

Suppression by RNA Polymerase I Inhibitors Varies Greatly Between Distinct RNA Polymerase I Transcribed Genes in Malaria Parasites


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Parasite Culture
2.2. P. falciparum Growth Inhibition Assay
2.3. RNA Polymerase I Inhibition Assay
2.4. RNA Isolation, cDNA Synthesis, and qPCR
3. Results and Discussion
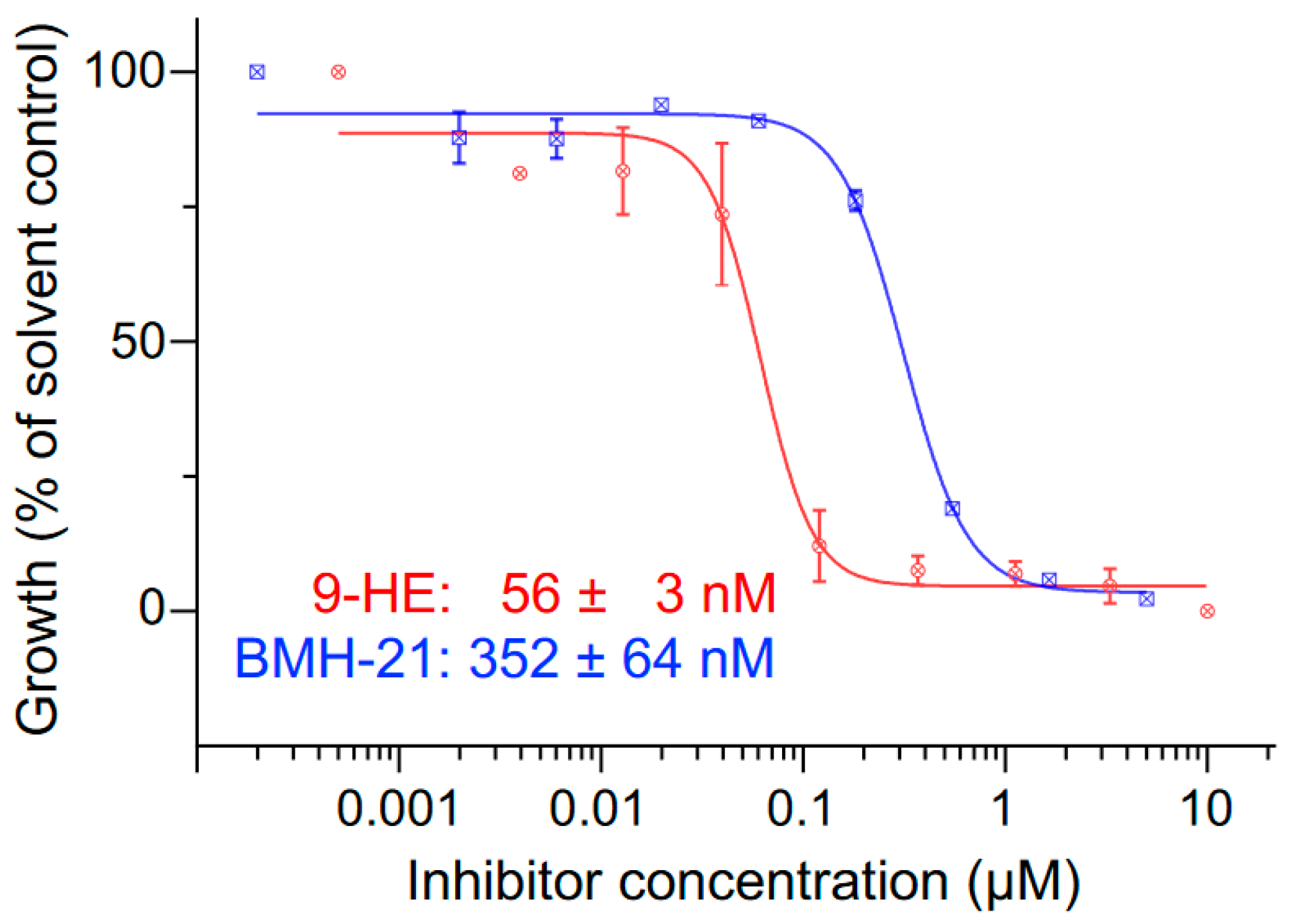
3.1. Inhibition of P. falciparum RNA Polymerase I Activity by 9-HE and BMH-21
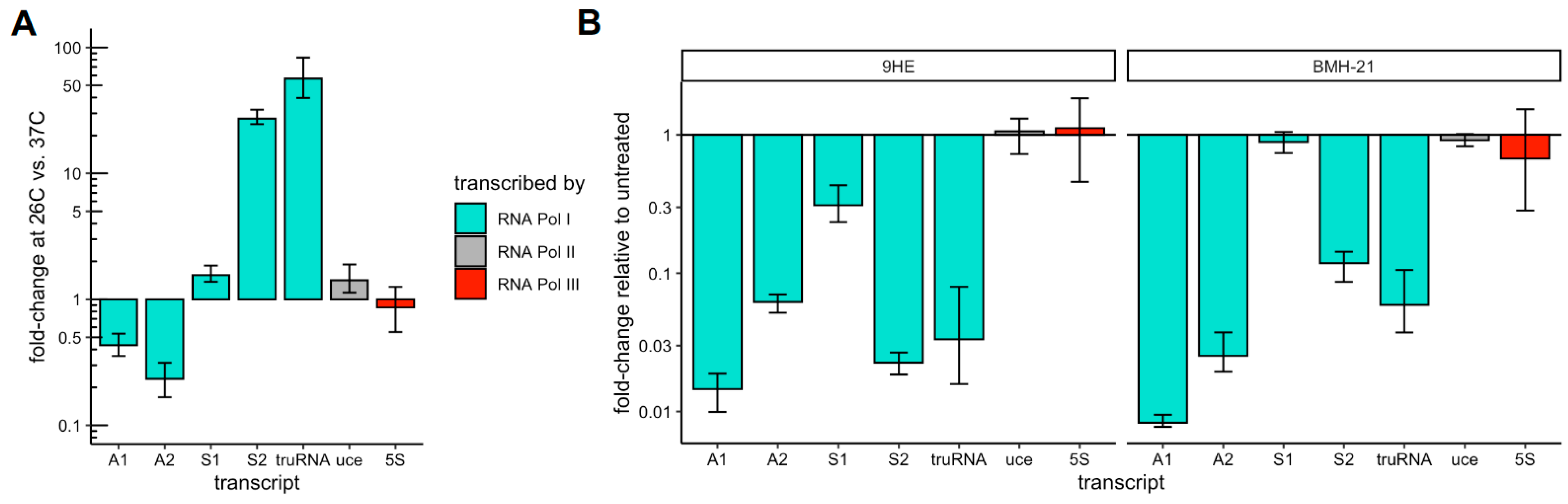
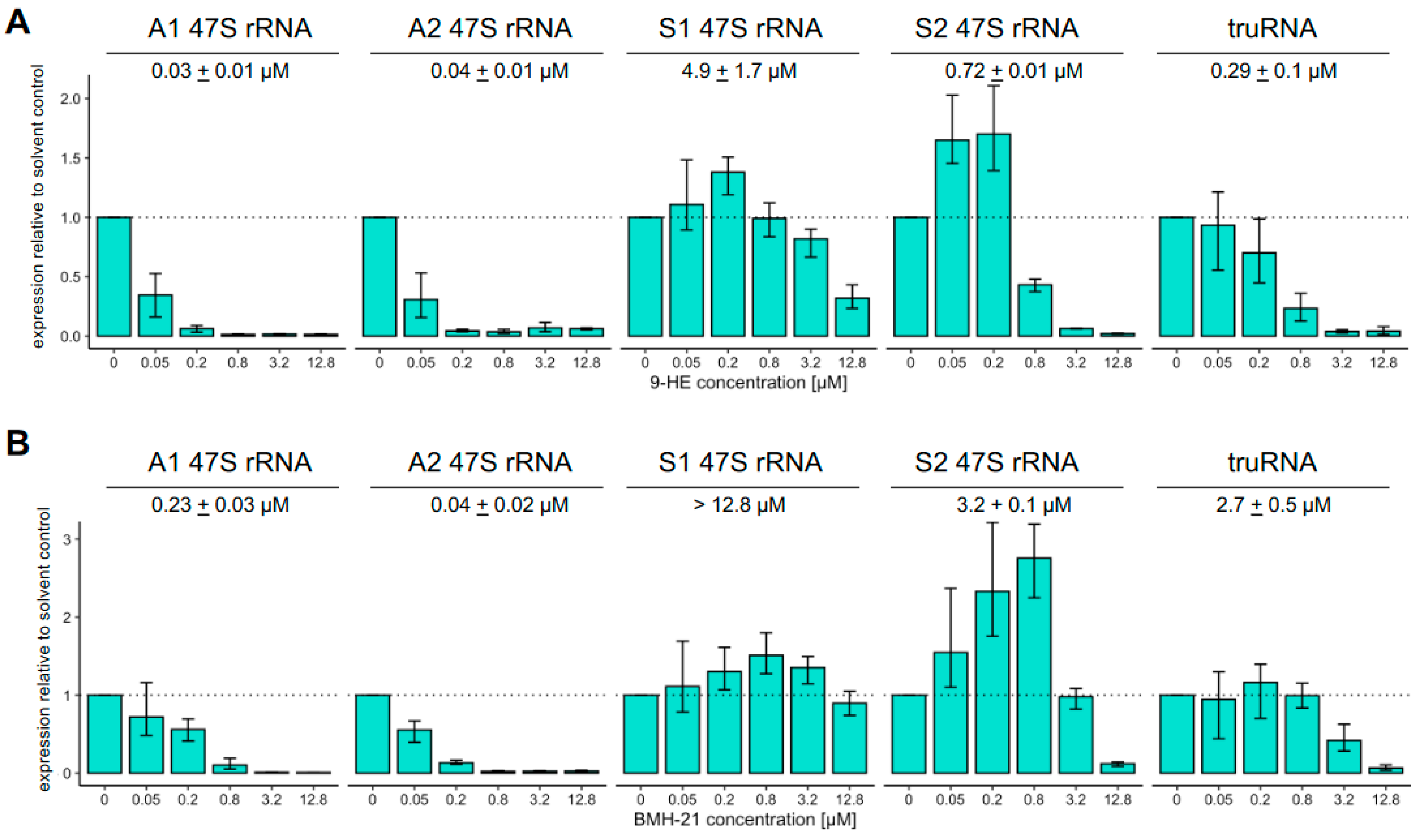
3.2. Susceptibility to Inhibitors of RNA Pol I Activity Differs Greatly Between Pol I Transcribed Genes
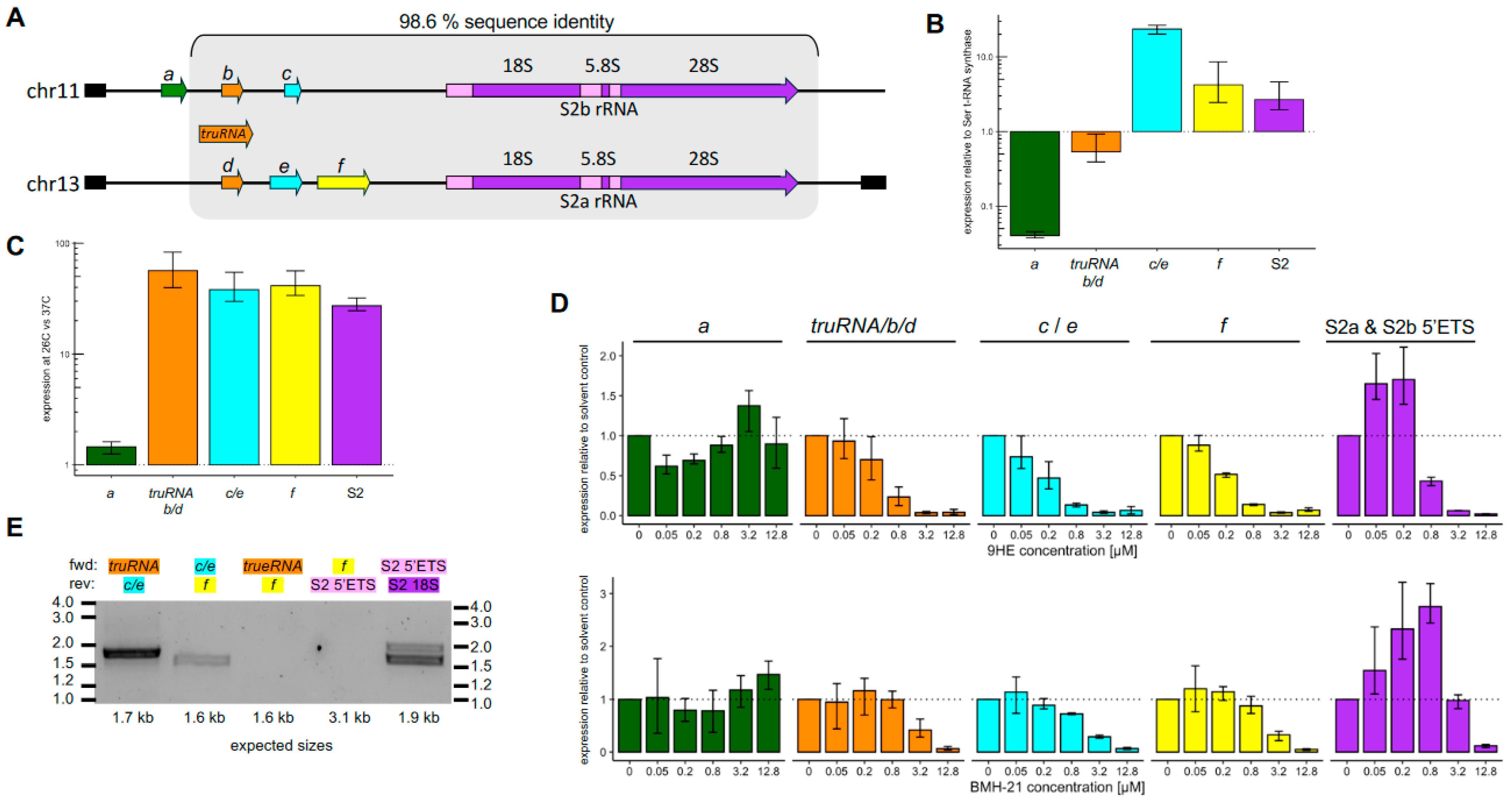
3.3. lncRNAs Downstream of the truRNA Likely Derive from a Single Longer Transcript
3.4. P. falciparum Lacks Identifiable Homologs to Many RNA Pol I-Specific Components
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Petrov, A.S.; Gulen, B.; Norris, A.M.; Kovacs, N.A.; Bernier, C.R.; Lanier, K.A.; Fox, G.E.; Harvey, S.C.; Wartell, R.M.; Hud, N.V.; et al. History of the Ribosome and the Origin of Translation. Proc. Natl. Acad. Sci. USA 2015, 112, 15396–15401. [Google Scholar] [CrossRef] [PubMed]
- Fox, G.E. Origin and Evolution of the Ribosome. Cold Spring Harb. Perspect. Biol. 2010, 2, a003483. [Google Scholar] [CrossRef] [PubMed]
- Cooper, G.M. Translation of MRNA; Sinauer Associates: Sunderland, MA, USA, 2000. [Google Scholar]
- Yang, Y.-M.; Karbstein, K. Ribosome Assembly and Repair. Annu. Rev. Cell Dev. Biol. 2024, 40, 241–264. [Google Scholar] [CrossRef] [PubMed]
