
Diversity of the Amoebozoa and Ciliophora Groups in Non-Human Primates Kept Ex Situ and in Their Handlers in Different Institutions in Brazil

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Fecal Samples Collection
2.2. Microscopic Parasitological Techniques
2.3. In Vitro Protist Cultivation
2.4. Wheatley Trichrome Staining—Amoebozoa Group
2.5. Differential Interference Contrast (DIC)—Ciliophora Group
2.6. Molecular Characterization
3. Results
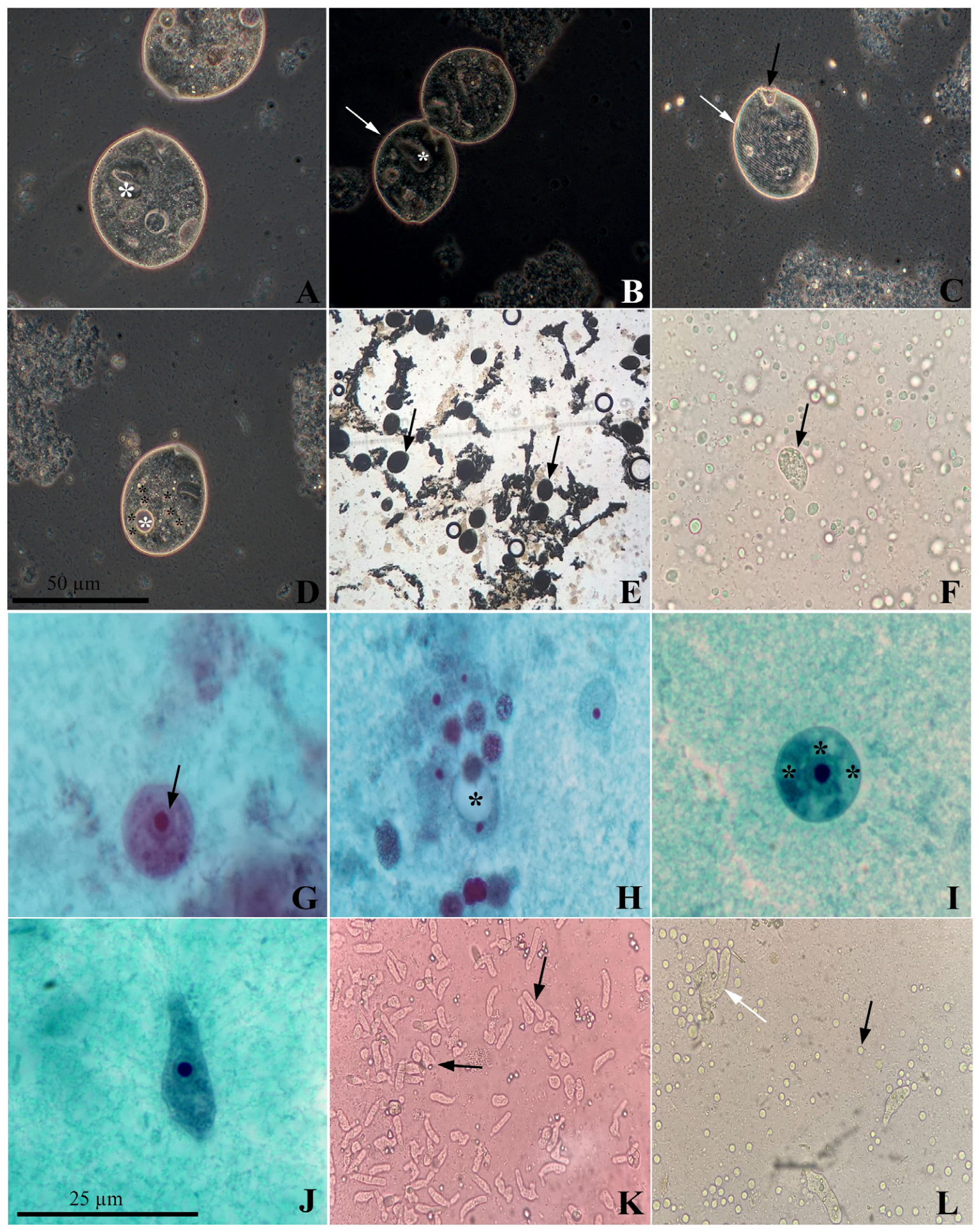
3.1. Microscopic Analysis and Protists’ In Vitro Isolation
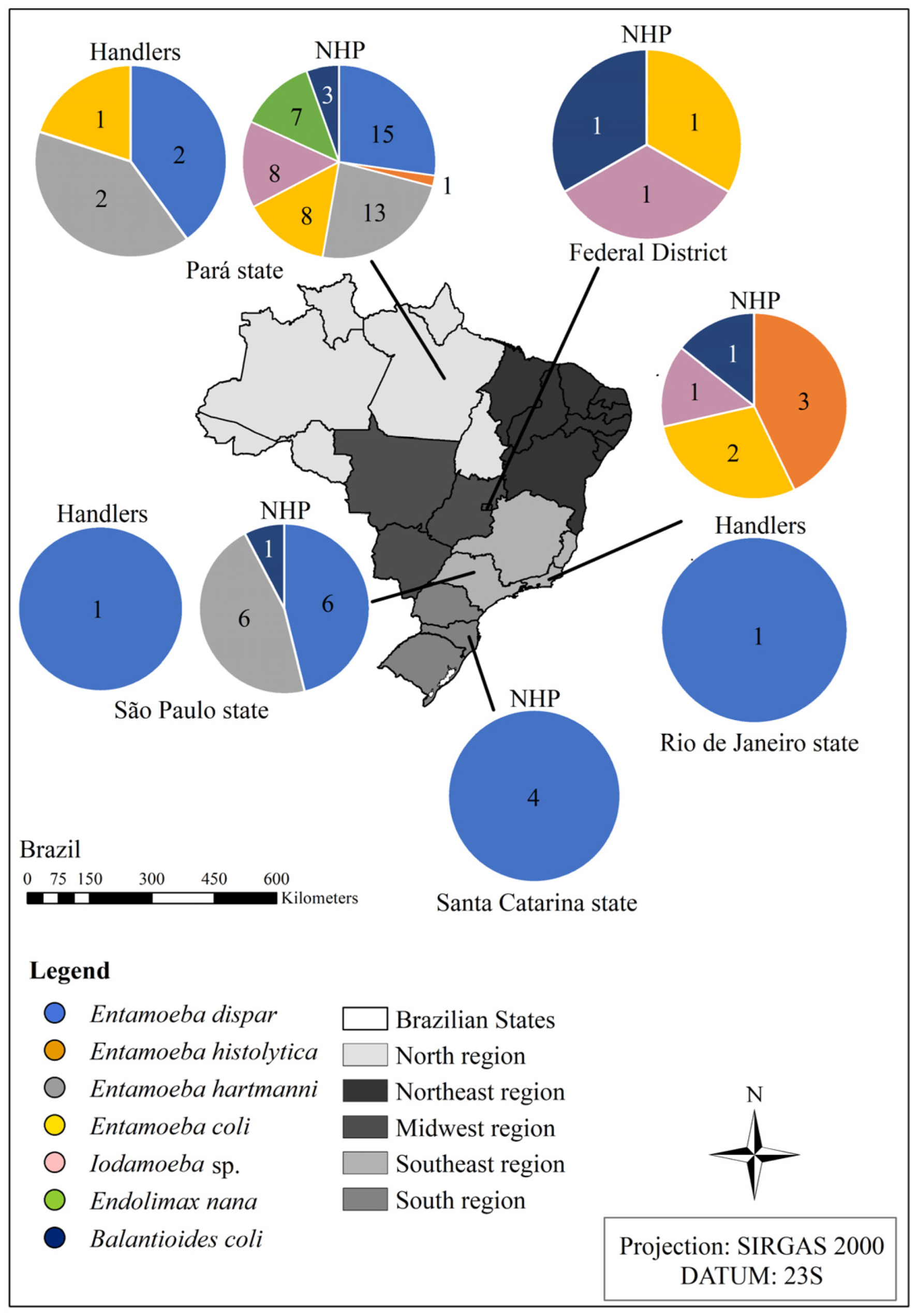
3.2. Protists’ Molecular Characterization and Distribution in Brazil
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Levecke, B.; Dorny, P.; Geurden, T.; Vercammen, F.; Vercruysse, J. Gastrointestinal Protozoa in Non-Human Primates of Four Zoological Gardens in Belgium. Vet. Parasitol. 2007, 148, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Vonfeld, I.; Prenant, T.; Polack, B.; Guillot, J.; Quintard, B. Gastrointestinal Parasites in Non-Human Primates in Zoological Institutions in France. Parasite 2022, 29, 43. [Google Scholar] [CrossRef] [PubMed]
- Haque, R.; Houston, C.; Hughes, M.; Houpt, E.; Petri, W., Jr. Amebiasis. N. Engl. J. Med. 2003, 348, 8. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.K.M.; Hossain, M.B.; Roy, S.; Ayeh-Kumi, P.F.; Petri, W.A.; Haque, R.; Clark, C.G. Entamoeba moshkovskii Infections in Children in Bangladesh. Emerg. Infect. Dis. 2003, 9, 580–584. [Google Scholar] [CrossRef]
- Calegar, D.A.; Nunes, B.C.; Monteiro, K.J.L.; Santos, J.P.D.; Toma, H.K.; Gomes, T.F.; Lima, M.M.; Bóia, M.N.; Carvalho-Costa, F.A. Frequency and Molecular Characterisation of Entamoeba histolytica, Entamoeba dispar, Entamoeba moshkovskii, and Entamoeba hartmanni in the Context of Water Scarcity in Northeastern Brazil. Mem. Inst. Oswaldo Cruz 2016, 111, 114–119. [Google Scholar] [CrossRef]
- Levecke, B.; Dorny, P.; Vercammen, F.; Visser, L.G.; Van Esbroeck, M.; Vercruysse, J.; Verweij, J.J. Transmission of Entamoeba nuttalli and Trichuris trichiura from Nonhuman Primates to Humans. Emerg. Infect. Dis. 2015, 21, 1871–1872. [Google Scholar] [CrossRef]
- Pomajbíková, K.; Oborník, M.; Horák, A.; Petrželková, K.J.; Grim, J.N.; Levecke, B.; Todd, A.; Mulama, M.; Kiyang, J.; Modrý, D. Novel Insights into the Genetic Diversity of Balantidium and Balantidium-like Cyst-Forming Ciliates. PLoS Negl. Trop. Dis. 2013, 7, e2140. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Ascuña-Durand, K.; Chihi, A.; Belkessa, S.; Kurt, Ö.; El-Badry, A.; Van Der Giezen, M.; Clark, C.G. Further Insight into the Genetic Diversity of Entamoeba coli and Entamoeba hartmanni. J. Eukaryot. Microbiol. 2023, 70, e12949. [Google Scholar] [CrossRef]
- Kantor, M.; Abrantes, A.; Estevez, A.; Schiller, A.; Torrent, J.; Gascon, J.; Hernandez, R.; Ochner, C. Entamoeba histolytica: Updates in Clinical Manifestation, Pathogenesis, and Vaccine Development. Can. J. Gastroenterol. Hepatol. 2018, 2018, 1–6. [Google Scholar] [CrossRef]
- Schuster, F.L.; Ramirez-Avila, L. Current World Status of Balantidium coli. Clin. Microbiol. Rev. 2008, 21, 626–638. [Google Scholar] [CrossRef]
