

Comparative Pathology of Animal Models for Influenza A Virus Infection




Abstract
:1. Introduction
2. Human Disease and Clinical Symptoms caused by IAV and IBV
3. Influenza Pathology

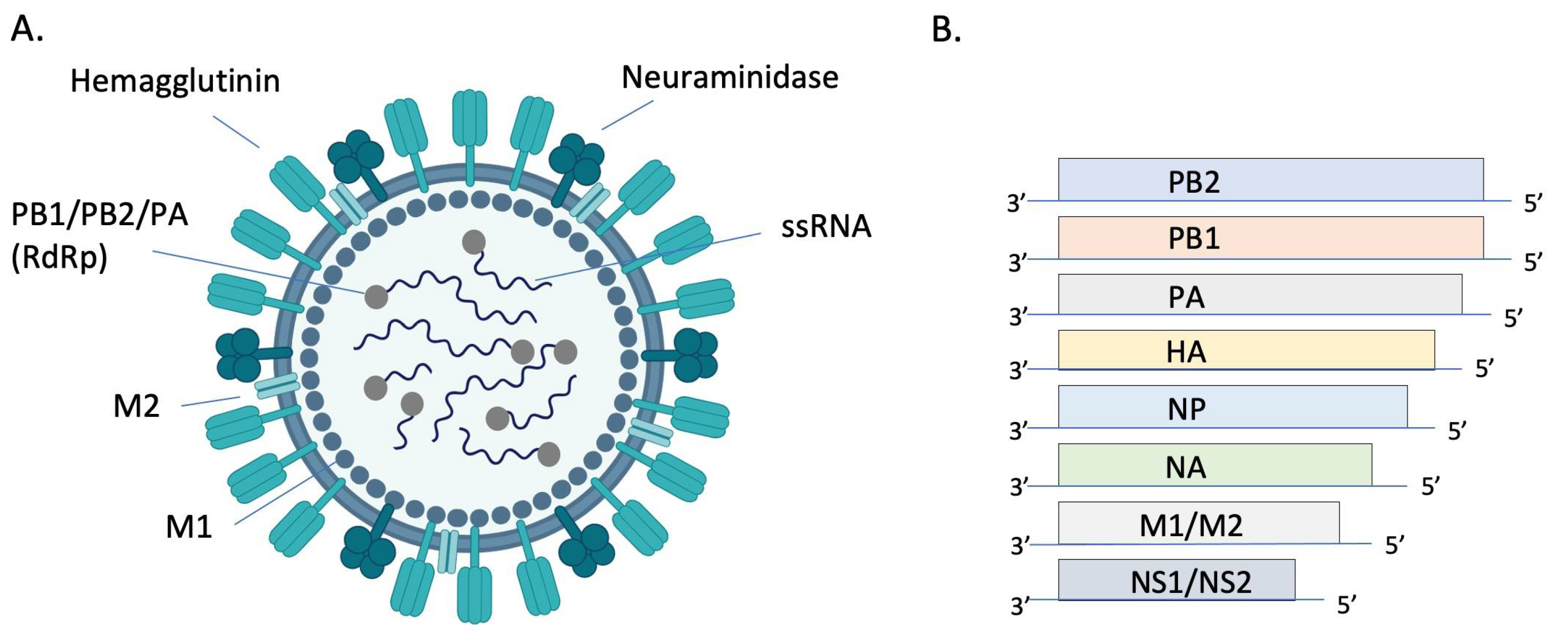
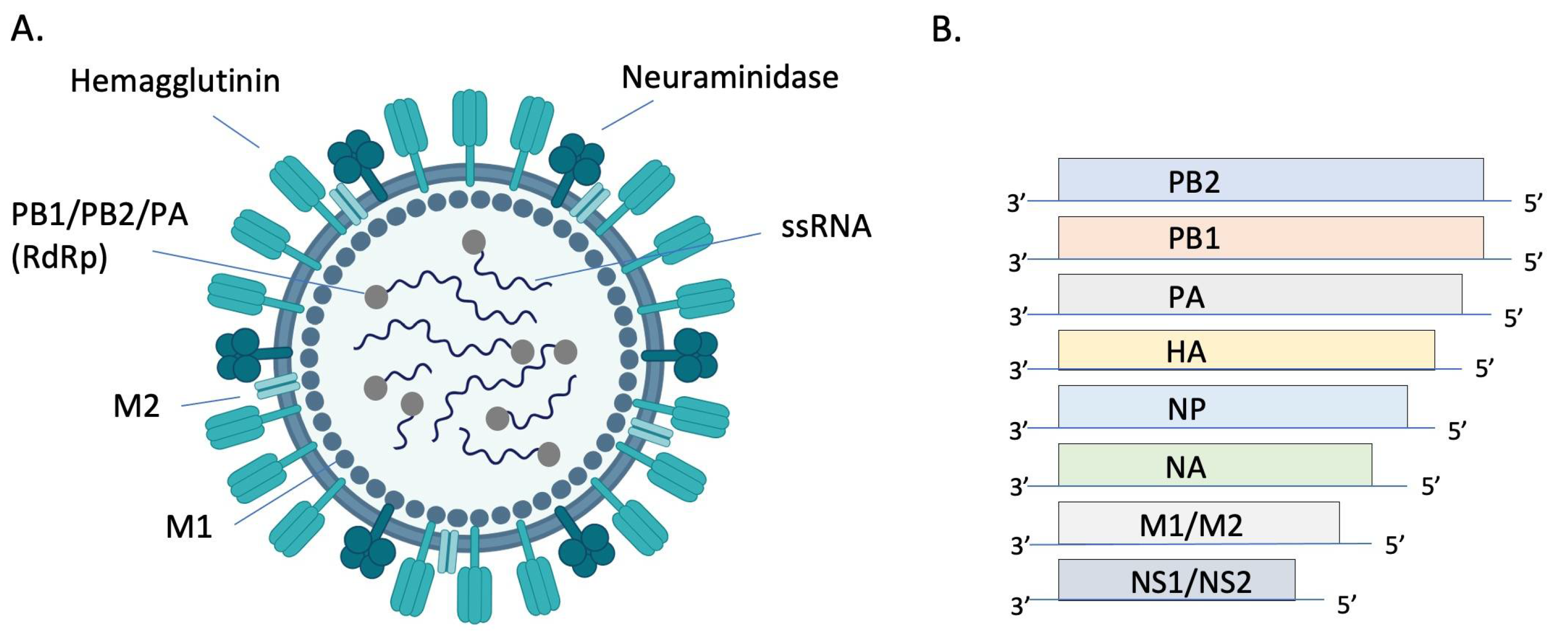{kind=link}
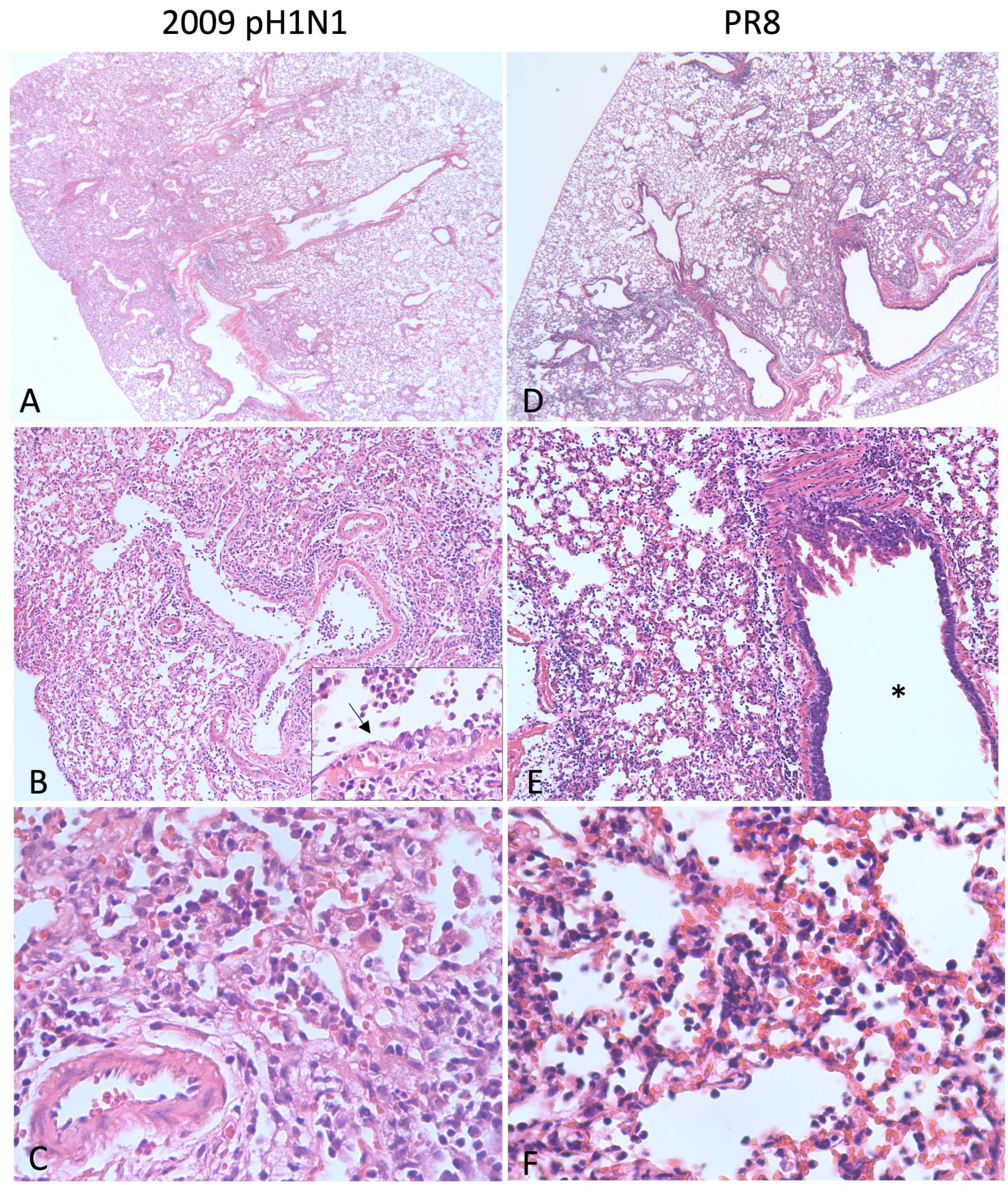
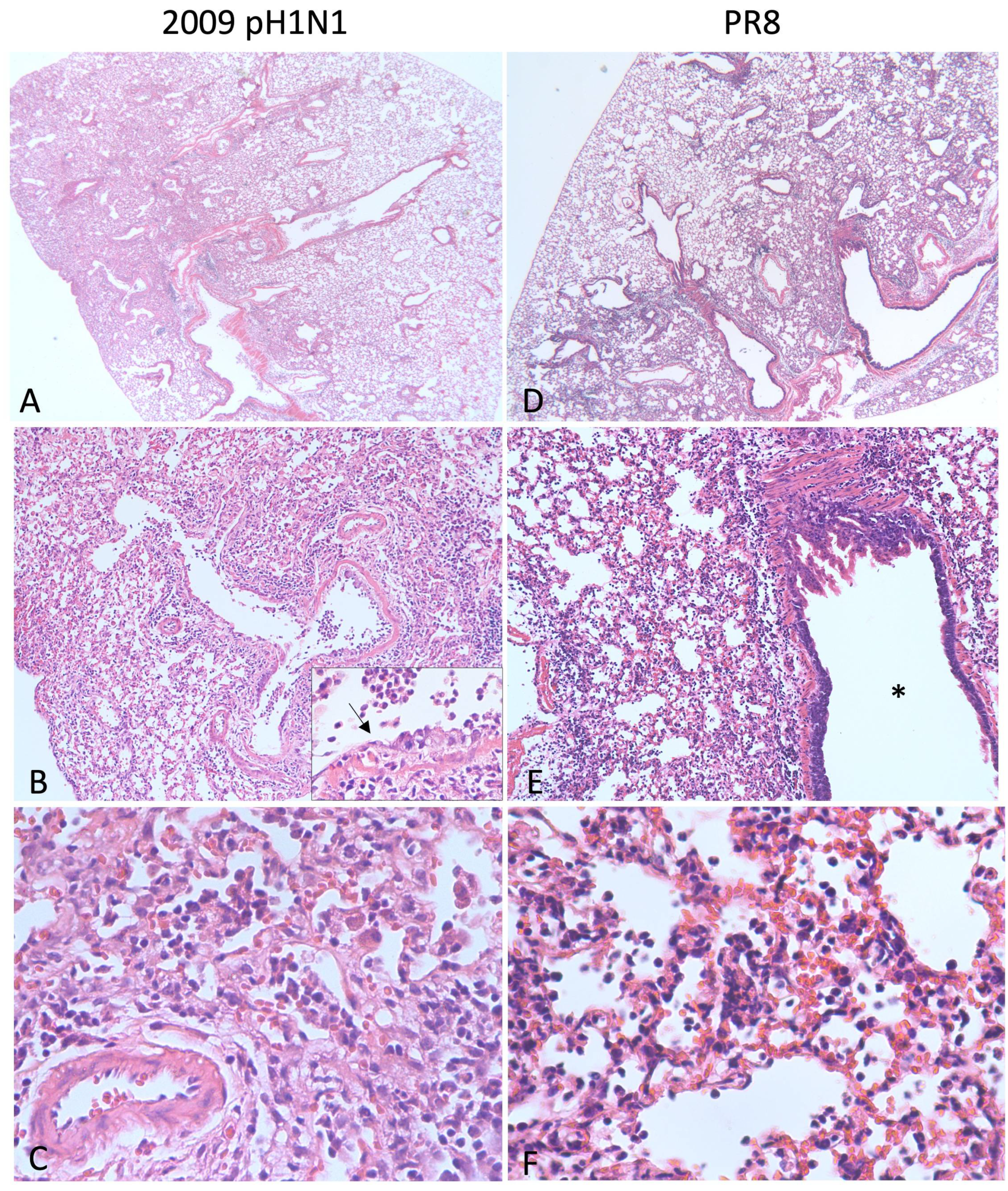{kind=link}
| Common Animal Strain/Species | Virus Strain | Clinical Signs | Microscopic Pathology | References | |
|---|---|---|---|---|---|
| Humans | N/A | Seasonal IAV | Varying degrees of fever, non-productive cough, dyspnea, coryza, fatigue, and myalgia (classic flu symptoms); vomiting and diarrhea in severe cases | Necrotizing tracheobronchitis and bronchointerstitial pneumonia with thrombi, edema, hemorrhage, hyaline membranes, and type II pneumocyte hyperplasia (diffuse alveolar damage) | [25,26,50] |
| 2009 pH1N1 | Mild to severe flu symptoms | Same as seasonal IAV | [30,34,50,52] | ||
| 1918 H1N1 | Mild to severe flu symptoms | Same as seasonal IAV | [17,25,53] | ||
