
PCR Detection, Genotyping, and Differentiation of Toxoplasma gondii from Hammondia hammondi Excreted in the Feces of Cats in Poland Between 2020 and 2024

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
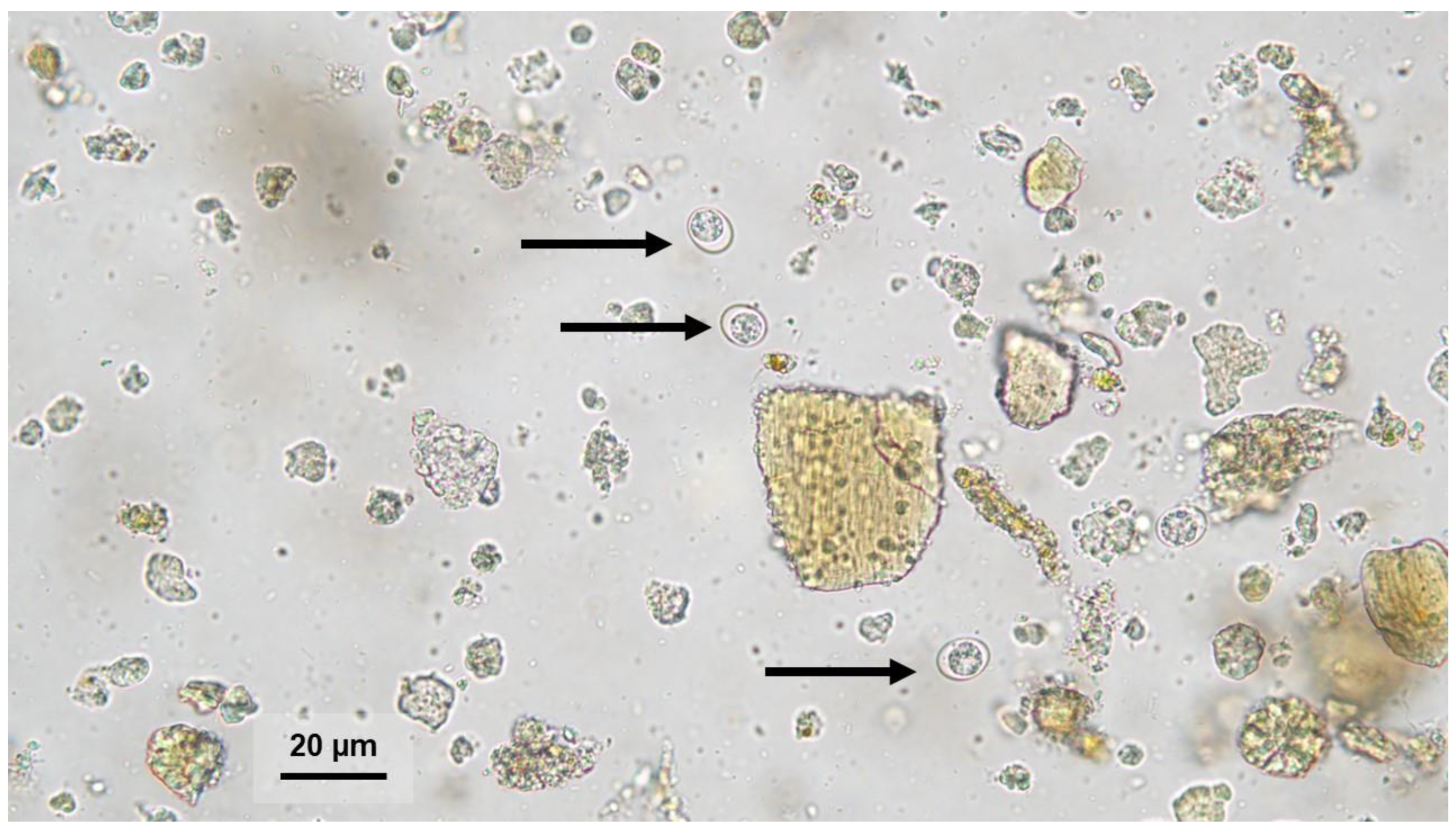
2.1. Sample Collection
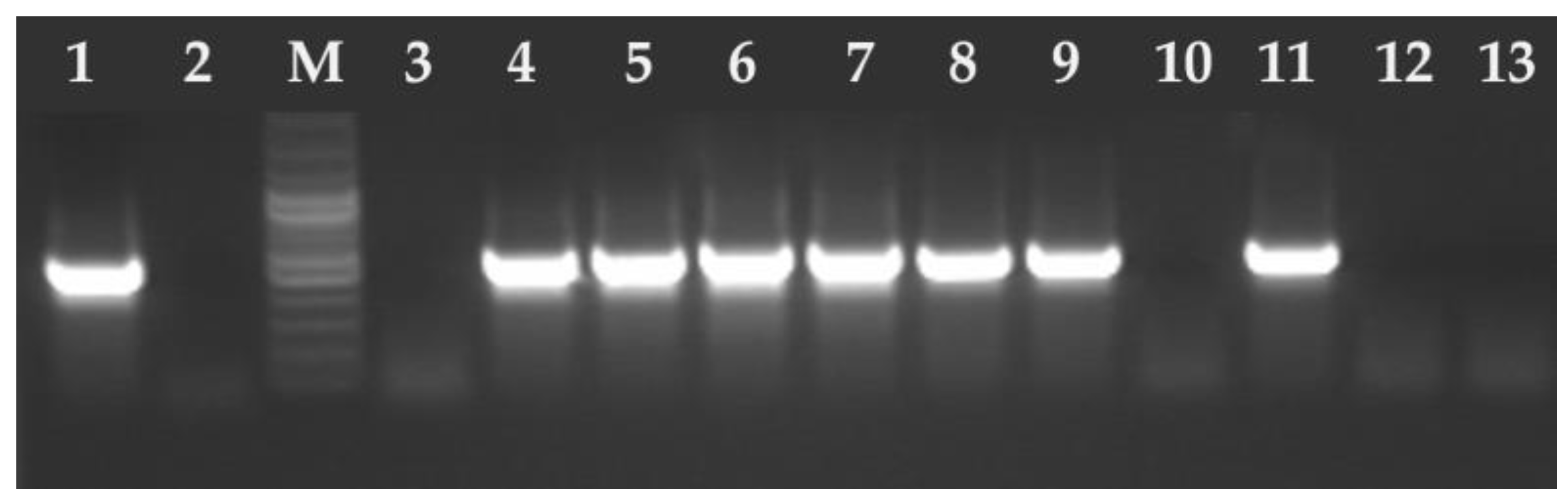
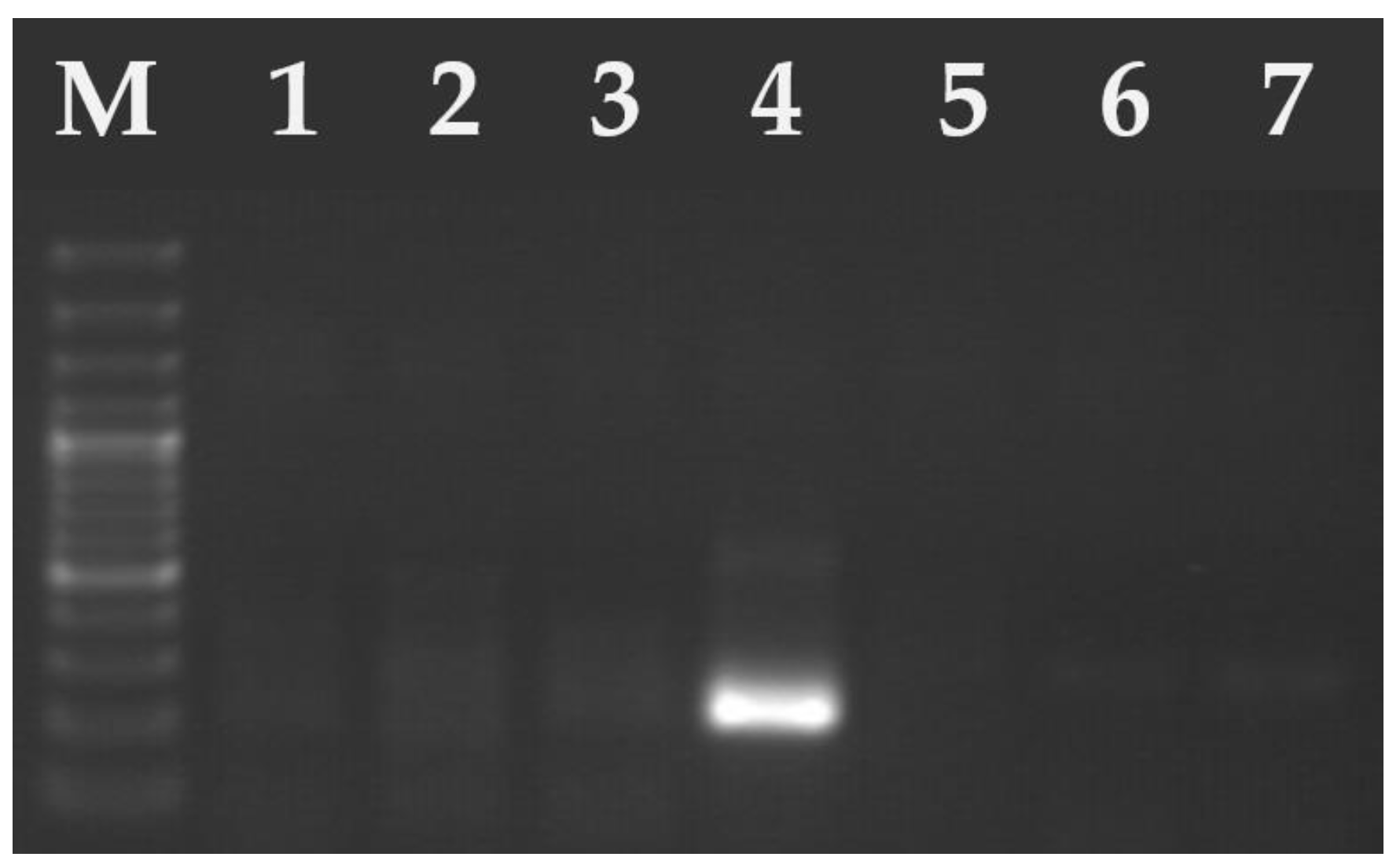
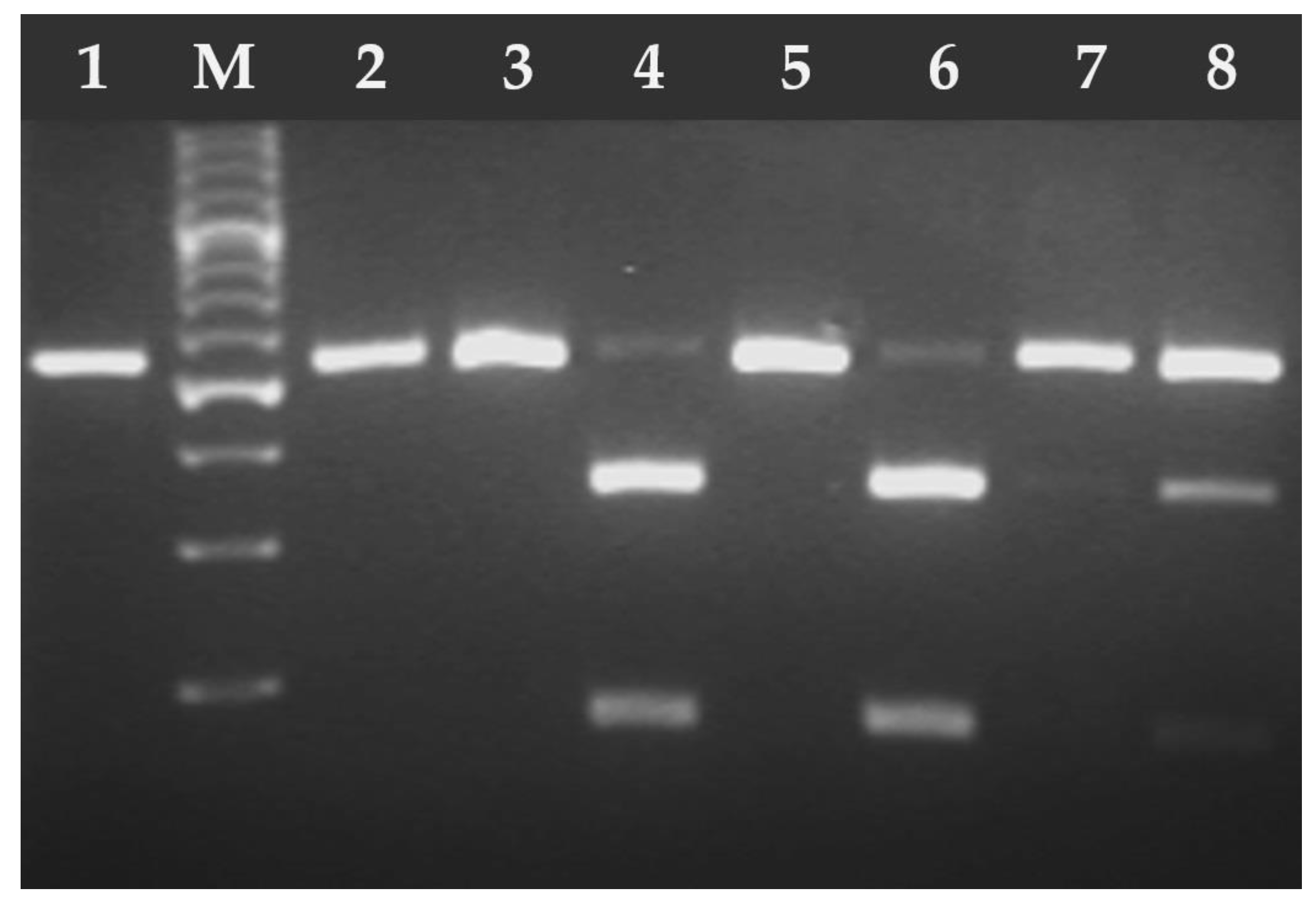
2.2. DNA Isolation and Molecular Analysis
2.3. Statistical Analysis
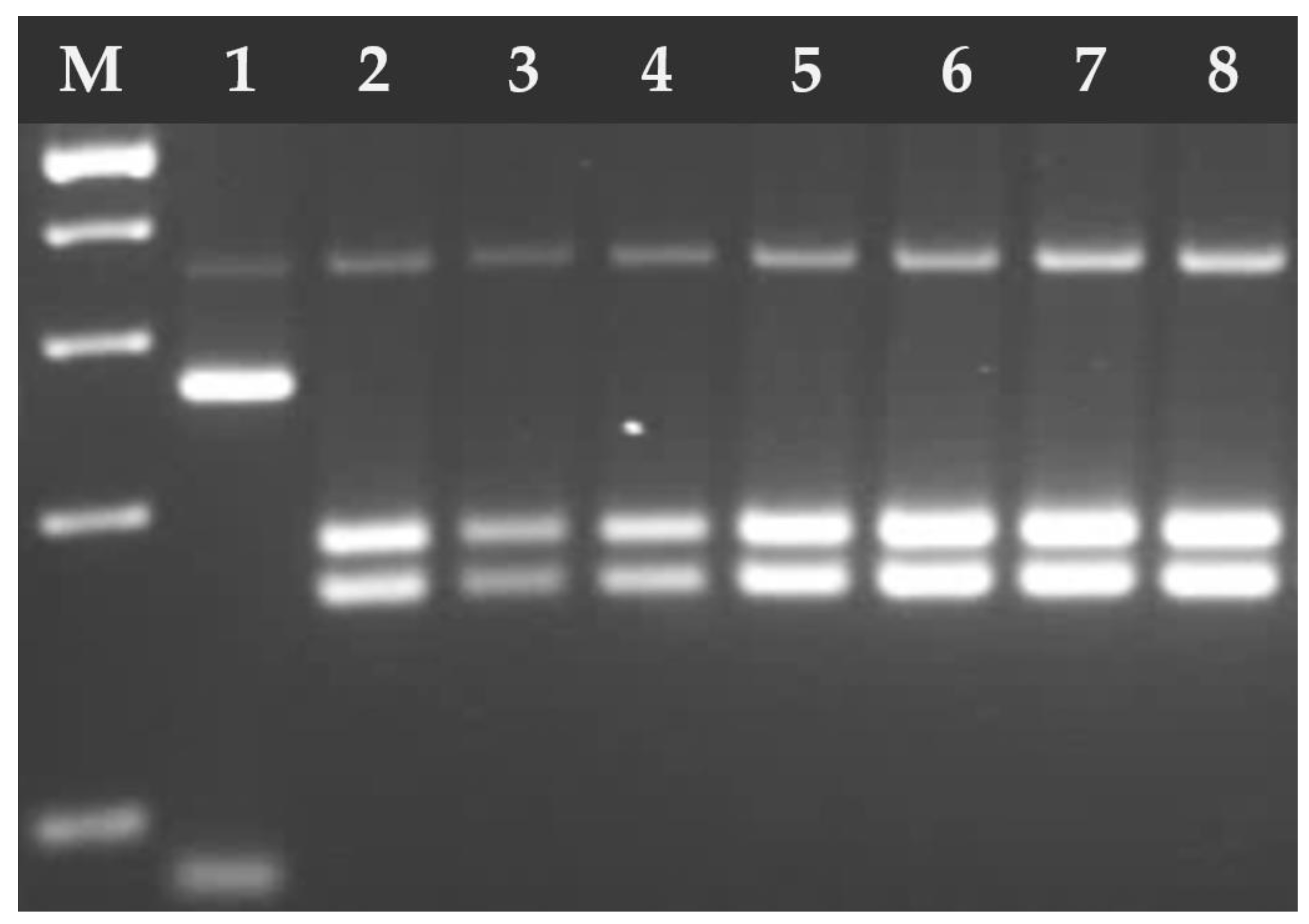
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dubey, J.P. Advances in the life cycle of Toxoplasma gondii. Int. J. Parasitol. 1998, 28, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. History of the discovery of the life cycle of Toxoplasma gondii. Int. J. Parasitol. 2009, 39, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Sreekumar, C. Redescription of Hammondia hammondi and its differentiation from Toxoplasma gondii. Int. J. Parasitol. 2003, 33, 1437–1453. [Google Scholar] [CrossRef]
- Dubey, J.P.; Miller, N.L.; Frenkel, J.K. The Toxoplasma gondii oocyst from cat feces. J. Exp. Med. 1970, 132, 636–662. [Google Scholar] [CrossRef]
- Dubey, J.P.; Frenkel, J.K. Cyst-Induced Toxoplasmosis in Cats. J. Protozool. 1972, 19, 155–177. [Google Scholar] [CrossRef]
- Dubey, J.P.; Steitel, R.H. Further studies on the transmission of Hammondia hammondi in cats. J. Parasitol. 1976, 62, 548–551. [Google Scholar] [CrossRef]
- Dubey, J.P.; Ferreira, L.R.; Martins, J.; Jones, J.L. Sporulation and survival of Toxoplasma gondii oocysts in different types of commercial cat litter. J. Parasitol. 2011, 97, 751–754. [Google Scholar] [CrossRef]
- Holland, G.N. Ocular Toxoplasmosis: A Global Reassessment. Part I: Epidemiology and Course of Disease. Am. J. Ophthalmol. 2003, 136, 973–988. [Google Scholar] [CrossRef]
- Kijlstra, A.; Jongert, E. Control of the Risk of Human Toxoplasmosis Transmitted by Meat. Int. J. Parasitol. 2008, 38, 1359–1370. [Google Scholar] [CrossRef]
- Robert-Gangneux, F.; Darde, M.L. Epidemiology of and Diagnostic Strategies for Toxoplasmosis. Clin. Microbiol. Rev. 2012, 25, 264–296. [Google Scholar] [CrossRef]
- Cook, A.J.C.; Gilbert, R.E.; Buffolano, W.; Petersen, E.; Jenum, P.A.; Foulon, W.; Semprini, A.E.; Dunn, D.T. Sources of Toxoplasma Infection in Pregnant Women: European Multicentre Case-Control Study. BMJ 2000, 321, 142–147. [Google Scholar] [CrossRef] [PubMed]
- El Deeb, H.K.; Salah-Eldin, H.; Khodeer, S.; Allah, A.A. Prevalence of Toxoplasma gondii Infection in Antenatal Population in Menoufia Governorate. Egypt. Acta Trop. 2012, 124, 185–191. [Google Scholar] [CrossRef]
- Tong, W.H.; Pavey, C.; O’Handley, R.; Vyas, A. Behavioral Biology of Toxoplasma gondii Infection. Parasit. Vectors 2021, 14, 77. [Google Scholar] [CrossRef]
- Dubey, J.P.; Carpenter, J.L. Histologically Confirmed Clinical Toxoplasmosis in Cats: 100 Cases (1952–1990). J. Am. Vet. Med. Assoc. 1993, 203, 1556–1566. [Google Scholar] [CrossRef]
- Dabritz, H.A.; Conrad, P.A. Cats and Toxoplasma: Implications for Public Health. Zoonoses Public Health 2010, 57, 34–52. [Google Scholar] [CrossRef] [PubMed]
- Zulpo, D.L.; Sammi, A.S.; Dos Santos, J.R.; Sasse, J.P.; Martins, T.A.; Minutti, A.F.; Cardim, S.T.; de Barros, L.D.; Navarro, I.T.; Garcia, J.L. Toxoplasma gondii: A study of oocyst re-shedding in domestic cats. Vet. Parasitol. 2018, 249, 17–20. [Google Scholar] [CrossRef]