- Barbosa, A.d.S.; Pissinatti, A.; Dib, L.V.; De Siqueira, M.P.; Cardozo, M.L.; Fonseca, A.B.M.; De Barros Oliveira, A.; Da Silva, F.A.; Uchôa, C.M.A.; Bastos, O.M.P.; et al. Balantidium coli and Other Gastrointestinal Parasites in Captives Non-human Primates of the Rio de Janeiro, Brazil. J. Med. Primatol. 2015, 44, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Arafa, W.M.; Mahrous, L.N.; Aboelhadid, S.M.; Abdel-Ghany, A.E. Investigation of Enteric Parasites of Zoo Animals and Zookeepers in Beni-Suef Governorate, Egypt. J. Vet. Med. Res. 2013, 22, 121–125. [Google Scholar] [CrossRef]
- Barbosa, A.D.S.; Barbosa, H.S.; Souza, S.M.D.O.; Dib, L.V.; Uchôa, C.M.A.; Bastos, O.M.P.; Amendoeira, M.R.R. Balantioides coli: Morphological and Ultrastructural Characteristics of Pig and Non-Human Primate Isolates. Acta Parasitol. 2018, 63, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.d.S.; Ponce-Gordo, F.; Dib, L.V.; Antunes Uchôa, C.M.; Bastos, O.M.P.; Pissinatti, A.; Amendoeira, M.R.R. First Molecular Characterization of Balantioides coli (Malmsten, 1857) Isolates Maintained In Vitro Culture and from Feces of Captive Animals, Rio de Janeiro, Brazil. Vet. Parasitol. Reg. Stud. Rep. 2017, 10, 102–113. [Google Scholar] [CrossRef]
- Feng, M.; Yang, B.; Yang, L.; Fu, Y.; Zhuang, Y.; Liang, L.; Xu, Q.; Cheng, X.; Tachibana, H. High Prevalence of Entamoeba Infections in Captive Long-Tailed Macaques in China. Parasitol. Res. 2011, 109, 1093–1097. [Google Scholar] [CrossRef]
- Nilles-Bije, M.L.; Rivera, W.L. Ultrastructural and Molecular Characterization of Balantidium coli Isolated in the Philippines. Parasitol. Res. 2010, 106, 387–394. [Google Scholar] [CrossRef]
- Tachibana, H.; Yanagi, T.; Akatsuka, A.; Kobayashi, S.; Kanbara, H.; Tsutsumi, V. Isolation and Characterization of a Potentially Virulent Species Entamoeba nuttalli from Captive Japanese Macaques. Parasitology 2009, 136, 1169–1177. [Google Scholar] [CrossRef]
- Takano, J.; Narita, T.; Tachibana, H.; Shimizu, T.; Komatsubara, H.; Terao, K.; Fujimoto, K. Entamoeba histolytica and Entamoeba dispar Infections in Cynomolgus Monkeys Imported into Japan for Research. Parasitol. Res. 2005, 97, 255–257. [Google Scholar] [CrossRef]
- Yan, W.; Wang, T.; Zhao, L.; Sun, C. Modified DMEM Xenic Culture Medium for Propagation, Isolation and Maintenance of Balantioides coli. Acta Trop. 2021, 214, 105762. [Google Scholar] [CrossRef]
- Barbosa, A.D.S.; Pinheiro, J.L.; Dos Santos, C.R.; De Lima, C.S.C.C.; Dib, L.V.; Echarte, G.V.; Augusto, A.M.; Bastos, A.C.M.P.; Antunes Uchôa, C.M.; Bastos, O.M.P.; et al. Gastrointestinal Parasites in Captive Animals at the Rio de Janeiro Zoo. Acta Parasit. 2020, 65, 237–249. [Google Scholar] [CrossRef]
- Dawet, A.; Yakubu, D.; Butu, H. Survey of Gastrointestinal Parasites of Non-Human Primates in Jos Zoological Garden. J. Primatol. 2013, 2, 1000108. [Google Scholar] [CrossRef]