| HPAI H5N1 and H7N9 | Mild to severe flu symptoms with history of contact with live poultry | Same as seasonal IAV Extrapulmonary necrotic lesions are common and hemophagocytosis may be the most prominent lesion | [19,35,36,37,50,51,70,71] | ||
| Mouse | BALB/c C57BL/6 DBA/2J A/J | PR8 | Dyspnea, ruffled fur, weight loss, and anorexia | Interstitial pneumonia, suppurative bronchiolitis and alveolitis, hyaline membranes, and alveolar edema | [72,73,74,75] |
| BALB/c C57BL/6 | 2009 pH1N1 | Variable weight loss (dose and strain dependent) | Histiocytic to neutrophilic bronchitis, bronchiolitis and alveolitis with varying epithelial necrosis (strain dependent) | [76,77,78,79] | |
| BALB/c | 1918 H1N1 | Weight loss and death | Interstitial pneumonia, suppurative bronchiolitis and alveolitis, hyaline membranes, and alveolar edema | [76,80,81] | |
| BALB/c DBA/2J | HPAI H5N1 | Dyspnea, ruffled fur, weight loss, and anorexia | Interstitial pneumonia, suppurative bronchiolitis and alveolitis, hyaline membranes, and alveolar edema Encephalitis and myocardial necrosis | [81,82,83,84,85,86,87,88] | |
| C57BL/6 BALB/c | H7N9 | Weight loss, ruffled fur, hunching | Bronchiolitis, patchy interstitial pneumonia, and varying amounts of bronchiolar and alveolar epithelial necrosis | [89,90,91,92] | |
