, S.; Matías, K.Y.; Andersen, L.E.; Colón, C.; Butler, A.E.; Roulo, R.; Bowzard, J.; Villanueva, J.M.; et al. Performance of the Trioplex real-time RT-PCR assay for detection of Zika, dengue, and chikungunya viruses. Nat. Commun. 2018, 9, 1391. [Google Scholar] [CrossRef]
- Reinhold, J.M.; Lazzari, C.R.; Lahondère, C. Effects of the environmental temperature on Aedes aegypti and Aedes albopictus mosquitoes: A Review. Insects 2018, 9, 158. [Google Scholar] [CrossRef]
- Silva, N.M.; Santos, N.C.; Martins, I.C. Dengue and Zika viruses: Epidemiological history, potential therapies, and promising vaccines. Trop. Med. Infect. Dis. 2020, 5, 150. [Google Scholar] [CrossRef]
- World Health Organization. Disease Outbreak News: Annual Compendium 2022; World Health Organization: Geneva, Switzerland, 2024; Available online: https://www.who.int/es/emergencies/disease-outbreak-news/item/2023-DON448 (accessed on 14 December 2024).
- Khan, M.B.; Yang, Z.S.; Lin, C.Y.; Hsu, M.C.; Urbina, A.N.; Assavalapsakul, W.; Wang, W.H.; Chen, Y.H.; Wang, S.F. Dengue overview: An updated systemic review. J. Infect. Public Health 2023, 16, 1625–1642. [Google Scholar] [CrossRef]
- Chen, Y.; Li, N.; Lourenço, J.; Wang, L.; Cazelles, B.; Dong, L.; Li, B.; Liu, Y.; Jit, M.; Bosse, N.I.; et al. Measuring the effects of COVID-19-related disruption on dengue transmission in southeast Asia and Latin America: A statistical modelling study. Lancet Infect. Dis. 2022, 22, 657–667. [Google Scholar] [CrossRef]
- Pan American Health Organization. Epidemiological Update—Dengue, Chikungunya and Zika. 2023. Available online: https://www.paho.org/en/documents/epidemiological-update-dengue-chikungunya-and-zika-10-june-2023 (accessed on 10 September 2023).
- MSP. Enfermedades Transmitidas por Vectores Dengue Ecuador SE 01-14/2020. Available online: https://www.salud.gob.ec/wp-content/uploads/2020/04/DENGUE-SE_14_2020_GACETA.pdf (accessed on 10 September 2023).
- Valero-Cedeño, N.; Baque-Arteaga, K.; Calderón-Pico, A.; Caiza-Defaz, C.; Escobar-Rivera, M. Prevalencia de zika y chikungunya en los cantones de Jipijapa y Puerto López de la Provincia de Manabí, Ecuador. 2015–2020. Polo Del Conoc. 2020, 46, 968–981. Available online: https://polodelconocimiento.com/ojs/index.php/es/article/view/2388/4877 (accessed on 22 November 2024).
- MSP. Enfermedades Transmitidas por Vectores Dengue Ecuador SE 03-1/2023. Available online: https://www.salud.gob.ec/wp-content/uploads/2023/02/Gaceta-SE-3_2023.pdf (accessed on 10 September 2023).