- Hinde, K. Milk Composition Varies in Relation to the Presence and Abundance of Balantidium coli in the Mother in Captive Rhesus Macaques (Macaca mulatta). Am. J. Primatol. 2007, 69, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.A.L.; Ngui, R.; Shukri, J.; Rohela, M.; Mat Naim, H.R. Intestinal Parasites in Various Animals at a Zoo in Malaysia. Vet. Parasitol. 2008, 157, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Nakauchi, K. The Prevalence of Balantidium coli Infection in Fifty-Six Mammalian Species. J. Vet. Med. Sci. 1999, 61, 63–65. [Google Scholar] [CrossRef]
- Pérez Cordón, G.; Hitos Prados, A.; Romero, D.; Sánchez Moreno, M.; Pontes, A.; Osuna, A.; Rosales, M.J. Intestinal Parasitism in the Animals of the Zoological Garden “Peña Escrita” (Almuñecar, Spain). Vet. Parasitol. 2008, 156, 302–309. [Google Scholar] [CrossRef]
- Rondón, S.; Cavallero, S.; Montalbano Di Filippo, M.; De Liberato, C.; Berrilli, F.; Capitani, N.; D’Amelio, S. Intestinal Parasites Infecting Captive Non-Human Primates in Italy. Front. Vet. Sci. 2024, 10, 1270202. [Google Scholar] [CrossRef]
- Nunn, C.; Altizer, S. Infectious Diseases in Primates: Behavior, Ecology and Evolution, 1st ed.; Oxford University Press: Oxford, UK, 2006; ISBN 978-0-19-172823-5. [Google Scholar]
- Acosta-Jamett, G.; Chaves, A. Ecology of Wildlife Diseases in the Neotropics, 1st ed.; Springer: Cham, Switzerland, 2024; ISBN 978-3-031-50531-7. [Google Scholar]
- Amato, K.R.; Sanders, J.G.; Song, S.J.; Nute, M.; Metcalf, J.L.; Thompson, L.R.; Morton, J.T.; Amir, A.; McKenzie, V.J.; Humphrey, G.; et al. Evolutionary Trends in Host Physiology Outweigh Dietary Niche in Structuring Primate Gut Microbiomes. ISME J. 2019, 13, 576–587. [Google Scholar] [CrossRef]
- Dougherty, E.R.; Carlson, C.J.; Bueno, V.M.; Burgio, K.R.; Cizauskas, C.A.; Clements, C.F.; Seidel, D.P.; Harris, N.C. Paradigms for Parasite Conservation. Conserv. Biol. 2016, 30, 724–733. [Google Scholar] [CrossRef]
- Verona, C.E.; Pissinatti, A. Primatas Do Novo Mundo (Sagui, Macaco-Prego, Macaco-Aranha, Bugio e Muruqui), 2nd ed.; Roca: São Paulo, Brazil, 2014. [Google Scholar]
- Dib, L.V.; Barbosa, A.; Da Silva, B.; Pissinatti, A.; Moreira, S.; Tavares, M.C.; Teixeira, R.; Da Costa, A.L.; Muniz, J.A.; Junglos, A.; et al. Gastrointestinal Parasites Affecting Non-Human Primates That Are Kept Ex Situ and Their Handlers in Different Brazilian Institutions: Diagnosis and Analysis of Risk Factors. Pathogens 2023, 12, 1410. [Google Scholar] [CrossRef]
- Faust, E.; D’Antoni, J.; Miller, V.; Sawitz, C.; Thomen, L.; Tobie, J.; Walker, J. A Critical Study of Clinical Laboratory Technics for the Diagnosis of Protozoan Cysts and Helminth Eggs in Feces. Am. J. Trop. Med. Hyg. 1938, 18, 14. [Google Scholar] [CrossRef]
- Ritchie, L. An Ether Sedimentation Technique for Routine Stool Examinations. Bull. U S Army Med. Dep. 1948, 8, 1. [Google Scholar]
- Young, K.H.; Bullock, S.L.; Melvin, D.M.; Spruill, C.L. Ethyl Acetate as a Substitute for Diethyl Ether in the Formalin-Ether Sedimentation Technique. J. Clin. Microbiol. 1979, 10, 852–853. [Google Scholar] [CrossRef] [PubMed]
- Lutz, A. Schistosomum mansoni and Schistosomatosis Observed in Brazil. Mem. Inst. Oswaldo Cruz 1919, 11, 30. [Google Scholar] [CrossRef]