| BALB/c | LPAI | Variable weight loss, ruffled fur, and hunching (strain dependent) | Necrotizing bronchitis and bronchiolitis with peribronchial pneumonia | [93,94,95,96,97] | |
| Hamster | Golden Syrian hamster | Seasonal IAV | Mild weight loss and temperature changes | None or mild necrotizing rhinitis and bronchopneumonia with perivascular cuffing | [98,99,100,101,102] |
| 2009 pH1N1 | Mild weight loss | Necrotizing rhinitis, bronchiolitis, perivasculitis, edema, and mild interstitial pneumonia | [101,103,104,105] | ||
| HPAI H5N1 | Not reported | Intranasal route: Bronchiolitis and bronchopneumonia Intragastric route: Interstitial pneumonia | [106] | ||
| Ferret | Mustela putorius furo | Seasonal IAV | Asymptomatic or mild lethargy with sneezing, nasal discharge, and mild weight loss | Conventional intranasal model: Rhinitis and mild bronchiolitis and pneumoniaHigh dose intratracheal model: Moderate rhinitis and severe necrotizing bronchointerstitial pneumonia with edema | [107,108,109,110,111,112] |
| 2009 pH1N1 | Lethargy, anorexia, dyspnea, and elevated body temperature | Necrotizing rhinotracheitis, bronchitis, and bronchiolitis with varying degrees of interstitial pneumonia and diffuse alveolar damage | [77,110,111,112] | ||
| 1918 H1N1 | Weight loss, sneezing, dyspnea, lethargy, and death | Necrotizing rhinitis, bronchiolitis and bronchointerstitial pneumonia with edema | [113,114,115] | ||
| HPAI H5N1 | Lethargy, anorexia, dyspnea, nasal discharge, sneezing, weight loss, elevated body temperature, diarrhea, and neurologic signs (ataxia, torticollis, and hind limb paresis) | Severe necrotizing bronchointerstitial pneumonia with diffuse alveolar damage Meningoencephalitis | [111,116,117,118,119] | ||
| LPAI | Transiently elevated body temperature and weight loss; occasional dyspnea and lethargy | Suppurative rhinitis | [91,120,121] | ||
| NHP | Cynomolgus macaques (most common), rhesus macaques, and common marmosets | Seasonal IAV | Asymptomatic or mild lethargy | Mild bronchointerstitial pneumonia and peribronchiolitis | [122,123] |
| 2009 pH1N1 | Asymptomatic or mildly elevated body temperature and lethargy; tachypnea, dyspnea, and nasal discharge reported for some strains of virus | Suppurative rhinitis in mild cases; necrotizing bronchopneumonia with edema and hyaline membranes in severe cases (strain dependent) | [124,125,126,127,128,129] | ||
| 1918 H1N1 | Anorexia, lethargy, cough, nasal discharge, and tachypnea | Necrotizing bronchointerstitial pneumonia with hemorrhage, edema, and hyaline membranes | [123,130,131] | ||
| HPAI H5N1 | Anorexia, lethargy, cough, tachypnea, elevated body temperature, diarrhea, and thrombocytopenia | Necrotizing bronchointerstitial pneumonia with hemorrhage, edema, hyaline membranes, and type II pneumocyte hyperplasia Lymphoid necrosis and renal tubular necrosis | [130,132,133,134] |
4. Comparative Pathology of Animal Models for Influenza A
4.1. Mouse Models
4.2. Ferret Model
4.3. Hamster Model
4.4. Non-Human Primate Models
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shi, M.; Lin, X.-D.; Chen, X.; Tian, J.-H.; Chen, L.-J.; Li, K.; Wang, W.; Eden, J.-S.; Shen, J.-J.; Liu, L.; et al. The Evolutionary History of Vertebrate RNA Viruses. Nature 2018, 556, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Influenza (Seasonal). Available online: https://www.who.int/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 18 September 2023).