- Dubey, J.P. Effect of immunization of cats with Isospora felis and BCG on immunity to reexcretion of Toxoplasma gondii oocysts. J. Protozool. 1978, 25, 380–382. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Toxoplasma, Hammondia, Sarcocystis, and Other Tissue Cyst-Forming Coccidia of Humans and Animals. In Parasitic Protozoa; Kreier, J.P., Ed.; Academic Press: New York, NY, USA, 1993; Volume 6, pp. 1–158. [Google Scholar]
- Schares, G.; Herrmann, D.C.; Beckert, A.; Schares, S.; Hosseininejad, M.; Pantchev, N.; Globokar Vrhovec, M.; Conraths, F.J. Characterization of a repetitive DNA fragment in Hammondia hammondi and its utility for the specific differentiation of H. hammondi from Toxoplasma gondii by PCR. Mol. Cell Probes 2008, 22, 244–251. [Google Scholar] [CrossRef]
- Monteiro, R.M.; Pena, H.F.; Gennari, S.M.; de Souza, S.O.; Richtzenhain, L.J.; Soares, R.M. Differential Diagnosis of Oocysts of Hammondia-like Organisms of Dogs and Cats by PCR-RFLP Analysis of 70-Kilodalton Heat Shock Protein (HSP70). Gene. Parasitol. Res. 2008, 103, 235–238. [Google Scholar] [CrossRef]
- Grigg, M.E.; Boothroyd, J.C. Rapid identification of virulent type I strains of the protozoan pathogen Toxoplasma gondii by PCR-Restriction Fragment Length Polymorphism analysis at the B1 gene. J. Clin. Microbiol. 2001, 39, 398–400. [Google Scholar] [CrossRef]
- Ajzenberg, D.; Banuls, A.L.; Su, C.; Dumetre, A.; Demar, M.; Carme, B.; Darde, M.L. Genetic Diversity, Clonality and Sexuality in Toxoplasma gondii. Int. J. Parasitol. 2004, 34, 1185–1196. [Google Scholar] [CrossRef]
- Howe, D.K.; Sibley, L.D. Toxoplasma gondii Comprises Three Clonal Lineages: Correlation of Parasite Genotype with Human Disease. J. Infect. Dis. 1995, 172, 1561–1566. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Zhang, X.; Dubey, J.P. Genotyping of Toxoplasma gondii by Multilocus PCR-RFLP Markers: A High-Resolution and Simple Method for Identification of Parasites. Int. J. Parasitol. 2006, 36, 841–848. [Google Scholar] [CrossRef]
- Sroka, J.; Kusyk, P.; Bilska-Zając, E.; Karamon, J.; Dutkiewicz, J.; Wójcik-Fatla, A.; Zajac, V.; Stojecki, K.; Rozycki, M.; Cencek, T. Seroprevalence of Toxoplasma gondii Infection in Goats from the South-West Region of Poland and the Detection of T. gondii DNA in Goat Milk. Folia Parasitol. 2017, 64, 23. [Google Scholar] [CrossRef] [PubMed]
- Homan, W.L.; Vercammen, M.; Debraekeleer, J.; Verschueren, H. Identification of a 200-to 300-fold repetitive 529 bp DNA fragment in Toxoplasma gondii, and its use for diagnostic and quantitative PCR. Int. J. Parasitol. 2000, 30, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Grigg, M.E.; Ganatra, J.; Boothroyd, J.C.; Margolis, T.P. Unusual abundance of atypical strains associated with human ocular toxoplasmosis. J. Infect. Dis. 2001, 184, 633–639. [Google Scholar] [CrossRef]
- Howe, D.K.; Honore, S.; Derouin, F.; Sibley, L.D. Determination of genotypes of Toxoplasma gondii strains isolated from patient with toxoplasmosis. J. Clin. Microbiol. 1997, 35, 1411–1414. [Google Scholar] [CrossRef]
- Khan, A.; Su, C.; German, M.; Storch, G.A.; Clifford, D.B.; Sibley, L.D. Genotyping of Toxoplasma gondii strains from immunocompromised patients reveals high prevalence of type I strains. J. Clin. Microbiol. 2005, 43, 5881–5887. [Google Scholar] [CrossRef]
- Fazaeli, A.; Carter, P.E.; Dardé, M.L.; Pennington, T.H. Molecular typing of Toxoplasma gondii strains by GRA6 gene sequence analysis. Int. J. Parasitol. 2000, 30, 637–642. [Google Scholar] [CrossRef]