- Goodfellow, S.J.; Zomerdijk, J.C.B.M. Basic Mechanisms in RNA Polymerase I Transcription of the Ribosomal RNA Genes. Subcell. Biochem. 2013, 61, 211–236. [Google Scholar] [PubMed]
- Sáez-Vásquez, J.; Delseny, M. Ribosome Biogenesis in Plants: From Functional 45S Ribosomal DNA Organization to Ribosome Assembly Factors. Plant Cell 2019, 31, 1945–1967. [Google Scholar] [CrossRef]
- Nomura, M.; Nogi, Y.; Oakes, M. Transcription of RDNA in the Yeast Saccharomyces Cerevisiae. In Madame Curie Bioscience Database [Internet]; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- McGee, J.P.; Armache, J.-P.; Lindner, S.E. Ribosome Heterogeneity and Specialization of Plasmodium Parasites. PLoS Pathog. 2023, 19, e1011267. [Google Scholar] [CrossRef]
- Rodríguez-Almonacid, C.C.; Kellogg, M.K.; Karamyshev, A.L.; Karamysheva, Z.N. Ribosome Specialization in Protozoa Parasites. Int. J. Mol. Sci. 2023, 24, 7484. [Google Scholar] [CrossRef]
- Waters, A.P.; van Spaendonk, R.M.; Ramesar, J.; Vervenne, R.A.; Dirks, R.W.; Thompson, J.; Janse, C.J. Species-Specific Regulation and Switching of Transcription between Stage-Specific Ribosomal RNA Genes in Plasmodium Berghei. J. Biol. Chem. 1997, 272, 3583–3589. [Google Scholar] [CrossRef]
- Waters, A.P.; Syin, C.; McCutchan, T.F. Developmental Regulation of Stage-Specific Ribosome Populations in Plasmodium. Nature 1989, 342, 438–440. [Google Scholar] [CrossRef]
- Rogers, M.J.; Gutell, R.R.; Damberger, S.H.; Li, J.; McConkey, G.A.; Waters, A.P.; McCutchan, T.F. Structural Features of the Large Subunit RRNA Expressed in Plasmodium Falciparum Sporozoites That Distinguish It from the Asexually Expressed Subunit RRNA. RNA 1996, 2, 134–145. [Google Scholar]