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Primer 2018, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and Ecology of Influenza A Viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef] [PubMed]
- Osterhaus, A.D.M.E.; Rimmelzwaan, G.F.; Martina, B.E.E.; Bestebroer, T.M.; Fouchier, R.A.M. Influenza B Virus in Seals. Science 2000, 288, 1051–1053. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Porter, E.; Lohman, M.; Lu, N.; Peddireddi, L.; Hanzlicek, G.; Marthaler, D.; Liu, X.; Bai, J. Influenza C Virus in Cattle with Respiratory Disease, United States, 2016–2018. Emerg. Infect. Dis. 2018, 24, 1926–1929. [Google Scholar] [CrossRef]
- Guo, Y.J.; Jin, F.G.; Wang, P.; Wang, M.; Zhu, J.M. Isolation of Influenza C Virus from Pigs and Experimental Infection of Pigs with Influenza C Virus. J. Gen. Virol. 1983, 64 Pt 1, 177–182. [Google Scholar] [CrossRef]
- Manuguerra, J.C.; Hannoun, C. Natural Infection of Dogs by Influenza C Virus. Res. Virol. 1992, 143, 199–204. [Google Scholar] [CrossRef]
- Thielen, B.K.; Friedlander, H.; Bistodeau, S.; Shu, B.; Lynch, B.; Martin, K.; Bye, E.; Como-Sabetti, K.; Boxrud, D.; Strain, A.K.; et al. Detection of Influenza C Viruses Among Outpatients and Patients Hospitalized for Severe Acute Respiratory Infection, Minnesota, 2013–2016. Clin. Infect. Dis. 2018, 66, 1092–1098. [Google Scholar] [CrossRef]
- Hause, B.M.; Ducatez, M.; Collin, E.A.; Ran, Z.; Liu, R.; Sheng, Z.; Armien, A.; Kaplan, B.; Chakravarty, S.; Hoppe, A.D.; et al. Isolation of a Novel Swine Influenza Virus from Oklahoma in 2011 Which Is Distantly Related to Human Influenza C Viruses. PLoS Pathog. 2013, 9, e1003176. [Google Scholar] [CrossRef]
- Hause, B.M.; Collin, E.A.; Liu, R.; Huang, B.; Sheng, Z.; Lu, W.; Wang, D.; Nelson, E.A.; Li, F. Characterization of a Novel Influenza Virus in Cattle and Swine: Proposal for a New Genus in the Orthomyxoviridae Family. mBio 2014, 5, e00031-14. [Google Scholar] [CrossRef]
- White, S.K.; Ma, W.; McDaniel, C.J.; Gray, G.C.; Lednicky, J.A. Serologic Evidence of Exposure to Influenza D Virus among Persons with Occupational Contact with Cattle. J. Clin. Virol. 2016, 81, 31–33. [Google Scholar] [CrossRef] [PubMed]
- Influenza Type A Viruses|Avian Influenza (Flu). Available online: https://www.cdc.gov/flu/avianflu/influenza-a-virus-subtypes.htm (accessed on 18 September 2023).
