

Eicosapentaenoic Acid and Docosahexaenoic Acid as an Antimicrobial Agent in Orthopedics—An In Vitro Study About the Race for Surface

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Substances
2.2. Cell Culture
2.3. Co-Culture
2.4. Scanning Electron Microscopy
2.5. Statistics
3. Results
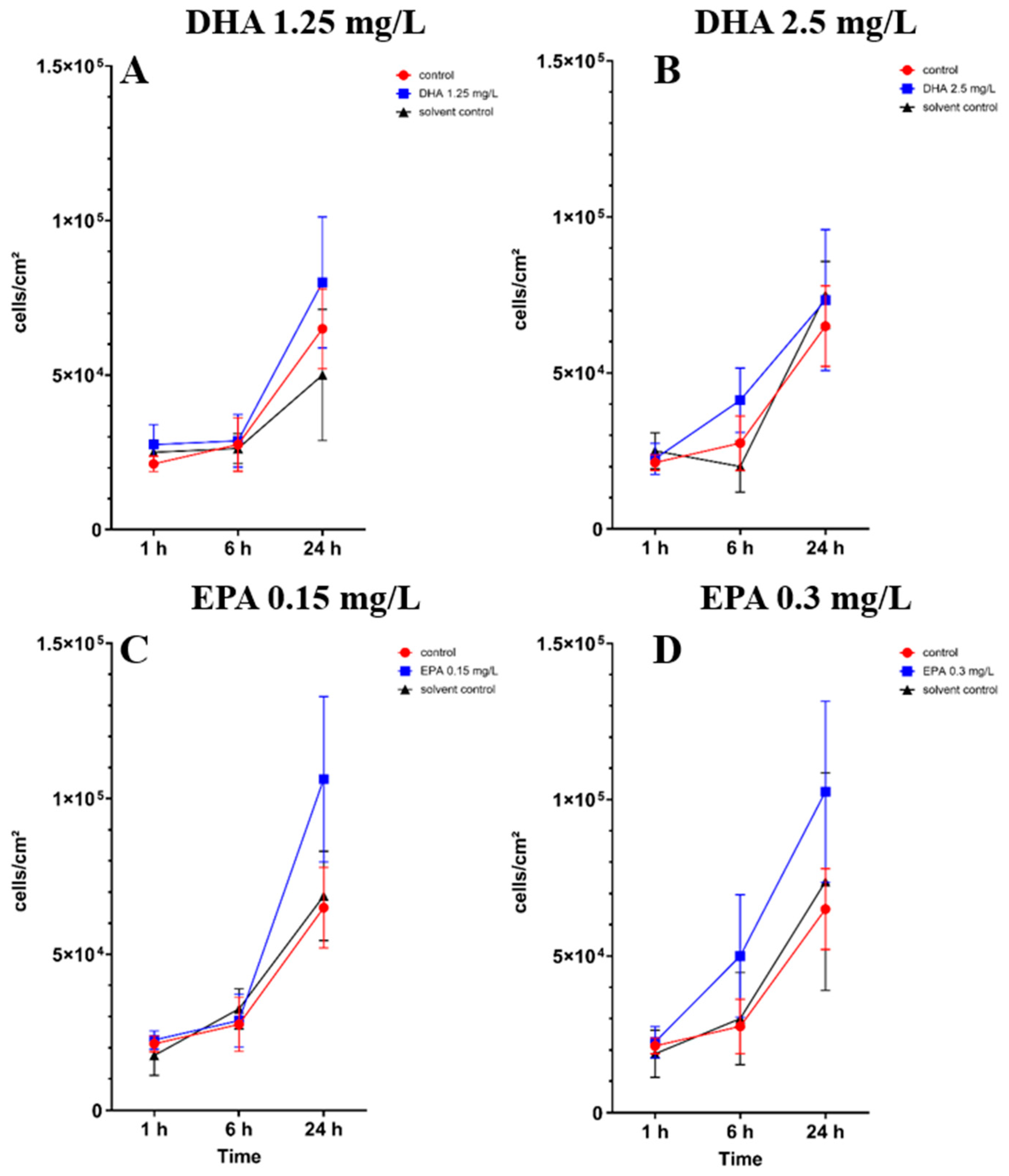
3.1. Influence of Omega-3 Fatty Acids on Osteoblast Cell Culture
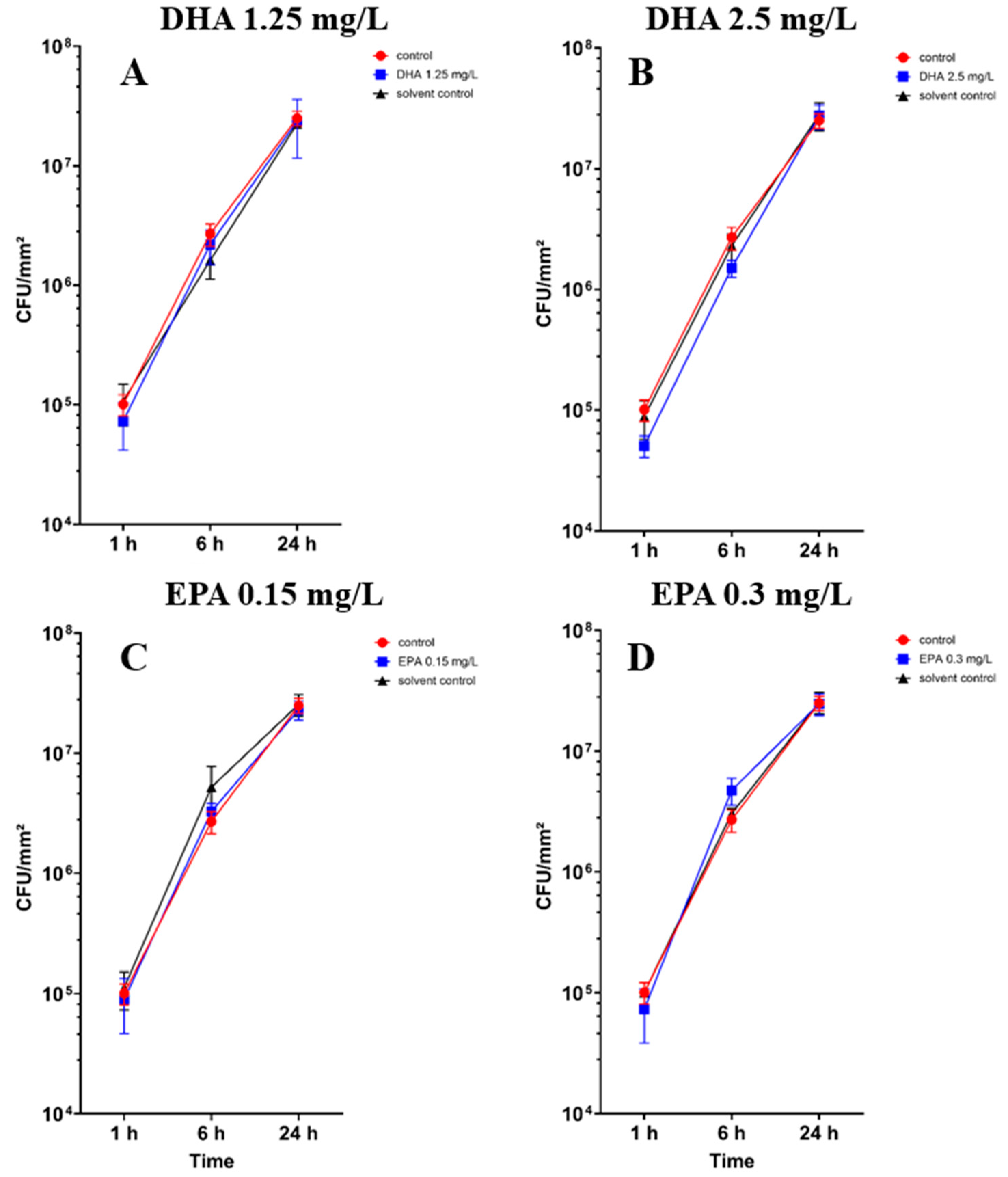
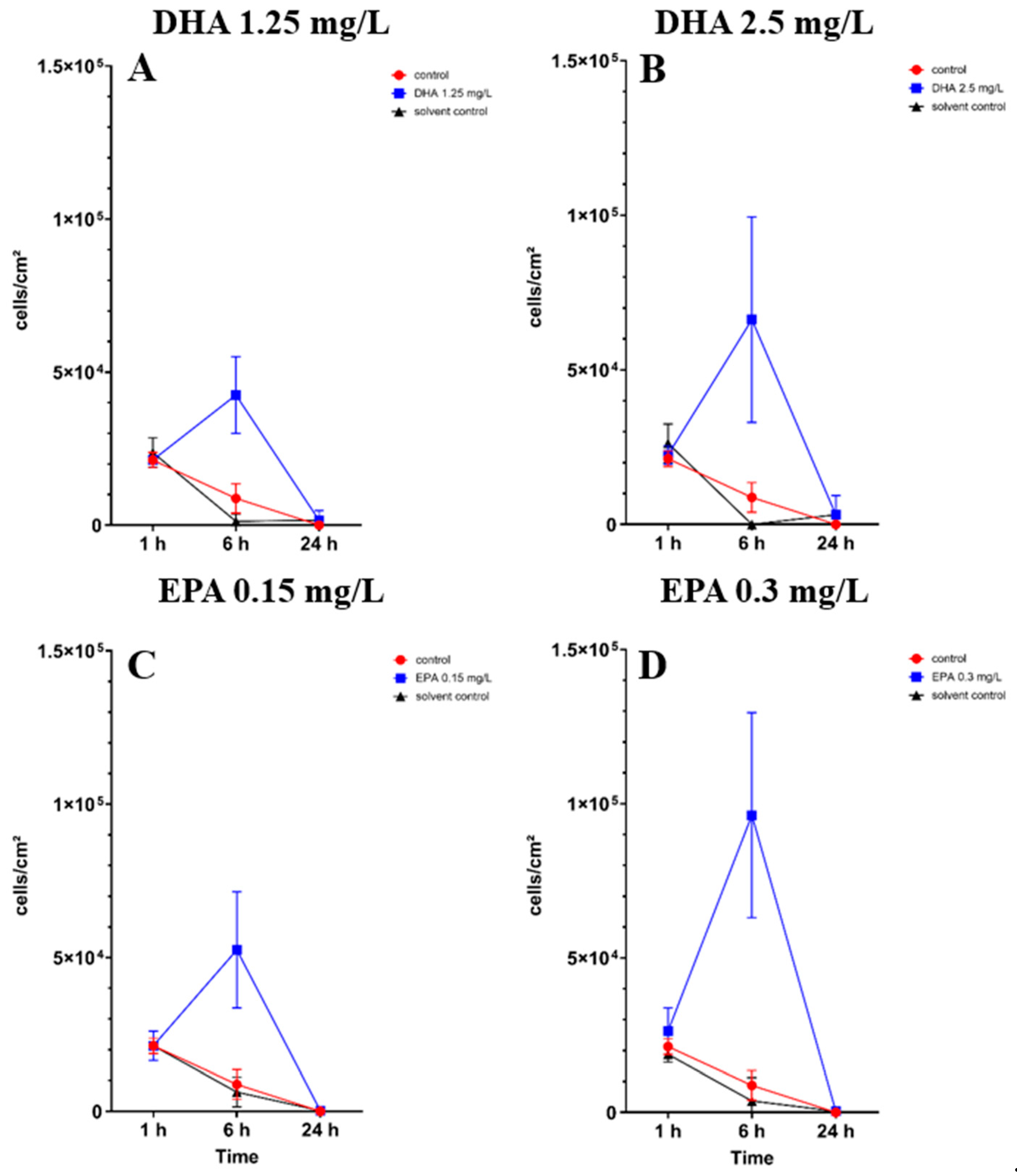
3.2. Influence of PUFAs on Co-Culture of Osteoblasts and Staphylococcus aureus
3.3. Scanning Electron Microscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ahmed, S.S.; Begum, F.; Kayani, B.; Haddad, F.S. Risk factors, diagnosis and management of prosthetic joint infection after total hip arthroplasty. Expert Rev. Med. Devices 2019, 16, 1063–1070. [Google Scholar] [CrossRef]
- Jin, X.; Luxan, B.G.; Hanly, M.; Pratt, N.L.; Harris, I.; de Steiger, R.; Graves, S.E.; Jorm, L. Estimating incidence rates of periprosthetic joint infection after hip and knee arthroplasty for osteoarthritis using linked registry and administrative health data. Bone Jt. J. 2022, 104, 1060–1066. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Luo, Y.; Hu, H.; Wang, W.; Zhao, Y.; Wang, S.; Zheng, Q.; Zhang, T.; Zhang, G.; Li, J.; et al. Clinically applicable optimized periprosthetic joint infection diagnosis via AI based pathology. NPJ Digit. Med. 2024, 7, 303. [Google Scholar] [CrossRef] [PubMed]
- Natsuhara, K.M.; Shelton, T.J.; Meehan, J.P.; Lum, Z.C. Mortality During Total Hip Periprosthetic Joint Infection. J. Arthroplast. 2019, 34, S337–S342. [Google Scholar] [CrossRef]