- Jones, W.R. The Experimental Infection of Rats with Entamoeba histolytica; with a Method for Evaluating the Anti-Amoebic Properties of New Compounds. Ann. Trop. Med. Parasitol. 1946, 40, 130–140. [Google Scholar] [CrossRef]
- Pavlova, E. Sur Les Méthodes de La Culture d’Entamoeba histolytica. Parazitol. Med. 1938, 7, 4. [Google Scholar]
- Diamond, L.S. A New Liquid Medium for Xenic Cultivation of Entamoeba histolytica and Other Lumen-Dwelling Protozoa. J. Parasitol. 1982, 68, 958. [Google Scholar] [CrossRef]
- Paglia, M.G.; Visca, P. An Improved PCR-Based Method for Detection and Differentiation of Entamoeba histolytica and Entamoeba dispar in Formalin-Fixed Stools. Acta Trop. 2004, 92, 273–277. [Google Scholar] [CrossRef]
- Lau, Y.L.; Anthony, C.; Fakhrurrazi, S.A.; Ibrahim, J.; Ithoi, I.; Mahmud, R. Real-Time PCR Assay in Differentiating Entamoeba histolytica, Entamoeba dispar, and Entamoeba moshkovskii Infections in Orang Asli Settlements in Malaysia. Parasites Vectors 2013, 6, 250. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Lebbad, M.; Victory, E.L.; Verweij, J.J.; Tannich, E.; Alfellani, M.; Legarraga, P.; Clark, C.G. Increased Sampling Reveals Novel Lineages of Entamoeba: Consequences of Genetic Diversity and Host Specificity for Taxonomy and Molecular Detection. Protist 2011, 162, 525–541. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Winiecka-Krusnell, J.; Lier, T.; Lebbad, M. Evaluation of a PCR Method for Detection of Entamoeba polecki, with an Overview of Its Molecular Epidemiology. J. Clin. Microbiol. 2018, 56, e00154-18. [Google Scholar] [CrossRef]
- Chihi, A.; Stensvold, C.R.; Ben-abda, I.; Ben-Romdhane, R.; Aoun, K.; Siala, E.; Bouratbine, A. Development and Evaluation of Molecular Tools for Detecting and Differentiating Intestinal Amoebae in Healthy Individuals. Parasitology 2019, 146, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, H.; Yanagi, T.; Pandey, K.; Cheng, X.-J.; Kobayashi, S.; Sherchand, J.B.; Kanbara, H. An Entamoeba Sp. Strain Isolated from Rhesus Monkey Is Virulent but Genetically Different from Entamoeba histolytica. Mol. Biochem. Parasitol. 2007, 153, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Ponce-Gordo, F.; Fonseca-Salamanca, F.; Martínez-Díaz, R.A. Genetic Heterogeneity in Internal Transcribed Spacer Genes of Balantidium coli (Litostomatea, Ciliophora). Protist 2011, 162, 774–794. [Google Scholar] [CrossRef]
- Parija, S.C.; Khairnar, K. Entamoeba moshkovskii and Entamoeba dispar-Associated Infections in Pondicherry, India. J. Health Popul. Nutr. 2005, 23, 292–295. [Google Scholar]
- Rivera, W.L.; Yason, J.A.D.L.; Adao, D.E.V. Entamoeba histolytica and E. Dispar Infections in Captive Macaques (Macaca fascicularis) in the Philippines. Primates 2010, 51, 69–74. [Google Scholar] [CrossRef]
- Tachibana, H.; Cheng, X.-J.; Kobayashi, S.; Matsubayashi, N.; Gotoh, S.; Matsubayashi, K. High Prevalence of Infection with Entamoeba dispar, but Not E. histolytica, in Captive Macaques. Parasitol. Res. 2001, 87, 14–17. [Google Scholar] [CrossRef]