- Zhu, J.D.; Waters, A.P.; Appiah, A.; McCutchan, T.F.; Lal, A.A.; Hollingdale, M.R. Stage-Specific Ribosomal RNA Expression Switches during Sporozoite Invasion of Hepatocytes. J. Biol. Chem. 1990, 265, 12740–12744. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; McCutchan, T.F. Thermoregulation in a Parasite’s Life Cycle. Nature 2002, 418, 742. [Google Scholar] [CrossRef]
- Fang, J.; Sullivan, M.; McCutchan, T.F. The Effects of Glucose Concentration on the Reciprocal Regulation of RRNA Promoters in Plasmodium Falciparum. J. Biol. Chem. 2004, 279, 720–725. [Google Scholar] [CrossRef]
- Sharma, I.; Fang, J.; Lewallen, E.A.; Deitsch, K.W.; McCutchan, T.F. Identification of a Long Noncoding RNA Required for Temperature Induced Expression of Stage-Specific RRNA in Malaria Parasites. Gene 2023, 877, 147516. [Google Scholar] [CrossRef]
- Cramer, P.; Armache, K.-J.; Baumli, S.; Benkert, S.; Brueckner, F.; Buchen, C.; Damsma, G.E.; Dengl, S.; Geiger, S.R.; Jasiak, A.J.; et al. Structure of Eukaryotic RNA Polymerases. Annu. Rev. Biophys. 2008, 37, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Warner, J.R.; Vilardell, J.; Sohn, J.H. Economics of Ribosome Biosynthesis. Cold Spring Harb. Symp. Quant. Biol. 2001, 66, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Moss, T.; Stefanovsky, V.Y. Promotion and Regulation of Ribosomal Transcription in Eukaryotes by RNA Polymerase I. Prog. Nucleic Acid Res. Mol. Biol. 1995, 50, 25–66. [Google Scholar]
- Teng, T.; Thomas, G.; Mercer, C.A. Growth Control and Ribosomopathies. Curr. Opin. Genet. Dev. 2013, 23, 63–71. [Google Scholar] [CrossRef]