- Okike, K.; Bhattacharyya, T. Trends in the management of open fractures. A critical analysis. JBJS 2006, 88, 2739–2748. [Google Scholar] [CrossRef]
- Arciola, C.R.; Campoccia, D.; Montanaro, L. Implant infections: Adhesion, biofilm formation and immune evasion. Nat. Rev. Microbiol. 2018, 16, 397–409. [Google Scholar] [CrossRef]
- Kurtz, S.M.; Higgs, G.B.; Lau, E.; Iorio, R.R.; Courtney, P.M.; Parvizi, J. Hospital Costs for Unsuccessful Two-Stage Revisions for Periprosthetic Joint Infection. J. Arthroplast. 2022, 37, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Arciola, C.R.; Baldassarri, L.; Montanaro, L. In catheter infections by Staphylococcus epidermidis the intercellular adhesion (ica) locus is a molecular marker of the virulent slime-producing strains. J. Biomed. Mater. Res. 2002, 59, 557–562. [Google Scholar] [CrossRef]
- Arciola, C.R.; Campoccia, D.; Ravaioli, S.; Montanaro, L. Polysaccharide intercellular adhesin in biofilm: Structural and regulatory aspects. Front. Cell. Infect. Microbiol. 2015, 5, 7. [Google Scholar] [CrossRef]
- Campoccia, D.; Baldassarri, L.; Pirini, V.; Ravaioli, S.; Montanaro, L.; Arciola, C.R. Molecular epidemiology of Staphylococcus aureus from implant orthopaedic infections: Ribotypes, agr polymorphism, leukocidal toxins and antibiotic resistance. Biomaterials 2008, 29, 4108–4116. [Google Scholar] [CrossRef]
- Montanaro, L.; Poggi, A.; Visai, L.; Ravaioli, S.; Campoccia, D.; Speziale, P.; Arciola, C.R. Extracellular DNA in Biofilms. Int. J. Artif. Organs 2011, 34, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Rachid, S.; Cho, S.; Ohlsen, K.; Hacker, J.; Ziebuhr, W. Induction of Staphylococcus epidermidis biofilm formation by environmental factors: The possible involvement of the alternative transcription factor sigB. Adv. Exp. Med. Biol. 2000, 485, 159–166. [Google Scholar]
