1. Introduction
The order Chiroptera comprises approximately 1487 species distributed worldwide [
1]. Bats are the second most speciose group of mammals, surpassed only by rodents [
2]. Among bats, 47 species are found in the European Union [
3], and 27 have been recorded in mainland Portugal [
4,
5]. These animals are the only mammalian species capable of flying [
6], making them a very diverse and widely geographically distributed group [
7]. They also provide essential ecosystem services, acting as seed dispersers [
8], pollinators [
9], controllers of insect pests [
10], and nutrient recyclers [
11].
Bats are often infested by numerous ectoparasites, such as bat flies (Diptera: Nycteribiidae and Streblidae), bugs (Hemiptera: Cimicidae and Polyctenidae), fleas (Siphonaptera: Ischnopsyllidae), mites (Mesostigmata: Spinturnicidae and Macronyssidae), and ticks (Acari: Ixodidae and Argasidae) [
12]. Due to most bat species’ colonial habits, which imply close contact between individuals, ectoparasite infestations can be intensified, especially in perennial shelters such as cavities [
13,
14].
These ectoparasites may play important roles in the transmission and maintenance of several pathogens, including some with zoonotic potential [
15,
16,
17]. While some of them may cause mortality in bats [
18], others have bats as their natural reservoirs [
19,
20].
Previous studies conducted in Europe have detected vector-borne agents in ectoparasites collected from bats, including piroplasmids (
Babesia spp. and
Theileria spp. [
21,
22]) and Gram-negative bacteria (
Bartonella spp. [
23,
24,
25],
Ehrlichia spp. [
20,
22],
Anaplasma spp. [
24,
26] and
Rickettsia spp. [
22,
24,
27,
28]).
Bartonella spp. are the most frequently detected bacteria in bat-associated ectoparasites. In Hungary,
Bartonella spp. have been reported in ticks (
Ixodes vespertilionis), mites (
Steatonyssus occidentalis and
Spinturnix myoti), and bat flies (
Nycteribia spp.) [
23,
24].
Bartonella DNA was detected in 28.4% of
S. myoti mites collected from bats in Poland [
24]. Szentiványi et al. [
25] detected
Bartonella spp. in
Nycteribia schmidlii and
N. schmidlii scotti bat flies in Spain.
A putative novel
Ehrlichia species, namely,
Ehrlichia sp. AvBat, was described in
Argas vespertilionis ticks collected from bats in France [
27]. Additionally,
Ehrlichia yunnan was detected in
Ixodes simplex ticks from bats in England [
22].
Anaplasma phagocytophilum was molecularly detected in the blood and guano of bats in Poland [
29] and France [
26], respectively.
Rickettsia helvetica was detected in
A. vespertilionis ticks collected from bats in Hungary [
28] and in
Ixodes ricinus in Poland [
23]. Additionally,
Rickettsia spp. was identified in
A. vespertilionis in France [
20] and England [
22]. More recently, [
24] reported the presence of
Rickettsia sp. in
Spinturnix myoti collected from bats in Poland.
DNA from various Babesia species (Babesia canis, Babesia crassa, Babesia venatorum, and Babesia vesperuginis) and Theileria spp. (Theileria capreoli and Theileria orientalis) has been detected in bat-associated ticks (e.g., A. vespertilionis, Ixodes ariadnae, I. simplex, and I. vespertilionis) in Hungary, Romania [
21], and England [
22]. In South America, putative novel lineages of piroplasmids have been detected in both non-hematophagous bats [
24] and vampire bats [
14], highlighting that the diversity of piroplasmids in bats is greater than previously recognized.