- Hwang, S.-P.; Denicourt, C. The Impact of Ribosome Biogenesis in Cancer: From Proliferation to Metastasis. NAR Cancer 2024, 6, zcae017. [Google Scholar] [CrossRef]
- Kang, J.; Brajanovski, N.; Chan, K.T.; Xuan, J.; Pearson, R.B.; Sanij, E. Ribosomal Proteins and Human Diseases: Molecular Mechanisms and Targeted Therapy. Signal Transduct. Target. Ther. 2021, 6, 323. [Google Scholar] [CrossRef]
- Bensaude, O. Inhibiting Eukaryotic Transcription: Which Compound to Choose? How to Evaluate Its Activity? Transcription 2011, 2, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Laham-Karam, N.; Pinto, G.P.; Poso, A.; Kokkonen, P. Transcription and Translation Inhibitors in Cancer Treatment. Front. Chem. 2020, 8, 276. [Google Scholar] [CrossRef] [PubMed]
- Günzl, A.; Bruderer, T.; Laufer, G.; Schimanski, B.; Tu, L.-C.; Chung, H.-M.; Lee, P.-T.; Lee, M.G.-S. RNA Polymerase I Transcribes Procyclin Genes and Variant Surface Glycoprotein Gene Expression Sites in Trypanosoma Brucei. Eukaryot. Cell 2003, 2, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Moll, K.; Ljungström, I.; Perlmann, H.; Scherf, A. Methods in Malaria Research, 6th ed.; MR4/ATCC: Manassas, VA, USA, 2013. [Google Scholar]
- Johnson, J.D.; Dennull, R.A.; Gerena, L.; Lopez-Sanchez, M.; Roncal, N.E.; Waters, N.C. Assessment and Continued Validation of the Malaria SYBR Green I-Based Fluorescence Assay for Use in Malaria Drug Screening. Antimicrob. Agents Chemother. 2007, 51, 1926–1933. [Google Scholar] [CrossRef]
- Smilkstein, M.; Sriwilaijaroen, N.; Kelly, J.X.; Wilairat, P.; Riscoe, M. Simple and Inexpensive Fluorescence-Based Technique for High-Throughput Antimalarial Drug Screening. Antimicrob. Agents Chemother. 2004, 48, 1803–1806. [Google Scholar] [CrossRef] [PubMed]