- Gristina, A.G.; Naylor, P.; Myrvik, Q. Infections from biomaterials and implants: A race for the surface. Med. Prog. Technol. 1988, 14, 205–224. [Google Scholar] [PubMed]
- Hickok, N.J.; Li, B.; Oral, E.; Zaat, S.A.J.; Armbruster, D.A.; Atkins, G.J.; Chen, A.F.; Coraça-Huber, D.C.; Dai, T.; Greenfield, E.M.; et al. The 2023 Orthopedic Research Society’s international consensus meeting on musculoskeletal infection: Summary from the in vitro section. J. Orthop. Res. 2024, 42, 512–517. [Google Scholar] [CrossRef]
- Coraça-Huber, D.C.; Steixner, S.; Wurm, A.; Nogler, M. Antibacterial and Anti-Biofilm Activity of Omega-3 Polyunsaturated Fatty Acids against Periprosthetic Joint Infections-Isolated Multi-Drug Resistant Strains. Biomedicines 2021, 9, 334. [Google Scholar] [CrossRef]
- Kim, Y.-G.; Lee, J.-H.; Raorane, C.J.; Oh, S.T.; Park, J.G.; Lee, J. Herring Oil and Omega Fatty Acids Inhibit Staphylococcus aureus Biofilm Formation and Virulence. Front. Microbiol. 2018, 9, 1241. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, C.; Steixner, S.J.M.; Coraça-Huber, D.C. Antibiofilm Activity of Omega-3 Fatty Acids and Its Influence on the Expression of Biofilm Formation Genes on Staphylococcus aureus. Antibiotics 2022, 11, 932. [Google Scholar] [CrossRef] [PubMed]
- Levental, K.R.; Surma, M.A.; Skinkle, A.D.; Lorent, J.H.; Zhou, Y.; Klose, C.; Chang, J.T.; Hancock, J.F.; Levental, I. ω-3 polyunsaturated fatty acids direct differentiation of the membrane phenotype in mesenchymal stem cells to potentiate osteogenesis. Sci. Adv. 2017, 3, eaao1193. [Google Scholar] [CrossRef]