- Su, C.; Shwab, E.K.; Zhou, P.; Zhu, X.Q.; Dubey, J.P. Moving towards an integrated approach to molecular detection and identification of Toxoplasma gondii. Parasitology 2010, 137, 1–11. [Google Scholar] [CrossRef]
- Brennan, A.; Donahoe, S.L.; Beatty, J.A.; Belov, K.; Lindsay, S.; Briscoe, K.A.; Šlapeta, J.; Barrs, V.R. Comparison of genotypes of Toxoplasma gondii in domestic cats from Australia with latent infection or clinical toxoplasmosis. Vet. Parasitol. 2016, 228, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Fenton, H.; Hall, J.; Rose, K.; Peddemors, V.M.; Šlapeta, J. Dolphins share Toxoplasma gondii Type II genotypes with terrestrial animals: Evidence of terrestrial T. gondii contamination in marine environments. Vet. Parasitol. 2025, 335, 110439. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Ferguson, D.J. Life Cycle of Hammondia hammondi (Apicomplexa: Sarcocystidae) in Cats. J. Eukaryot. Microbiol. 2015, 62, 346–352. [Google Scholar] [CrossRef]
- Dubey, J.P.; Hoover, E.A.; Walls, K.W. Effect of age and sex on the acquisition of immunity to toxoplasmosis in cats. J. Protozool. 1977, 24, 184–186. [Google Scholar] [CrossRef]
- Lappin, M.R.; Greene, C.E.; Winston, S.; Toll, S.L.; Epstein, M.E. Clinical feline toxoplasmosis: Serological diagnosis and therapeutic management of 15 cases. J. Vet. Intern. Med. 1989, 3, 139–143. [Google Scholar] [CrossRef]
- Frenkel, J.K.; Dubey, J.P. Hammondia hammondi gen. nov., sp.nov., from domestic cats, a new coccidian related to Toxoplasma and Sarcocystis. Z. Parasitenkd. 1975, 46, 3–12. [Google Scholar] [CrossRef]
- Berger-Schoch, A.E.; Herrmann, D.C.; Schares, G.; Müller, N.; Bernet, D.; Gottstein, B.; Frey, C.F. Prevalence and genotypes of Toxoplasma gondii in feline faeces (oocysts) and meat from sheep, cattle and pigs in Switzerland. Vet. Parasitol. 2011, 177, 290–297. [Google Scholar] [CrossRef]
- Dubey, J.P. Duration of immunity to shedding of Toxoplasma gondii oocysts by cats. J. Parasitol. 1995, 81, 410–415. [Google Scholar] [CrossRef]
- Elmore, S.A.; Jones, J.L.; Conrad, P.A.; Patton, S.; Lindsay, D.S.; Dubey, J.P. Toxoplasma gondii: Epidemiology, feline clinical aspects, and prevention. Trends Parasitol. 2010, 26, 190–196. [Google Scholar] [CrossRef]
- Lappin, M.R.; VanLare, K.A.; Seewald, W.; Roycroft, L.M.; Scorza, A.V.; King, S.; Roberts, E.S. Effect of oral administration of cyclosporine on Toxoplasma gondii infection status of cats. Am. J. Vet. Res. 2015, 76, 351–357. [Google Scholar] [CrossRef]
- Last, R.D.; Suzuki, Y.; Manning, T.; Lindsay, D.; Galipeau, L.; Whitbread, T.J. A case of fatal systemic toxoplasmosis in a cat being treated with cyclosporin A for feline atopy. Vet. Dermatol. 2004, 15, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Cohen, T.M.; Blois, S.; Vince, A.R. Fatal extraintestinal toxoplasmosis in a young male cat with enlarged mesenteric lymph nodes. Can. Vet. J. 2016, 57, 483–486. [Google Scholar]
- Dubey, J.P. Toxoplasmosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 7–56. [Google Scholar]
- Scorza, A.V.; Tyrrell, P.; Wennogle, S.; Chandrashekar, R.; Lappin, M.R. Experimental infection of cats with Cystoisospora felis. J. Vet. Intern. Med. 2021, 35, 269–272. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Prowell, M. Ante-mortem diagnosis, diarrhea, oocyst shedding, treatment, isolation, and genetic typing of Toxoplasma gondii associated with clinical toxoplasmosis in a naturally infected cat. J. Parasitol. 2013, 99, 158–160. [Google Scholar] [CrossRef]