- Bancells, C.; Llorà-Batlle, O.; Poran, A.; Nötzel, C.; Rovira-Graells, N.; Elemento, O.; Kafsack, B.F.C.; Cortés, A. Revisiting the Initial Steps of Sexual Development in the Malaria Parasite Plasmodium Falciparum. Nat. Microbiol. 2019, 4, 144–154. [Google Scholar] [CrossRef]
- Salanti, A.; Staalsoe, T.; Lavstsen, T.; Jensen, A.T.R.; Sowa, M.P.K.; Arnot, D.E.; Hviid, L.; Theander, T.G. Selective Upregulation of a Single Distinctly Structured Var Gene in Chondroitin Sulphate A-Adhering Plasmodium Falciparum Involved in Pregnancy-Associated Malaria. Mol. Microbiol. 2003, 49, 179–191. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New Capabilities and Interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Jacobs, R.Q.; Huffines, A.K.; Laiho, M.; Schneider, D.A. The Small-Molecule BMH-21 Directly Inhibits Transcription Elongation and DNA Occupancy of RNA Polymerase I In Vivo and In Vitro. J. Biol. Chem. 2022, 298, 101450. [Google Scholar] [CrossRef]
- Andrews, W.J.; Panova, T.; Normand, C.; Gadal, O.; Tikhonova, I.G.; Panov, K.I. Old Drug, New Target: Ellipticines Selectively Inhibit Rna Polymerase I Transcription. J. Biol. Chem. 2013, 288, 4567–4582. [Google Scholar] [CrossRef]
- Kerry, L.E.; Pegg, E.E.; Cameron, D.P.; Budzak, J.; Poortinga, G.; Hannan, K.M.; Hannan, R.D.; Rudenko, G. Selective Inhibition of RNA Polymerase I Transcription as a Potential Approach to Treat African Trypanosomiasis. PLoS Negl. Trop. Dis. 2017, 11, e0005432. [Google Scholar] [CrossRef] [PubMed]