- Agarwal, A.; Jain, A. Glucose & sodium chloride induced biofilm production & ica operon in clinical isolates of staphylococci. Indian J. Med. Res. 2013, 138, 262–266. [Google Scholar] [PubMed]
- Steixner, S.J.M.; Spiegel, C.; Dammerer, D.; Wurm, A.; Nogler, M.; Coraça-Huber, D.C. Influence of Nutrient Media Compared to Human Synovial Fluid on the Antibiotic Susceptibility and Biofilm Gene Expression of Coagulase-Negative Staphylococci In Vitro. Antibiotics 2021, 10, 790. [Google Scholar] [CrossRef]
- Parente, R.; Possetti, V.; Schiavone, M.L.; Campodoni, E.; Menale, C.; Loppini, M.; Doni, A.; Bottazzi, B.; Mantovani, A.; Sandri, M.; et al. 3D Cocultures of Osteoblasts and Staphylococcus aureus on Biomimetic Bone Scaffolds as a Tool to Investigate the Host–Pathogen Interface in Osteomyelitis. Pathogens 2021, 10, 837. [Google Scholar] [CrossRef]
- Stracquadanio, S.; Musso, N.; Costantino, A.; Lazzaro, L.M.; Stefani, S.; Bongiorno, D. Internalization in Osteoblast Cells: Mechanisms, Interactions and Biochemical Processes. What Did We Learn from Experimental Models? Pathogens 2021, 10, 239. [Google Scholar] [CrossRef] [PubMed]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and virulence of. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef]
- Gongadze, E.; Kabaso, D.; Bauer, S.; Slivnik, T.; Schmuki, P.; Rienen, U.V.; Iglič, A. Adhesion of osteoblasts to a nanorough titanium implant surface. Int. J. Nanomed. 2011, 6, 1801–1816. [Google Scholar]
- Houston, P.; Rowe, S.E.; Pozzi, C.; Waters, E.M.; O’Gara, J.P. Essential Role for the Major Autolysin in the Fibronectin-Binding Protein-Mediated Staphylococcus aureus Biofilm Phenotype. Infect. Immun. 2011, 79, 1153–1165. [Google Scholar] [CrossRef]