- Zanetti, A.d.S.; Malheiros, A.F.; De Matos, T.A.; Dos Santos, C.; Battaglini, P.F.; Moreira, L.M.; Lemos, L.M.S.; Castrillon, S.K.I.; Da Costa Boamorte Cortela, D.; Ignotti, E.; et al. Diversity, Geographical Distribution, and Prevalence of Entamoeba Spp. in Brazil: A Systematic Review and Meta-Analysis. Parasite 2021, 28, 17. [Google Scholar] [CrossRef]
- Graffeo, R.; Archibusacci, C.M.; Soldini, S.; Romano, L.; Masucci, L. Entamoeba dispar: A Rare Case of Enteritis in a Patient Living in a Nonendemic Area. Case Rep. Gastrointest. Med. 2014, 2014, 1–3. [Google Scholar] [CrossRef]
- Berrilli, F.; Prisco, C.; Friedrich, K.G.; Di Cerbo, P.; Di Cave, D.; De Liberato, C. Giardia Duodenalis Assemblages and Entamoeba Species Infecting Non-Human Primates in an Italian Zoological Garden: Zoonotic Potential and Management Traits. Parasites Vectors 2011, 4, 199. [Google Scholar] [CrossRef]
- Levecke, B.; Dreesen, L.; Dorny, P.; Verweij, J.J.; Vercammen, F.; Casaert, S.; Vercruysse, J.; Geldhof, P. Molecular Identification of Entamoeba Spp. in Captive Nonhuman Primates. J. Clin. Microbiol. 2010, 48, 2988–2990. [Google Scholar] [CrossRef]
- Regan, C.S.; Yon, L.; Hossain, M.; Elsheikha, H.M. Prevalence of Entamoeba Species in Captive Primates in Zoological Gardens in the UK. PeerJ 2014, 2, e492. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, R.; Böer, M.; Herder, V.; Spitzbarth, I.; Hewicker-Trautwein, M.; Baumgärtner, W.; Wohlsein, P. Epizootic Fatal Amebiasis in an Outdoor Group of Old World Monkeys: Entamoeba histolytica in Colobinae. J. Med. Primatol. 2010, 39, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Verweij, J.J.; Vermeer, J.; Brienen, E.A.T.; Blotkamp, C.; Laeijendecker, D.; Van Lieshout, L.; Polderman, A.M. Entamoeba histolytica Infections in Captive Primates. Parasitol. Res. 2003, 90, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Loomis, M.; Britt, O., Jr.; Gendron, A.; Howard, E. Hepatic and Gastric Amebiasis in Black and White Colobus Monkeys. J. Am. Vet. Med. Assoc. 1983, 183, 4. [Google Scholar]
- Palmieri, J.; Dalgard, D.; Connor, D. Gastric Amebiasis in a Silvered Leaf Monkey. J. Am. Vet. Med. Assoc. 1984, 185, 2. [Google Scholar]
- Wallach, J.D.; Boever, W.J. Diseases of Exotic Animals. Medical and Surgical Management; W.B. Saunders Co.: Philadelphia, PA, USA, 1983; ISBN 0-7216-9105-6. [Google Scholar]
- Menu, E.; Davoust, B.; Mediannikov, O.; Akiana, J.; Mulot, B.; Diatta, G.; Levasseur, A.; Ranque, S.; Raoult, D.; Bittar, F. Occurrence of Ten Protozoan Enteric Pathogens in Three Non-Human Primate Populations. Pathogens 2021, 10, 280. [Google Scholar] [CrossRef]
- Pomajbíková, K.; Petrželková, K.J.; Profousová, I.; Petrášová, J.; Modrý, D. Discrepancies in the Occurrence of Balantidium coli Between Wild and Captive African Great Apes. J. Parasitol. 2010, 96, 1139–1144. [Google Scholar] [CrossRef]
- Sá, L.C.E.F.; Silva, E.M.L.D.; Morais, C.V.F.; Castro, L.F.D.; Rocha, C.R.R.; Maldonade, I.R.; Machado, E.R. Prevalence of Enteroparasites in Primates Kept at the Brasília Zoo, Brazil. Res. Soc. Dev. 2023, 12, e27112541739. [Google Scholar] [CrossRef]