- Deksne, G.; Petrusēviča, A.; Kirjušina, M. Seroprevalence and factors associated with Toxoplasma gondii infection in domestic cats from urban areas in Latvia. J. Parasitol. 2013, 99, 48–50. [Google Scholar] [CrossRef]
- Raue, K.; Heuer, L.; Böhm, C.; Wolken, S.; Epe, C.; Strube, C. 10-year parasitological examination results (2003 to 2012) of faecal samples from horses, ruminants, pigs, dogs, cats, rabbits and hedgehogs. Parasitol. Res. 2017, 116, 3315–3330. [Google Scholar] [CrossRef]
- Wąsiatycz, G. Prevalence of Toxoplasma gondii infection of cats in Poznań and its district in the aspect of the danger of parasite invasion of people. Wiad. Parazytol. 1998, 44, 693–704. [Google Scholar]
- Schares, G.; Vrhovec, M.G.; Pantchev, N.; Herrmann, D.C.; Conraths, F.J. Occurrence of Toxoplasma gondii and Hammondia hammondi oocysts in the faeces of cats from Germany and other European countries. Vet. Parasitol. 2008, 152, 34–45. [Google Scholar] [CrossRef]
- Pena, H.F.; Soares, R.M.; Amaku, M.; Dubey, J.P.; Gennari, S.M. Toxoplasma gondii infection in cats from São Paulo state, Brazil: Seroprevalence, oocyst shedding, isolation in mice, and biologic and molecular characterization. Res. Vet. Sci. 2006, 81, 58–67. [Google Scholar] [CrossRef]
- Frenkel, J.; Ruiz, A.; Chinchilla, M. Soil survival of Toxoplasma oocysts in Kansas and Costa Rica. Am. J. Trop. Med. Hyg. 1975, 24, 439–443. [Google Scholar] [CrossRef]
- Grigg, M.E.; Bonnefoy, S.; Hehl, A.B.; Suzuki, Y.; Boothroyd, J.C. Success and virulence in Toxoplasma as the result of sexual recombination between two distinct ancestries. Science 2001, 294, 161–165. [Google Scholar] [CrossRef]
- Długońska, H. Toxoplasma gondii—known and unknown parasite. Ann. Parasitol. 2008, 54, 199–204. [Google Scholar]
- Sibley, L.D.; Boothroyd, J.C. Virulent strains of Toxoplasma gondii comprise a single clonal lineage. Nature 1992, 359, 82–85. [Google Scholar] [CrossRef]
- Khan, A.; Fux, B.; Su, C.; Dubey, J.P.; Dardé, M.L.; Ajioka, J.W.; Rosenthal, B.M.; Sibley, L.D. Recent transcontinental sweep of Toxoplasma gondii driven by a single monomorphic chromosome. Proc. Natl. Acad. Sci. USA 2007, 104, 14872–14877. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wang, H.; Wang, H.; Qin, H.; Xiao, J. Land use and soil contamination with Toxoplasma gondii oocysts in urban areas. Sci. Total Environ. 2016, 568, 1086–1091. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, T.; Marcet, P.L.; Graham, D.H.; Dahl, E.R.; Dubey, J.P. Globalization and the population structure of Toxoplasma gondii. Proc. Natl. Acad. Sci. USA 2006, 103, 11423–11428. [Google Scholar] [CrossRef] [PubMed]
- Chemoh, W.; Sawangjaroen, N.; Nissapatorn, V.; Sermwittayawong, N. Genotyping of Toxoplasma gondii isolated from cat feces in Songkhla, Southern Thailand. Vet. Parasitol. Reg. Stud. Rep. 2018, 13, 105–109. [Google Scholar] [CrossRef]
- Khodaverdi, M.; Razmi, G. Prevalence and genotyping of Toxoplasma gondii in stray cats in Mashhad area, Iran. BMC Vet. Res. 2019, 15, 463. [Google Scholar] [CrossRef]
- Fernández-Escobar, M.; Schares, G.; Maksimov, P.; Joeres, M.; Ortega-Mora, L.M.; Calero-Bernal, R. Toxoplasma gondii genotyping: A closer look into Europe. Front. Cell. Infect. Microbiol. 2022, 12, 842595. [Google Scholar] [CrossRef]
- Spycher, A.; Geigy, C.; Howard, J.; Posthaus, H.; Gendron, K.; Gottstein, B.; Debache, K.; Herrmann, D.C.; Schares, G.; Frey, C.F. Isolation and genotyping of Toxoplasma gondii causing fatal systemic toxoplasmosis in an immunocompetent 10-year-old cat. J. Vet. Diagn. Invest. 2011, 23, 104–108. [Google Scholar] [CrossRef]