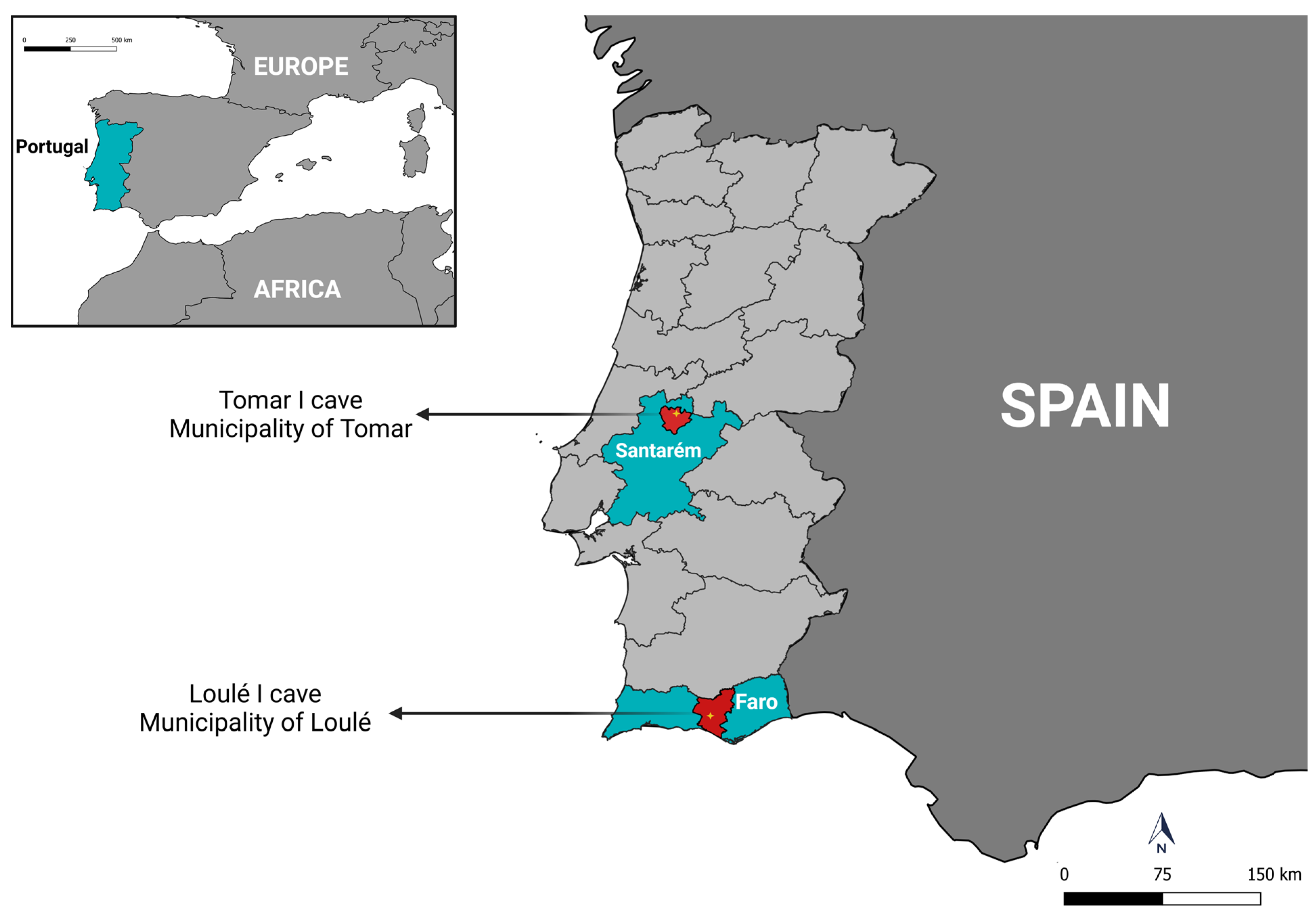
This state of the art shows that studies focused on bat ectoparasites and the vector-borne agents they may carry are restricted to a few countries in Europe, and very little is known about this topic in Portugal. Therefore, the present study aimed to investigate the molecular occurrence of vector-borne bacteria (Anaplasma spp., Bartonella spp., Ehrlichia spp., and Rickettsia spp.) and protozoa (Babesia spp. and Theileria spp.) in ticks, mites, and bat flies parasitizing cave-dwelling bats from mainland Portugal.