- Josse, J.; Laurent, F.; Diot, A. Staphylococcal Adhesion and Host Cell Invasion: Fibronectin-Binding and Other Mechanisms. Front. Microbiol. 2017, 8, 2433. [Google Scholar] [CrossRef]
- Hawkins, J.; Kodali, S.; Matsuka, Y.V.; McNeil, L.K.; Mininni, T.; Scully, I.L.; Vernachio, J.H.; Severina, E.; Girgenti, D.; Jansen, K.U.; et al. A Recombinant Clumping Factor A-Containing Vaccine Induces Functional Antibodies to Staphylococcus aureus That Are Not Observed after Natural Exposure. Clin. Vaccine Immunol. 2012, 19, 1641–1650. [Google Scholar] [CrossRef]
- Foster, T.J. The remarkably multifunctional fibronectin binding proteins of Staphylococcus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1923–1931. [Google Scholar] [CrossRef] [PubMed]
- Longo, A.B.; E Ward, W. PUFAs, Bone Mineral Density, and Fragility Fracture: Findings from Human Studies. Adv. Nutr. Int. Rev. J. 2016, 7, 299–312. [Google Scholar] [CrossRef]
- Hinojosa, C.A.; Gonzalez-Juarbe, N.; Rahman, M.; Fernandes, G.; Orihuela, C.J.; Restrepo, M.I. Omega-3 fatty acids in contrast to omega-6 protect against pneumococcal pneumonia. Microb. Pathog. 2020, 141, 103979. [Google Scholar] [CrossRef]
- Menni, C.; Zierer, J.; Pallister, T.; Jackson, M.A.; Long, T.; Mohney, R.P.; Steves, C.J.; Spector, T.D.; Valdes, A.M. Omega-3 Fatty Acids Correlate with Gut Microbiome Diversity and Production of N-Carbamylglutamate in Middle Aged and Elderly Women. Sci. Rep. 2017, 7, 11079. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters, version 11.0. 2021. Available online: https://www.eucast.org/ (accessed on 23 December 2024).
- Floros, S.; Toskas, A.; Pasidi, E.; Vareltzis, P. Bioaccessibility and Oxidative Stability of Omega-3 Fatty Acids in Supplements, Sardines and Enriched Eggs Studied Using a Static In Vitro Gastrointestinal Model. Molecules 2022, 27, 415. [Google Scholar] [CrossRef]