- Talbot, B.; Sander, B.; Cevallos, V.; González, C.; Benítez, D.; Carissimo, C.; Carrasquilla Ferro, M.C.; Gauto, N.; Litwiñiuk, S.; López, K.; et al. Determinants of Aedes mosquito density as an indicator of arbovirus transmission risk in three sites affected by co-circulation of globally spreading arboviruses in Colombia, Ecuador and Argentina. Parasites Vectors 2021, 14, 482. [Google Scholar] [CrossRef]
- López-Rosero, A.; Sippy, R.; Stewart-Ibarra, A.M.; Ryan, S.J.; Mordecai, E.; Heras, F.; Beltrán, E.; Costales, J.A.; Neira, M. High prevalence of Zika virus infection in populations of Aedes aegypti from South-western Ecuador. PLoS Negl. Trop. Dis. 2024, 18, e0011908. [Google Scholar] [CrossRef] [PubMed]
- Ryan, S.J.; Mundis, S.J.; Aguirre, A.; Lippi, C.A.; Beltrán, E.; Heras, F.; Sanchez, V.; Borbor-Cordova, M.J.; Sippy, R.; Stewart-Ibarra, A.M.; et al. Seasonal and geographic variation in insecticide resistance in Aedes aegypti in southern Ecuador. PLoS. Negl. Trop. Dis. 2019, 13, e0007448. [Google Scholar] [CrossRef] [PubMed]
- Morales, D.; Ponce, P.; Cevallos, V.; Espinosa, P.; Vaca, D.; Quezada, W. Resistance Status of Aedes aegypti to Deltamethrin, Malathion, and Temephos in Ecuador. J. Am. Mosq. Control. Assoc. 2019, 35, 113–122. [Google Scholar] [CrossRef]
- Grijalva, M.J.; Villacís, A.G.; Moncayo, A.L.; Ocaña-Mayorga, S.; Yumiseva, C.A.; Baus, E.G. Distribution of triatomine species in domestic and peridomestic environments in central coastal Ecuador. PLoS Negl. Trop. Dis. 2017, 11, e0005970. [Google Scholar] [CrossRef]
- Black, C.L.; Ocaña, S.; Riner, D.; Costales, J.A.; Lascano, M.S.; Davila, S.; Arcos-Teran, L.; Seed, J.R.; Grijalva, M.J. Household risk factors for Trypanosoma cruzi seropositivity in two geographic regions of Ecuador. J. Parasitol. 2007, 93, 12–16. [Google Scholar] [CrossRef]
- Villacís, A.G.; Marcet, P.L.; Yumiseva, C.A.; Dotson, E.M.; Tibayrenc, M.; Brenière, S.F.; Grijalva, M.J. Pioneer study of population genetics of Rhodnius ecuadoriensis (Hemiptera: Reduviidae) from the central coast and southern Andean regions of Ecuador. Infect. Genet. Evol. 2017, 53, 116–127. [Google Scholar] [CrossRef]
- Hidalgo Zambrano, R.V.; Milanes, C.B.; Pérez Montero, O.; Mestanza-Ramón, C.; Nexar Bolivar, L.O.; Cobeña Loor, D.; García Flores De Válgaz, R.G.; Cuker, B.A. Sustainable Proposal for a Cultural Heritage Declaration in Ecuador: Vernacular Housing of Portoviejo. Sustainability 2023, 15, 1115. [Google Scholar] [CrossRef]
- Vazquez-Prokopec, G.M.; Galvin, W.A.; Kelly, R.; Kitron, U. A New, Cost-Effective, Battery-Powered Aspirator for Adult Mosquito Collections. J. Med. Entomol. 2009, 6, 1256–1259. [Google Scholar] [CrossRef]
- Kuno, G. Universal diagnostic RT-PCR protocol for arboviruses. J. Virol. Methods 1998, 72, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, M.; Proebster, B.; Kinney, R.M.; Kaaden, O.R. Genus-specific detection of alphaviruses by a semi-nested reverse transcription-polymerase chain reaction. Am. J. Trop. Med. Hyg. 1997, 57, 709–718. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Calisher, C.H.; Gubler, D.J.; Chang, G.J.; Vorndam, A.V. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 545–551. [Google Scholar] [CrossRef]
- Faye, O.; Faye, O.; Dupressoir, A.; Weidmann, M.; Ndiaye, M.; Alpha Sall, A. One-step RT-PCR for detection of Zika virus. J. Clin. Virol. 2008, 43, 96–101. [Google Scholar] [CrossRef]
- BioPerfectus Technologies. Zika Virus Real Time PCR Kit. Available online: https://www.bioperfectus.com/ProductDetail/ZikaVirusRealTimePCRKit (accessed on 18 September 2023).