- Mancianti, F.; Nardoni, S.; Mugnaini, L.; Zambernardi, L.; Guerrini, A.; Gazzola, V.; Papini, R.A. A retrospective molecular study of select intestinal protozoa in healthy pet cats from Italy. J. Feline Med. Surg. 2015, 17, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Montoya, A.; Miró, G.; Mateo, M.; Ramírez, C.; Fuentes, I. Molecular characterization of Toxoplasma gondii isolates from cats in Spain. J. Parasitol. 2008, 94, 1044–1046. [Google Scholar] [CrossRef]
- Herrmann, D.C.; Pantchev, N.; Vrhovec, M.G.; Barutzki, D.; Wilking, H.; Fröhlich, A.; Lüder, C.G.; Conraths, F.J.; Schares, G. Atypical Toxoplasma gondii genotypes identified in oocysts shed by cats in Germany. Int. J. Parasitol. 2010, 40, 285–292. [Google Scholar] [CrossRef]
- Sroka, J.; Wójcik-Fatla, A.; Bilska-Zając, E.; Karamon, J.; Zdybel, J.M.; Piotrowska, W.; Dąbrowska, J.; Samorek-Pieróg, M.; Korpysa-Dzirba, W.; Cencek, T. Molecular confirmation of systemic toxoplasmosis in a cat. Ann. Agric. Environ. Med. 2023, 30, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Pardo Gil, M.; Hegglin, D.; Briner, T.; Ruetten, M.; Müller, N.; Moré, G.; Frey, C.F.; Deplazes, P.; Basso, W. High prevalence rates of Toxoplasma gondii in cat-hunted small mammals—Evidence for parasite-induced behavioural manipulation in the natural environment? Int. J. Parasitol. Parasites Wildl. 2023, 20, 108–116. [Google Scholar] [CrossRef]
- Sroka, J. Occurrence of Toxoplasma gondii infection among free-living small rodents and insectivores in the Lublin Province—the role of these animals in the epidemiology of toxoplasmosis. Med. Og. Nauk. Zdr. 2021, 27, 448–452. [Google Scholar] [CrossRef]
- Grzybek, M.; Antolová, D.; Tołkacz, K.; Alsarraf, M.; Behnke-Borowczyk, J.; Nowicka, J.; Paleolog, J.; Biernat, B.; Behnke, J.M.; Bajer, A. Seroprevalence of Toxoplasma gondii among sylvatic rodents in Poland. Animals 2021, 11, 1048. [Google Scholar] [CrossRef]
- Nowicka, J.; Antolova, D.; Lass, A.; Biernat, B.; Baranowicz, K.; Goll, A.; Krupinska, M.; Ferra, B.; Strachecka, A.; Behnke, J.M.; et al. Identification of Toxoplasma gondii in wild rodents in Poland by molecular and serological techniques. Ann. Agric. Environ. Med. 2024, 31, 626–630. [Google Scholar] [CrossRef]
- Must, K.; Hytonen, M.K.; Orro, T.; Lohi, H.; Jokelainen, P. Toxoplasma gondii seroprevalence varies by cat breed. PLoS ONE 2017, 12, e0184659. [Google Scholar] [CrossRef]
- Gyorke, A.; Opsteegh, M.; Mircean, V.; Iovu, A.; Cozma, V. Toxoplasma gondii in Romanian household cats: Evaluation of serological test, epidemiology and risk factors. Prev. Vet. Med. 2011, 102, 321–328. [Google Scholar] [CrossRef]
- Brennan, A.; Hawley, J.; Dhand, N.; Boland, L.; Beatty, J.A.; Lappin, M.R.; Barrs, V.R. Seroprevalence and risk factors for Toxoplasma gondii infection in owned domestic cats in Australia. Vector Borne Zoonotic Dis. 2020, 20, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Chessa, G.; Chisu, V.; Porcu, R.; Masala, G. Molecular characterization of Toxoplasma gondii Type II in sheep abortion in Sardinia, Italy. Parasite 2014, 21, 6. [Google Scholar] [CrossRef] [PubMed]
- Aspinall, T.V.; Marlee, D.; Hyde, J.E.; Sims, P.F. Prevalence of Toxoplasma gondii in commercial meat products as monitored by polymerase chain reaction—Food for thought? Int. J. Parasitol. 2002, 32, 1193–1199. [Google Scholar] [CrossRef]