- Carlson, S.J.; Fallon, E.M.; Kalish, B.T.; Gura, K.M.; Puder, M. The role of the ω-3 fatty acid DHA in the human life cycle. JPEN J. Parenter. Enter. Nutr. 2013, 37, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.D.; Nessler, J.M.; Horazdovsky, R.D.; Vang, S.; Thomas, A.J.; Marston, S.B. Serum and Wound Vancomycin Levels After Intrawound Administration in Primary Total Joint Arthroplasty. J. Arthroplast. 2017, 32, 924–928. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spiegel, C.; Ünalan, B.; Kaserbacher, A.; Arora, R.; Coraça-Huber, D.C. Eicosapentaenoic Acid and Docosahexaenoic Acid as an Antimicrobial Agent in Orthopedics—An In Vitro Study About the Race for Surface. Pathogens 2025, 14, 57. https://doi.org/10.3390/pathogens14010057
Spiegel C, Ünalan B, Kaserbacher A, Arora R, Coraça-Huber DC. Eicosapentaenoic Acid and Docosahexaenoic Acid as an Antimicrobial Agent in Orthopedics—An In Vitro Study About the Race for Surface. Pathogens. 2025; 14(1):57. https://doi.org/10.3390/pathogens14010057
Chicago/Turabian StyleSpiegel, Christopher, Burak Ünalan, Andreas Kaserbacher, Rohit Arora, and Débora C. Coraça-Huber. 2025. "Eicosapentaenoic Acid and Docosahexaenoic Acid as an Antimicrobial Agent in Orthopedics—An In Vitro Study About the Race for Surface" Pathogens 14, no. 1: 57. https://doi.org/10.3390/pathogens14010057
APA StyleSpiegel, C., Ünalan, B., Kaserbacher, A., Arora, R., & Coraça-Huber, D. C. (2025). Eicosapentaenoic Acid and Docosahexaenoic Acid as an Antimicrobial Agent in Orthopedics—An In Vitro Study About the Race for Surface. Pathogens, 14(1), 57. https://doi.org/10.3390/pathogens14010057