- Al-Tayib, O.; Abdoun, K.A. Balantidium coli Infection in Hamadryas Baboon (Papio hamadryas) in Saudi Arabia: A Case Report. J. Anim. Plant Sci. 2013, 23, 4. [Google Scholar]
- Kim, J.C.S.; Abee, C.R.; Wolf, R.H. Balantidiosis in a Chimpanzee (Pan troglodytes). Lab. Anim. 1978, 12, 231–233. [Google Scholar] [CrossRef]
- Lankester, F.; Mätz-Rensing, K.; Kiyang, J.; Jensen, S.A.; Weiss, S.; Leendertz, F.H. Fatal Ulcerative Colitis in a Western Lowland Gorilla (Gorilla gorilla gorilla). J. Med. Primatol. 2008, 37, 080505182215573. [Google Scholar] [CrossRef]
- Calegar, D.A.; Monteiro, K.J.L.; Bacelar, P.A.A.; Evangelista, B.B.C.; Almeida, M.M.; Dos Santos, J.P.; Boia, M.N.; Coronato-Nunes, B.; Jaeger, L.H.; Carvalho-Costa, F.A. Epidemiology, Species Composition and Genetic Diversity of Tetra- and Octonucleated Entamoeba spp. in Different Brazilian Biomes. Parasites Vectors 2021, 14, 160. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Non-Human Primates | Samples |
|---|---|
| Callitrichidae | |
| Callithrix sp. (Erxleben, 1777) | 19 |
| Callithrix aurita (Geoffroy, 1812) | 16 |
| Callithrix geoffroyi (Humboldt, 1812) | 1 |
| Callithrix jacchus (Linnaeus, 1758) | 7 |
| Callithrix penicillata (Geoffroy, 1812) | 19 |
| Cebuella pygmaea (Spix, 1823) | 2 |
| Mico argentatus (Linnaeus, 1771) | 1 |
| Mico chrysoleucus (Wagner, 1842) | 1 |
| Mico humeralifer (Geoffroy, 1812) | 1 |
| Mico mauesi (Mittermeier, Schwarz & Ayres, 1992) | 1 |
| Mico melanurus (Geoffroy, 1812) | 1 |
| Saguinus bicolor (Spix, 1823) | 11 |
| Saguinus fuscicollis (Spix, 1823) | 1 |
| Saguinus martinsi (Thomas, 1912) | 2 |
| Saguinus midas (Linnaeus, 1758) | 5 |
| Saguinus niger (Geoffroy, 1803) | 1 |
| Saguinus ursulus (Hoffmannsegg, 1807) | 8 |
| Leontocebus weddelli (Deville, 1849) | 11 |
| Leontopithecus sp. (Lesson 1820) | 1 |
| Leontopithecus chrysomelas (Kuhl, 1820) | 99 |
| Leontopithecus chrysopygus (Mikan, 1823) | 4 |
| Leontopithecus rosalia (Linnaeus, 1766) | 9 |
| Callimico goeldii (Thomas, 1904) | 4 |
| Aotidae | |
| Aotus sp. (Illiger, 1811) | 2 |
| Aotus infulatus (Kuhl, 1820) | 48 |
| Aotus nigriceps (Dollman, 1909) | 1 |
| Aotus trivirgatus (Humboldt, 1811) | 3 |
| Cebidae | |
| Cebus albifrons (Humboldt, 1812) | 3 |
| Cebus kaapori (Queiroz, 1992) | 1 |
| Cebus olivaceus (Schomburgk, 1848) | 5 |
| Sapajus sp. (Kerr, 1792) | 2 |
| Sapajus apella (Linnaeus, 1758) | 22 |
| Sapajus libidinosus (Spix, 1823) | 15 |
| Sapajus nigritus (Goldfuss, 1809) | 2 |
| Sapajus xanthosternos (Wied-Neuwied, 1826) | 12 |
| Sapajus robustus (Kuhl, 1820) | 3 |
| Saimiri boliviensis (Geoffroy & Blainville, 1834) | 1 |
| Saimiri collinsi (Osgood, 1916) | 30 |
| Pitheciidae | |
| Callicebus melanochir (Wied-Neuwied, 1820) | 1 |
| Plecturocebus caquetensis (Defler, Bueno & García, 2010) | 1 |
| Plecturocebus hoffmannsi (Thomas, 1908) and Cheracebus purinus (Thomas, 1927) | 2 |
| Plecturocebus dubius (Hershkovitz, 1988) | 1 |
| Plecturocebus vieirai (Gualda-Barros, Nascimento & Amaral, 2012) | 4 |
| Pithecia mittermeieri (Marsh, 2014) | 1 |
| Pithecia monachus (Geoffroy, 1812) | 1 |
| Chiropotes satanas (Hoffmannsegg, 1807) | 1 |
| Chiropotes utahickae (Hershkovitz, 1985) | 1 |