- Sroka, J.; Bilska-Zając, E.; Wójcik-Fatla, A.; Zając, V.; Dutkiewicz, J.; Karamon, J.; Piotrowska, W.; Cencek, T. Detection and Molecular Characteristics of Toxoplasma gondii DNA in Retail Raw Meat Products in Poland. Foodborne Pathog. Dis. 2019, 16, 195–204. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parasite Species | Forward Primer | Sequence (5′-3′) | Reverse Primer | Sequence (5′-3′) |
|---|---|---|---|---|
| Hammondia hammondi | Hham34F | ATCCCATTCCGGCTTCAGTCTTTC | Hham3R | ACAGCGGAGCCGAAGTTGGTTT |
| Toxoplasma gondii | TOX4 | CGCTGCAGGGAGGAAGACGAAAGTTG | TOX5 | CGCTGCAGACACAGTGCATCTGGATT |
| Markers | PCR External Primers (Sequence 5′-3′) | Nested PCR Internal Primers (Sequence 5′-3′) | Nested PCR Product | Restriction Enzymes |
|---|---|---|---|---|
| B1 | F:TGTTCTGTCCTATCGCAACG | F:TCTTCCCAGACGTGGATTTC | 530 bp | Pml1 |
| R:ACGGATGCAGTTCCTTTCTG | R:CTCGACAATACGCTGCTTGA | |||
| SAG1 | F:GTTCTAACCACGCACCCTGAG | F:CAATGTGCACCTGTAGGAAGC | 390 bp | Sau96I and HaeII (double digest) |
| R:AAGAGTGGGAGGCTCTGTGA | R:GTGGTTCTCCGTCGGTGTGAG | |||
| 5′-SAG2 | F:GCTACCTCGAACAGGAACAC | F:GCATCAACAGTCTTCGTTGC | 242 bp | Sau3AI |
| R:GAAATGTTTCAGGTTGCTGC | R:GCAAGAGCGAACTTGAACAC | |||
| 3′-SAG2 | F:TCTGTTCTCCGAAGTGACTCC | F:ATTCTCATGCCTCCGCTTC | 222 bp | HhaI |
| R:TCAAAGCGTGCATTATCGC | R:AACGTTTCACGAAGGCACAC | |||
| alt. SAG2 | F:GGAACGCGAACAATGAGTTT | F:ACCCATCTGCGAAGAAAACG | 546 bp | HinfI and TaqI (separate digest) |
| R:GCACTGTTGTCCAGGGTTTT | R:ATTTCGACCAGCGGGAGCAC | |||
| SAG3 | F:CAACTCTCACCATTCCACCC | F:TCTTGTCGGGTGTTCACTCA | 225 bp | NciI |
| R:GCGCGTTGTTAGACAAGACA | R:CACAAGGAGACCGAGAAGGA | |||
| BTUB | F:TCCAAAATGAGAGAAATCGT | F:GAGGTCATCTCGGACGAACA | 411 bp | BsiE and TaqI (double digest) |
| R:AAATTGAAATGACGGAAGAA | R:TTGTAGGAACACCCGGACGC | |||
| GRA6 | F:ATTTGTGTTTCCGAGCAGGT | F:TTTCCGAGCAGGTGACCT | 344 bp | MseI |
| R:GCACCTTCGCTTGTGGTT | R:TCGCCGAAGAGTTGACATAG | |||
| C22-8 | F:TGATGCATCCATGCGTTTAT | F:TCTCTCTACGTGGACGCC | 521 bp | BsmAI and MboII (double digest) |
| R:CCTCCACTTCTTCGGTCTCA | R:AGGTGCTTGGATATTCGC | |||
| C29-2 | F:ACCCACTGAGCGAAAAGAAA | F:AGTTCTGCAGAGTGTCGC | 446 bp | HpyCH4IV and RsaI (double digest) |
| R:AGGGTCTCTTGCGCATACAT | R:TGTCTAGGAAAGAGGCGC | |||
| L358 | F:TCTCTCGACTTCGCCTCTTC | F:AGGAGGCGTAGCGCAAGT | 419 bp | HaeIII and NlaIII (double digest) |
| R:GCAATTTCCTCGAAGACAGG | R:CCCTCTGGCTGCAGTGCT | |||
| PK1 | F:GAAAGCTGTCCACCCTGAAA | F:CGCAAAGGGAGACAATCAGT | 903 bp | AvaI and RsaI (double digest) |
| R:AGAAAGCTCCGTGCAGTGAT | R:TCATCGCTGAATCTCATTGC | |||
| Apico | F:TGGTTTTAACCCTAGATTGTGG | F:GCAAATTCTTGAATTCTCAGTT | 640 bp | AflII and DdeI (double digest) |
| F:AAACGGAATTAATGAGATTTGAA | R:GGGATTCGAACCCTTGATA |
| Toxoplasma gondii | B1 | SAG1 | SAG2 5′+3′ | SAG2 alt. | SAG3 | BTUB | GRA6 | C22-8 | C29-2 | L358 | PK1 | Apico | ToxoDB | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Type I (GT1) | I | I | I | I | I | I | I | I | I | I | I | I | #10 | |||
| Type II (ME49) | II/ III | II/ III | II | II | II | II | II | II | II | II | II | II | #1 | |||
| Type III (CTG) | II/ III | II/ III | III | III | III | III | III | III | III | III | III | III | #2 | |||
| Cats 5, 7, 8 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 | |||
| Genotypes of Toxoplasma gondii detected in cats feces in Poland 2020–2024 | ||||||||||||||||
| T. gondii DNA isolate | Sex | Age | Year | B1 | SAG1 | SAG2 5′+3′ | SAG2 alt. | SAG3 | BTUB | GRA6 | C22-8 | C29-2 | L358 | PK1 | Apico | ToxoDB |
| Cat 1 | F | 5M | 2020 | I | I | I | I | I | I | I | I | I | I | I | I | #10 |
| Cat 2 | M | 7M | 2020 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | II | #1 |