3. Results
3.1. Bat Identification and Diversity
A total of 280 bats were collected in cave A. Of these, 270 (96.4%) were identified as belonging to the species Miniopterus schreibersii (Kühl, 1817) (Miniopteridae), of which 172 (63.7%) were female and 98 (36.3%) were male. The remaining ten specimens included four (1.4%) identified as males belonging to the species Myotis myotis (Borkhausen, 1797) (Vespertilionidae); three (1.1%) identified as females belonging to the species Rhinolophus mehelyi Matschie, 1901 (Rhinolophidae); two (0.7%) (one male and one female) identified as Rhinolophus euryale Blasius, 1853; and one (0.4%) identified as a female belonging to the species Myotis escalerai Cabrera, 1904.
In cave B, 110 bats were collected. Of these, 108 (98.2%) were identified as belonging to the species M. schreibersii, including 84 (77.8%) males and 24 (22.2%) females, and 2 (1.8%) females were identified as belonging to the species R. mehelyi.
Table 1 presents the abundance of bats, species richness, Shannon’s index of bat species diversity, evenness index, and Simpson’s diversity index of bats in caves A and B. It shows that cave A presented greater abundance, species richness, and diversity than cave B and that one species (
M. schreibersii) dominates the community in both caves.
3.2. Ectoparasites Sampling and Identification
As a convenience sample, 42 specimens of
M. schreibersii from each cave (29 females and 13 males from cave A, and 11 females and 31 males from cave B) were inspected for the presence of ectoparasites. Four species of ectoparasites were identified on males and females of
M. schreibersii from both caves, totaling 679 specimens. Among these, one species of hard tick,
Ixodes simplex Neumann, 1906 (
Supplementary File), one species of wing mite,
Spinturnix myoti (Kolenati, 1856), and two species of nycteribiid flies,
Penicillidia conspicua Speiser, 1901, and
Nycteribia schmidlii Schiner, 1853, were found.
On these bats, there was a 100% prevalence of the spinturnicid mite
S. myoti. Interestingly, this mite species also exhibited the highest mean intensity and mean abundance indices, yet it had the lowest aggregation rate compared to the other ectoparasite species in the sample (
Table 2). The nycteribiid fly
P. conspicua had the second-highest prevalence, both when considering data from both caves combined and when analyzed separately by cave (
Table 2). Parasitological indices for all sampled ectoparasite species are detailed in
Table 2. When we compared the number of ectoparasites on bats separately by sex, only the tick
I. simplex was significantly more abundant on female hosts in cave A (
Table 3).
Out of the 13
M. schreibersii male bats sampled in cave A, 6 (46.15%) were infested only with
S. myoti, 5 (38.46%) were coinfested with
S. myoti and
P. conspicua, 1 (7.69%) was coinfested with
S. myoti and
N. schmidlii, and 1 (7.69%) was triply infested with
I. simplex,
S. myoti, and
N. schmidlii. Among the 29 females, 7 (24.13%) were infested only with
S. myoti, 6 (20.68%) were triply infested with
S. myoti,
I. simplex, and
P. conspicua, 5 (17.24%) were coinfested with
S. myoti and
I. simplex, 5 (17.24%) were coinfested with
S. myoti and
P. conspicua, and 2 (6.89%) were coinfested with
S. myoti and
N. schmidlii. Two (6.89%) bats were triply infested with
S. myoti,
I. simplex, and
N. schmidlii, and two (6.89%) bats were triply infested with
S. myoti,
P. conspicua, and
N. schmidlii (
Figure 2, cave A).