- Montoia, A.; Rocha E Silva, L.F.; Torres, Z.E.; Costa, D.S.; Henrique, M.C.; Lima, E.S.; Vasconcellos, M.C.; Souza, R.C.Z.; Costa, M.R.F.; Grafov, A.; et al. Antiplasmodial Activity of Synthetic Ellipticine Derivatives and an Isolated Analog. Bioorg. Med. Chem. Lett. 2014, 24, 2631–2634. [Google Scholar] [CrossRef] [PubMed]
- Bénard, J.; Dat-Xuong, N.; Riou, G. Trypanocidal activity of some ellipticine derivatives against Trypanosoma cruzi cultured in vitro. C.R. Hebd. Seances Acad. Sci. Ser. D Sci. Nat. 1975, 280, 1177–1180. [Google Scholar]
- Hughes, J.M.; Ares, M., Jr. Depletion of U3 Small Nucleolar RNA Inhibits Cleavage in the 5′ External Transcribed Spacer of Yeast Pre-Ribosomal RNA and Impairs Formation of 18S Ribosomal RNA. EMBO J. 1991, 10, 4231–4239. [Google Scholar] [CrossRef] [PubMed]
- Aubert, M.; O’Donohue, M.-F.; Lebaron, S.; Gleizes, P.-E. Pre-Ribosomal RNA Processing in Human Cells: From Mechanisms to Congenital Diseases. Biomolecules 2018, 8, 123. [Google Scholar] [CrossRef]
- Elcock, A.H.; Rodger, A.; Richards, W.G. Theoretical Studies of the Intercalation of 9-Hydroxyellipticine in DNA. Biopolymers 1996, 39, 309–326. [Google Scholar] [CrossRef]
- Albert, B.; Perez-Fernandez, J.; Léger-Silvestre, I.; Gadal, O. Regulation of Ribosomal RNA Production by RNA Polymerase I: Does Elongation Come First? Genet. Res. Int. 2012, 2012, 276948. [Google Scholar] [CrossRef]
- Nguyen, T.N.; Schimanski, B.; Günzl, A. Active RNA Polymerase I of Trypanosoma Brucei Harbors a Novel Subunit Essential for Transcription. Mol. Cell. Biol. 2007, 27, 6254–6263. [Google Scholar] [CrossRef]
- Takebe, S.; Witola, W.H.; Schimanski, B.; Günzl, A.; Ben Mamoun, C. Purification of Components of the Translation Elongation Factor Complex of Plasmodium Falciparum by Tandem Affinity Purification. Eukaryot. Cell 2007, 6, 584–591. [Google Scholar] [CrossRef]