| Cat 3 | F | 4Y | 2020 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 4 | F | 5M | 2020 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 5 | M | 2Y | 2020 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 6 | F | 9M | 2020 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 7 | M | 3M | 2020 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 8 | F | 7M | 2020 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | II | #1 |
| Cat 9 | F | 1Y | 2020 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 10 | M | 18Y | 2020 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 11 | F | 8M | 2020 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 12 | F | 9M | 2020 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 13 | M | 4M | 2021 | I | I | I | I | I | I | I | I | I | I | I | I | #10 |
| Cat 14 | M | 4M | 2021 | I | I | I | I | I | I | I | I | I | I | I | I | #10 |
| Cat 15 | M | 3M | 2021 | I | I | I | I | I | I | I | I | I | I | I | I | #10 |
| Cat 16 | F | 2M | 2021 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 17 | M | 11Y | 2021 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 18 | F | 3Y | 2021 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 19 | M | 10Y | 2021 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 20 | F | 8M | 2022 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | II | #1 |
| Cat 21 | M | 13Y | 2022 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 22 | F | 1Y | 2022 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | II | #1 |
| Cat 23 | F | 6M | 2022 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 24 | M | 8Y | 2022 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 25 | M | 9Y | 2022 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 26 | F | 1Y | 2022 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 27 | M | 1Y | 2022 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 28 | M | 2Y | 2022 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | II | #1 |
| Cat 29 | M | 4Y | 2023 | I | I | I | I | I | I | I | I | I | I | I | I | #10 |
| Cat 30 | F | 7M | 2023 | I | I | I | I | I | I | I | I | I | I | I | I | #10 |
| Cat 31 | F | 1Y | 2023 | I | I | I | I | I | I | I | I | I | I | I | I | #10 |
| Cat 32 | F | 6M | 2023 | I | I | I | I | I | I | I | I | I | I | I | I | #10 |
| Cat 33 | M | 13Y | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 34 | M | 2Y | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 35 | F | 4Y | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | II | #1 |
| Cat 36 | M | 8M | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 37 | F | 3Y | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 38 | F | 6Y | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 39 | F | 4M | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 40 | M | 2M | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 41 | M | 3M | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 42 | M | 16Y | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 43 | M | 11Y | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 44 | F | 2M | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 45 | F | 5M | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 46 | M | 9M | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 47 | F | 5M | 2023 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | II | #1 |
| Cat 48 | F | 6M | 2024 | I | I | I | I | I | I | I | I | I | I | I | I | #10 |