Out of the 31
M. schreibersii male bats sampled in cave B, 18 (58.06%) were infested only with
S. myoti, 6 (19.35%) were coinfested with
S. myoti and
P. conspicua, 2 (6.45%) were coinfested with
I. simplex and
S. myoti, 2 (6.45%) were coinfested with
S. myoti and
N. schmidlii, 2 (6.45%) were triply infested with
S. myoti,
P. conspicua, and
N. schmidlii, and 1 (3.22%) was infested with
S. myoti,
I. simplex, and
P. conspicua. Of the 11 females, 9 (81.81%) were infested only with
S. myoti, and 2 (18.18%) were coinfested with
S. myoti and
P. conspicua (
Figure 2, cave B). The consensus sequence confirming the taxonomic identification of ticks as
I. simplex was deposited in the GenBank database (accession number: PQ834791).
3.3. Molecular Screening for Vector-Borne Pathogens
All 576 S. myoti pools and all individually processed bat flies (44 P. conspicua and 15 N. schmidlii) tested positive in cPCR assays targeting a 710 base pair fragment of the cox-1 gene. Additionally, all 41 I. simplex tested positive in cPCR assays targeting a 460 base pair fragment of the 16S rRNA gene. All these ectoparasite DNA samples tested negative in PCR assays for Anaplasmataceae agents, Rickettsia spp., and piroplasmids.
The screening for
Bartonella DNA using qPCR for the 16S–23S intergenic transcribed spacer region identified 105 positive samples, of which 15/30 (50%) pools of
S. myoti and 2/5 (40%) specimens of
P. conspicua were obtained from males of the species
M. schreibersii collected in cave A (
Table 4); 32/61 (52.40%) pools of
S. myoti, 11/25 (44%) specimens of
P. conspicua, 2/7 (28.57%) specimens of
N. schmidlii, and 2/28 (14.28%) specimens of
I. simplex were obtained from females of the species
M. schreibersii collected in cave A (
Table 5); 22/73 (30.13%) pools of
S. myoti, 6/16 (37.50%) specimens of
P. conspicua, and 1/15 (2.22%) specimen of
I. simplex were obtained from males of the species
M. schreibersii collected in cave B (
Table 6); and 8/29 (27.58%) pools of
S. myoti and 2/3 (66.67%) specimens of
P. conspicua were obtained from females of the species
M. schreibersii collected in cave B (
Table 7).
Figure 3 illustrates the relationships between male and female bats of the species
M. schreibersii sampled from two caves in Portugal, their ectoparasites, and their association with
Bartonella.
3.4. Molecular Characterization of Bartonella spp.
Out of 105 ectoparasite samples that tested positive for
Bartonella spp. in the qPCR targeting the 16S–23S intergenic transcribed spacer region, 15 samples (3
P. conspicua, 3
N. schmidlii, 6
S. myoti, and 3
I. simplex) with the lowest Cq values (25–26.5) were selected for direct sequencing; however, this approach yielded unreadable sequences. Consequently, six
gltA readable sequences were obtained: one from
S. myoti collected from female bats in cave A (GS 106); two from
N. schmidlii collected from female bats in cave A (GS10 and GS13); and three from
S. myoti, two of which were collected from female bats and one collected from a male bat in cave B (GS53, GS55, and GS62). Readable sequences corresponding to positive samples for the
gltA gene are represented in
Table 5,
Table 6 and
Table 7 with blue diamonds.
The four Bartonella gltA sequences from S. myoti were identical to each other, constituting a single genotype. On the other hand, the other two sequences (from N. schmidlii) corresponded to two distinct genotypes. These three sequences were deposited in the GenBank database (accession numbers: PQ835041–PQ835043).
When comparing the sequences available in GenBank, the S. myoti-associated Bartonella gltA genotype obtained herein showed 100% identity (E-value: 5 × 10−169; query cover: 89%) to Bartonella sp. (MK140192), which was detected in P. conspicua collected from M. schreibersii in Hungary.