| Cacajao melanocephalus (Humboldt, 1812) | 1 |
| Atelidae | |
| Alouatta sp. (Lacepede, 1799) | 20 |
| Alouatta caraya (Humboldt, 1812) | 28 |
| Alouatta discolor (Spix, 1823) | 1 |
| Alouatta guariba (Humboldt, 1812) | 33 |
| Ateles chamek (Humboldt, 1812) | 3 |
| Ateles paniscus (Linnaeus, 1758) | 2 |
| Ateles marginatus (Geoffroy, 1809) | 7 |
| Brachyteles arachnoides (Geoffroy, 1806) | 1 |
| Cercopithecidae | |
| Mandrillus sphinx (Linnaeus, 1758) | 1 |
| Papio hamadryas (Linnaeus, 1758) | 4 |
| Chlorocebus aethiops (Linnaeus, 1758) | 11 |
| Hominidae | |
| Pan troglodytes (Linnaeus, 1758) | 8 |
| Lemuridae | |
| Lemur catta (Linnaeus, 1758) | 3 |
| Host | Positive Samples for Amoebozoa Subjected to Isolation | Positive Samples for Ciliophora Subjected to Isolation | Molecular Identification | NI ** | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Feces | In Vitro Cultivation | Feces | In Vitro Cultivation | |||||||||
| E. dispar | E. histolytica | E. hartmanni | E. coli | Iodamoeba sp. | E. nana | Iodamoeba sp. | B. coli | B. coli | ||||
| Callitrichidae | ||||||||||||
| Callithrix penicillata | 2 | - | - | - | - | - | 1 | - | - | - | - | 1 |
| Callithrix jacchus | 1 | - | - | - | - | - | 1 | - | - | - | - | - |
| Callithrix humeralifer and Leontopithecus chrysomelas | 1 | - | - | - | - | - | - | - | - | - | - | 1 |
| Hybrid C. jacchus × C. penicillata | - | 1 | - | - | - | - | - | - | - | 1 | - | - |
| Leontopithecus chrysomelas | 1 | - | - | 1 | - | - | - | - | - | - | - | - |
| Saguinus bicolor | 2 | - | - | 1 | - | - | - | - | - | - | - | 1 |
| Saguinus ursulus | 1 | - | - | - | - | - | - | - | - | - | - | 1 |
| Saguinus midas and Callicebus sp.* | 1 | 1 | - | - | - | 1 | - | 1 | - | 1 | - | - |
| Leontocebus weddelli | 1 | - | - | - | - | - | - | - | - | - | - | 1 |
| Callimico goeldii | 1 | - | - | - | - | - | - | - | - | - | - | 1 |
| Aotidae | ||||||||||||
| Aotus trivirgatus | 1 | - | - | - | - | - | - | - | - | - | - | 1 |
| Aotus infulatus * | 12 | - | 8 | - | 5 | 3 | 5 | 1 | - | - | - | 2 |
| Cebidae | ||||||||||||
| Saimiri collinsi | 3 | - | - | - | 1 | - | - | - | - | - | - | 2 |
| Cebus olivaceus | 1 | - | - | - | - | - | - | - | - | - | - | 1 |
| Sapajus libidinosus * | 1 | 2 | - | - | - | 1 | 1 | - | - | 2 | - | - |
| Sapajus xanthosternos | 1 | - | - | - | - | - | - | - | 1 | - | - | - |
| Sapajus apella, Sapajus libidinosus and Cebus albifrons | 1 | - | - | 1 | - | - | - | - | - | - | - | - |
| Pitheciidae | - | |||||||||||
| Pithecia mittermeieri | 1 | - | - | - | - | 1 | - | - | - | - | - | - |
| Cheracebus purinus and Plecturocebus hoffmannsi | 1 | - | - | - | - | - | - | - | - | - | - | 1 |
| Atelidae | ||||||||||||
| Alouatta caraya * | 7 | - | 4 | - | 1 | 2 | 1 | 2 | - | - | - | 1 |
| Alouatta guariba * | 8 | 1 | 4 | 1 | - | 1 | - | - | - | 1 | - | 1 |
| Cercopithecidae | ||||||||||||
| Chlorocebus aethiops * | 7 | - | 3 | - | 6 | 2 | - | 3 | - | - | - | - |
| Hominidae | ||||||||||||
| Pan troglodytes * | 6 | 1 | 6 | - | 6 | - | - | - | - | - | 1 | - |
| Homo sapiens | 12 | - | 2 | - | 2 | 1 | - | - | - | - | - | 7 |
| Total | 73 | 6 | 27 | 4 | 21 | 12 | 9 | 7 | 1 | 5 | 1 | 22 |