| Cat 49 | F | 3Y | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 50 | M | 8Y | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 51 | M | 2Y | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 52 | F | 3M | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 53 | F | 9M | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 54 | F | 2Y | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 55 | M | 8M | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 56 | M | 2Y | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 57 | M | 4M | 2024 | I | I | I | I | I | I | I | I | I | I | I | I | #10 |
| Cat 58 | F | 7M | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | II | #1 |
| Cat 59 | F | 3M | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 60 | F | 1Y | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 61 | F | 2Y | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 62 | M | 11M | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 63 | F | 4M | 2024 | I | I | I | I | I | I | I | I | I | I | I | I | #10 |
| Cat 64 | F | 6M | 2024 | I | I | I | I | I | I | I | I | I | I | I | I | #10 |
| Cat 65 | F | 6M | 2024 | I | I | I | I | I | I | I | I | I | I | I | I | #10 |
| Cat 66 | M | 7M | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Cat 67 | M | 7M | 2024 | II/ III | II/ III | II | II | II | II | II | II | II | II | II | I | #3 |
| Examined Feline Stool Samples | Toxoplasma gondii Positive Samples | Hammondia hammondi Positive Samples | |||
|---|---|---|---|---|---|
| Year | n | n | % | n | % |
| 2020 | 1780 | 12 | 0.67 | 1 | 0.06 |
| 2021 | 5760 | 7 | 0.12 | 6 | 0.10 |
| 2022 | 11,641 | 9 | 0.08 | 5 | 0.04 |
| 2023 | 19,121 | 19 | 0.10 | 4 | 0.02 |
| 2024 | 23,346 | 20 | 0.09 | 2 | 0.01 |
| Total: | 61,648 | 67 | 0.11 | 18 | 0.03 |
| T. gondii | H. hammondi | |||
|---|---|---|---|---|
| Age <1 Y | Age ≥1 | Age <1 | Age ≥1 | |
| Total | 37 | 30 | 8 | 10 |
| Male | 14 | 17 | 5 | 4 |
| Female | 23 | 13 | 3 | 6 |
| Diarrhea | 36 | 18 | 5 | 1 |
| Formed stool | 1 | 12 | 3 | 9 |
| co-infection | 9 | 5 | 1 | 2 |
| Giardia duodenalis | 4 | 1 | 0 | 1 |
| Cystoisospora felis | 2 | 3 | 0 | 0 |
| Tritrichomonas foetus | 1 | 1 | 1 | 1 |
| Toxocara cati | 2 | 0 | 0 | 0 |
| no co-infection | 28 | 25 | 7 | 8 |
| T. gondii clonal lineage I | 11 | 2 | - | - |
| T. gondii clonal lineage II | 26 | 28 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jańczak, D.; Stryjek, R.; Maj, A.K.; Olszewski, J.; Szaluś-Jordanow, O. PCR Detection, Genotyping, and Differentiation of Toxoplasma gondii from Hammondia hammondi Excreted in the Feces of Cats in Poland Between 2020 and 2024. Pathogens 2025, 14, 444. https://doi.org/10.3390/pathogens14050444
Jańczak D, Stryjek R, Maj AK, Olszewski J, Szaluś-Jordanow O. PCR Detection, Genotyping, and Differentiation of Toxoplasma gondii from Hammondia hammondi Excreted in the Feces of Cats in Poland Between 2020 and 2024. Pathogens. 2025; 14(5):444. https://doi.org/10.3390/pathogens14050444
Chicago/Turabian StyleJańczak, Dawid, Rafał Stryjek, Aleksandra Kornelia Maj, Jakub Olszewski, and Olga Szaluś-Jordanow. 2025. "PCR Detection, Genotyping, and Differentiation of Toxoplasma gondii from Hammondia hammondi Excreted in the Feces of Cats in Poland Between 2020 and 2024" Pathogens 14, no. 5: 444. https://doi.org/10.3390/pathogens14050444
APA StyleJańczak, D., Stryjek, R., Maj, A. K., Olszewski, J., & Szaluś-Jordanow, O. (2025). PCR Detection, Genotyping, and Differentiation of Toxoplasma gondii from Hammondia hammondi Excreted in the Feces of Cats in Poland Between 2020 and 2024. Pathogens, 14(5), 444. https://doi.org/10.3390/pathogens14050444