Additionally, one of the obtained Bartonella gltA sequences detected in N. schmidlii was 100% (E-value: 0.0 to 5 × 10−169; query cover: 77 to 98%) identical to Bartonella spp. (MW007702-11, MK140343-48, LC461055, MK140254, MK140349, MK140255, MK140259-60) detected in Miniopterus natalensis and associated bat flies (Nycteribia schmidlii schmidlii and Nycteribia schmidlii scottii) from Romania and South Africa, respectively; in Eptesicus serotinus, Miniopterus schreibersii, and Myotis blythii from Georgia; and in Eucampsipoda africana from Zambia.
The other gltA sequence detected in N. schmidlii was 100% (E-value: 0.0 to 9 × 10−172; query cover: 81 to 100%) identical to Bartonella sp. (KT751152, KT751155, KY679154, MK140283, MK140286, MK140353-54) detected in Miniopterus schreibersii from Georgia; P. conspicua and N. schmidlii from Romania; and Nycteribia stylidiopsis from Madagascar.
3.5. Phylogenetic Analyses
The Maximum Likelihood (ML) phylogenetic analysis, based on a 355 bp alignment of the
gltA gene and implemented with the GTR + I + G evolutionary model, positioned the genotype detected in
S. myoti into a clade with other
Bartonella sequences from bats in Europe and China, as well as their dipteran ectoparasites, with a bootstrap of 100% (
Figure 4). This clade is sister to a clade containing
Bartonella spp. sequences detected in
S. myoti from Poland and China, as well as in bats from Georgia and China. Both clades were located close to the clade of
B. henselae,
B. koehlerae, and
B. quintana. On the other hand, the sequences detected in
N. schmidlii were positioned in two different clades, both of which are composed of
Bartonella genotypes detected in
Miniopterus bat species and their respective dipteran ectoparasites collected from different regions of Europe and Africa (
Figure 4).
4. Discussion
This study provides novel insights into the molecular detection of
Bartonella spp. in ectoparasites associated with cave-dwelling bats from Portugal, while highlighting the absence of
Ehrlichia spp.,
Anaplasma spp.,
Rickettsia spp., and
Babesia/
Theileria spp. Notably, the
Bartonella genotypes detected in
S. myoti and
N. schmidlii expand the known geographic and host distribution of these bacteria. Although these findings align with previous studies conducted in Europe [
23,
24,
25,
66,
67], they represent the first molecular evidence of these genotypes in bat ectoparasites from Portugal, emphasizing the need for further surveys of pathogen diversity in this country. The absence of other vector-borne pathogens, such as
Rickettsia spp., which are commonly associated with bat ectoparasites in studies from Eastern Europe (Poland and Hungary) [
24,
28,
29], raises questions about regional or ecological factors that may influence pathogen presence in these ectoparasites.
The two obligate blood-feeding bat fly species,
Nycteribia schmidlii and
Penicillidia conspicua, found in our study are classified as oligoxenous since they were found on a restricted number of bat species (16 and 13, respectively), including
Miniopterus schreibersii, which represents a quite common host–parasite interaction [
28]. Interestingly, these two bat fly species are associated with
R. euryale,
R. mehelyi,
R. ferrumequinum, and
M. schreibersii, all of which share similar roosting ecology [
33].
The observed difference in the abundance of
I. simplex between male and female bats from cave A suggests that host biology, including sex-specific behaviors or physiological traits, may influence ectoparasite load and, consequently, the likelihood of pathogen transmission (e.g., [
68,
69,
70,
71,
72]). In contrast, no statistically significant difference was found for other ectoparasite species or in cave B. Identifying patterns in host–parasite interactions in natural environments is inherently challenging due to the multitude of variables that may exert influence. Biological and ecological factors related to both the host (e.g., age, reproductive stage) and the parasite (e.g., developmental stage, dispersal strategy) play a crucial role [
69,
70,
73]. Additionally, cave-specific environmental characteristics, such as microclimate and roosting density, may further shape these dynamics. Given these complexities, future studies should adopt a multifactorial approach that integrates both biotic and abiotic data to provide a more comprehensive understanding of these intricate host–parasite relationships.