- Schimanski, B.; Nguyen, T.N.; Günzl, A. Highly Efficient Tandem Affinity Purification of Trypanosome Protein Complexes Based on a Novel Epitope Combination. Eukaryot. Cell 2005, 4, 1942–1950. [Google Scholar] [CrossRef]
- Brandenburg, J.; Schimanski, B.; Nogoceke, E.; Nguyen, T.N.; Padovan, J.C.; Chait, B.T.; Cross, G.A.M.; Günzl, A. Multifunctional Class I Transcription in Trypanosoma Brucei Depends on a Novel Protein Complex. EMBO J. 2007, 26, 4856–4866. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samuel, H.; Campelo Morillo, R.; Kafsack, B.F.C. Suppression by RNA Polymerase I Inhibitors Varies Greatly Between Distinct RNA Polymerase I Transcribed Genes in Malaria Parasites. Pathogens 2024, 13, 924. https://doi.org/10.3390/pathogens13110924
Samuel H, Campelo Morillo R, Kafsack BFC. Suppression by RNA Polymerase I Inhibitors Varies Greatly Between Distinct RNA Polymerase I Transcribed Genes in Malaria Parasites. Pathogens. 2024; 13(11):924. https://doi.org/10.3390/pathogens13110924
Chicago/Turabian StyleSamuel, Hermela, Riward Campelo Morillo, and Björn F. C. Kafsack. 2024. "Suppression by RNA Polymerase I Inhibitors Varies Greatly Between Distinct RNA Polymerase I Transcribed Genes in Malaria Parasites" Pathogens 13, no. 11: 924. https://doi.org/10.3390/pathogens13110924
APA StyleSamuel, H., Campelo Morillo, R., & Kafsack, B. F. C. (2024). Suppression by RNA Polymerase I Inhibitors Varies Greatly Between Distinct RNA Polymerase I Transcribed Genes in Malaria Parasites. Pathogens, 13(11), 924. https://doi.org/10.3390/pathogens13110924