- Stellwagen, N.C. Electrophoresis of DNA in agarose gels, polyacrylamide gels and in free solution. Electrophoresis 2009, 30, S188–S195. [Google Scholar] [CrossRef]
- Ryan, S.J.; Lippi, C.A.; Stewart-Ibarra, A.M. Mapping geographic and demographic shifts for container breeding mosquito-borne disease transmission suitability in Central and South America in a warming world. PLoS Clim. 2024, 49, e0000312. [Google Scholar] [CrossRef]
- Ciota, A.T. The role of co-infection and swarm dynamics in arbovirus transmission. Virus Res. 2019, 35, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Rückert, C.; Weger-Lucarelli, J.; Garcia-Luna, S.M.; Young, M.C.; Byas, A.D.; Murrieta, R.A.; Fauver, J.R.; Ebel, G.D. Impact of simultaneous exposure to arboviruses on infection and transmission by Aedes aegypti mosquitoes. Nat. Commun. 2017, 8, 15412. [Google Scholar] [CrossRef]
- Hummon, A.B.; Lim, S.R.; Difilippantonio, M.J.; Ried, T. Isolation and solubilization of proteins after TRIzol® extraction of RNA and DNA from patient material following prolonged storage. Biotechniques 2007, 42, 467–472. [Google Scholar] [CrossRef]
- Mhamadi, M.; Mencattelli, G.; Gaye, A.; Ndiaye, E.H.; Sow, A.A.; Faye, M.; Ndione, M.H.D.; Diagne, M.M.; Mhamadi, M.; Faye, O.; et al. Rapid On-Site Detection of Arboviruses by a Direct RT-qPCR Assay. Biosensors 2023, 13, 1035. [Google Scholar] [CrossRef]
- Bos, S.; Gadea, G.; Despres, P. Dengue: A growing threat requiring vaccine development for disease prevention. Pathog. Glob. Health 2018, 112, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Harada, S. RNA isolation from mammalian samples. Curr. Protoc. Mol. Biol. 2013, 103, 4–16. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Primer Pair (Code) | Amplicon | Source | |
|---|---|---|---|
| Flavivirus | MA: 5′-CATGATGGGRAARAGRGARRAG-3′ CFD2: 5′-GTGTCCCAGCCGGCGGTGTCATCAGC-3′ | 260 bp | [25] |
| Alphavirus | M2W: 5′-(CT)AGAGC(AGT)TTTTCGCA(CT)(GC)T(AG)GC(ACT)(AT) cM3W: 3′-ACAT(AG)AAN(GT)GNGTNGT(AG)TC(AG)AANCC(AGT)A(CT)CC | 434 bp | [26] |
| dengue | D1-F: 5′-TCAATATGCTGAAACGCGCGAGAAACCG-3′ D2-R: 5′-TTGCACCAACAGTCAATGTCTTCAGGTTC-3′ | 511 bp | [27] |
| Zika | ZIKVENVF: 5′-GCTGGDGCRGACACHGGRACT-3′ ZIKVENVR: 5′-RTCYACYGCCATYTGGRCTG-3′ | 364 bp | [28] |
| End-Point PCR | RT-PCR Kit (bioPerfectus) | Sanger Sequencing | |
|---|---|---|---|
| Flavivirus | >75% | N/A | N/A |
| dengue | 11 | 0 | 0 |
| Zika | 7 | 0 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilca-Cepeda, A.; López-Rosero, A.; Yumiseva, C.A.; Grijalva, M.J.; Villacís, A.G. Arbovirus Detection in Aedes aegypti Mosquitoes in Manabí, Ecuador. Pathogens 2025, 14, 446. https://doi.org/10.3390/pathogens14050446
Wilca-Cepeda A, López-Rosero A, Yumiseva CA, Grijalva MJ, Villacís AG. Arbovirus Detection in Aedes aegypti Mosquitoes in Manabí, Ecuador. Pathogens. 2025; 14(5):446. https://doi.org/10.3390/pathogens14050446
Chicago/Turabian StyleWilca-Cepeda, Alvaro, Andrea López-Rosero, Cesar A. Yumiseva, Mario J. Grijalva, and Anita G. Villacís. 2025. "Arbovirus Detection in Aedes aegypti Mosquitoes in Manabí, Ecuador" Pathogens 14, no. 5: 446. https://doi.org/10.3390/pathogens14050446
APA StyleWilca-Cepeda, A., López-Rosero, A., Yumiseva, C. A., Grijalva, M. J., & Villacís, A. G. (2025). Arbovirus Detection in Aedes aegypti Mosquitoes in Manabí, Ecuador. Pathogens, 14(5), 446. https://doi.org/10.3390/pathogens14050446