Spinturnix myoti was the only spinturnicid mite species found among the bats sampled in this study. Indeed, the genus
Spinturnix is the most abundant and widespread genus within the family Spinturnicidae.
Spinturnix myoti has a broad distribution in the Palearctic region, including Portugal [
74], and shows a strong parasitic preference for bats of the
Myotis genus. Among these,
M. schreibersii is the only bat species from the family Miniopteridae that has been recorded as a host for
S. myoti [
56], which corroborates the results found herein. Additionally, this mite species has also been found parasitizing other bat genera within the Vespertilionidae and Rhinolophidae families [
74]. This finding highlights the adaptability of this mite species, demonstrating its ability to infest a variety of bat species across different families, further illustrating the ecological complexity of its parasitism. The high prevalence of
S.
myoti parasitizing bats in the studied caves, together with the significant association with
Bartonella spp., in addition to the wide distribution and adaptability of this mite species to several bat species, may favor its role in the transmission of bat-associated pathogens. The real role of Spinturnicidae mites in the epidemiology of bat-associated
Bartonella spp. should be further explored.
Ecological interactions between bats and their ectoparasites are inherently complex, with host behavior, roosting patterns, and parasite lifecycles all contributing to observed patterns of infestation and pathogen transmission. The colonial habits of
M. schreibersii [
69], which lead to high levels of social interaction and proximity, likely facilitate the spread and maintenance of ectoparasites such as
S. myoti and
N. schmidlii. Importantly, the focal bat species,
M. schreibersii, is classified as Vulnerable by the International Union for Conservation of Nature (IUCN) [
75] and faces additional challenges, such as habitat loss and anthropogenic pressures. This means that understanding the pathogens carried by this species can provide valuable insights into their health and survival threats.
The present study identified three
Bartonella genotypes; while one of these included all four sequences detected in
S. myoti obtained from bats captured in two different caves, the other two genotypes each comprised a single sequence detected in
N. schmidlii collected from bats in the same cave. Previous studies have identified a wide variety of
Bartonella gltA genotypes in Nycteribiidae bat flies in Europe, Asia, and Africa [
72,
76,
77,
78], which supports our findings. Despite the small number of sequences obtained from DNA samples extracted from flies, each sequence formed a distinct genotype. Strikingly, such diversity was not observed in the sequences obtained from
S. myoti, contrary to the findings of [
24], who detected three distinct
gltA genotypes of
Bartonella spp. in
S. myoti collected from bats in Poland. These authors reported that genotype A consisted of 18 sequences, while the other two genotypes each contained only one sequence. The number of sequences obtained in the present study was probably not enough to reveal the full diversity of genotypes in this mite species.
The large diversity of
Bartonella spp. circulating in bats and associated ectoparasites is clearly evidenced by phylogenetic studies, which reveal numerous clades formed by sequences detected in these animals, distributed throughout the phylogenetic tree [
66,
67,
72,
76,
79,
80,
81,
82]. In this study, the phylogenetic analysis grouped the sequences into three distinct clades, all containing sequences previously detected in bats and associated ectoparasites. The clade containing the sequence detected in
S. myoti was a sister clade to the one containing sequences detected in the same host from Poland and China, suggesting that this might represent a novel strain circulating in these mites in Portugal.
Herein, all
I. simplex collected from cave-dwelling bats tested negative in the qPCR for
Bartonella spp. Previously,
Bartonella spp. DNA was detected in 4% of
I. vespertilionis and 11% of
I. ariadnae collected from bats in Hungary and Romania [
28]. Recently, Szentiványi et al. [
82] detected
Bartonella sp. DNA in one male specimen of
I. vespertilionis collected in Eastern Europe. According to the authors, although the
Bartonella DNA might represent remnant DNA from the tick’s previous blood meal on its bat host (
Rhinolophus ferrumequinum), the vectorial competence of ticks in the transmission of
Bartonella spp. cannot be ruled out [
82].



 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}