| Sequences | This Study | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| E. dispar (A. caraya) | E. dispar handler | E. hartmanni (C. aethiops) | E. hartmanni handler | E. coli (A. infulatus) | E. coli handler | E. histolytica (A. guariba) | B. coli A0 (P. troglodytes) | B. coli B0 (C. jacchus × C. penicillata) | |
| E. dispar handler (this study) | 100% | - | - | - | - | - | - | - | - |
| E. dispar human (KP722600) | 100% | 100% | - | - | - | - | - | - | - |
| E. dispar—Macaca mulatta (Zimmermann, 1780) (AB282661) | 100% | 100% | - | - | - | - | - | - | - |
| E. hartmanni handler (this study) | - | - | 97.66% | - | - | - | - | - | - |
| E. hartmanni human (MG925066) | - | - | 97.71% | 100% | - | - | - | - | - |
| E. hartmanni—Macaca fascicularis (PP490721) | - | - | 96.65% | 100% | - | - | - | - | - |
| E. coli handler (this study) | - | - | - | - | 86.38% | - | - | - | - |
| E. coli human (MG925062) | - | - | - | - | 86.38% | 99.63% | - | - | - |
| E. coli—Gorilla gorilla (Savage, 1847) (AB444953) | - | - | - | - | 98.02% | 85.29% | - | - | - |
| E. histolytica human (KP233840) | - | - | - | - | - | - | 100% | - | - |
| E. histolytica—Macaca fascicularis (GQ423749) | - | - | - | - | - | - | 100% | - | - |
| B. coli variant A0—human (JF444758) | - | - | - | - | - | - | - | 99.74% | 92.73% |
| B coli variant A1—Gorilla gorilla (JF444760) | - | - | - | - | - | - | - | 98.18% | 93.07% |
| B coli variant B1—Gorilla gorilla (JF444761) | - | - | - | - | - | - | - | 91.88% | 99.08% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dib, L.; da Silva, B.; Correa, L.; Pissinatti, A.; Moreira, S.; Tavares, M.C.; Teixeira, R.; da Costa, A.L.; Muniz, J.A.; Junglos, A.; et al. Diversity of the Amoebozoa and Ciliophora Groups in Non-Human Primates Kept Ex Situ and in Their Handlers in Different Institutions in Brazil. Pathogens 2025, 14, 56. https://doi.org/10.3390/pathogens14010056
Dib L, da Silva B, Correa L, Pissinatti A, Moreira S, Tavares MC, Teixeira R, da Costa AL, Muniz JA, Junglos A, et al. Diversity of the Amoebozoa and Ciliophora Groups in Non-Human Primates Kept Ex Situ and in Their Handlers in Different Institutions in Brazil. Pathogens. 2025; 14(1):56. https://doi.org/10.3390/pathogens14010056
Chicago/Turabian StyleDib, Laís, Breno da Silva, Lais Correa, Alcides Pissinatti, Silvia Moreira, Maria Clotilde Tavares, Rodrigo Teixeira, André Luíz da Costa, José Augusto Muniz, Amauri Junglos, and et al. 2025. "Diversity of the Amoebozoa and Ciliophora Groups in Non-Human Primates Kept Ex Situ and in Their Handlers in Different Institutions in Brazil" Pathogens 14, no. 1: 56. https://doi.org/10.3390/pathogens14010056
APA StyleDib, L., da Silva, B., Correa, L., Pissinatti, A., Moreira, S., Tavares, M. C., Teixeira, R., da Costa, A. L., Muniz, J. A., Junglos, A., Hirano, Z. M., Dada, A., da Silva, S., Amendoeira, M. R., & Barbosa, A. (2025). Diversity of the Amoebozoa and Ciliophora Groups in Non-Human Primates Kept Ex Situ and in Their Handlers in Different Institutions in Brazil. Pathogens, 14(1), 56. https://doi.org/10.3390/pathogens14010056