- Brandenburg, B.; Koudstaal, W.; Goudsmit, J.; Klaren, V.; Tang, C.; Bujny, M.V.; Korse, H.J.W.M.; Kwaks, T.; Otterstrom, J.J.; Juraszek, J.; et al. Mechanisms of Hemagglutinin Targeted Influenza Virus Neutralization. PLoS ONE 2013, 8, e80034. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.G.; Govorkova, E.A. Continuing Challenges in Influenza. Ann. N. Y. Acad. Sci. 2014, 1323, 115–139. [Google Scholar] [CrossRef] [PubMed]
- Palese, P. Influenza: Old and New Threats. Nat. Med. 2004, 10, S82–S87. [Google Scholar] [CrossRef] [PubMed]
- Palese, P.; Tumpey, T.M.; Garcia-Sastre, A. What Can We Learn from Reconstructing the Extinct 1918 Pandemic Influenza Virus? Immunity 2006, 24, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zeng, X.; Cui, P.; Yan, C.; Chen, H. Alarming Situation of Emerging H5 and H7 Avian Influenza and Effective Control Strategies. Emerg. Microbes Infect. 2022, 12, 2155072. [Google Scholar] [CrossRef] [PubMed]
- Gambotto, A.; Barratt-Boyes, S.M.; De Jong, M.D.; Neumann, G.; Kawaoka, Y. Human Infection with Highly Pathogenic H5N1 Influenza Virus. Lancet 2008, 371, 1464–1475. [Google Scholar] [CrossRef]
- Killingley, B.; Nguyen-Van-Tam, J. Routes of Influenza Transmission. Influenza Other Respir. Viruses 2013, 7, 42–51. [Google Scholar] [CrossRef]
- Stebbins, S.; Cummings, D.A.T.; Stark, J.H.; Vukotich, C.; Mitruka, K.; Thompson, W.; Rinaldo, C.; Roth, L.; Wagner, M.; Wisniewski, S.R.; et al. Reduction in the Incidence of Influenza A but Not Influenza B Associated with Use of Hand Sanitizer and Cough Hygiene in Schools: A Randomized Controlled Trial. Pediatr. Infect. Dis. J. 2011, 30, 921–926. [Google Scholar] [CrossRef]
- Torner, N.; Soldevila, N.; Garcia, J.J.; Launes, C.; Godoy, P.; Castilla, J.; Domínguez, A.; CIBERESP Cases and Controls in Pandemic Influenza Working Group, Spain. Effectiveness of Non-Pharmaceutical Measures in Preventing Pediatric Influenza: A Case-Control Study. BMC Public Health 2015, 15, 543. [Google Scholar] [CrossRef]
- CDC Seasonal Flu Vaccine Effectiveness Studies|CDC. Available online: https://www.cdc.gov/flu/vaccines-work/effectiveness-studies.htm (accessed on 18 September 2023).
- Influenza—Vaccine Preventable Diseases Surveillance Manual|CDC. Available online: https://www.cdc.gov/vaccines/pubs/surv-manual/chpt06-influenza.html (accessed on 18 September 2023).
- Taubenberger, J.K.; Morens, D.M. The Pathology of Influenza Virus Infections. Annu. Rev. Pathol. 2008, 3, 499–522. [Google Scholar] [CrossRef] [PubMed]
- Minodier, L.; Charrel, R.N.; Ceccaldi, P.-E.; Van der Werf, S.; Blanchon, T.; Hanslik, T.; Falchi, A. Prevalence of Gastrointestinal Symptoms in Patients with Influenza, Clinical Significance, and Pathophysiology of Human Influenza Viruses in Faecal Samples: What Do We Know? Virol. J. 2015, 12, 215. [Google Scholar] [CrossRef] [PubMed]
- Rothberg, M.B.; Haessler, S.D.; Brown, R.B. Complications of Viral Influenza. Am. J. Med. 2008, 121, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Morens, D.M.; Taubenberger, J.K.; Fauci, A.S. Predominant Role of Bacterial Pneumonia as a Cause of Death in Pandemic Influenza: Implications for Pandemic Influenza Preparedness. J. Infect. Dis. 2008, 198, 962–970. [Google Scholar] [CrossRef] [PubMed]
- Dunning, J.; Thwaites, R.S.; Openshaw, P.J.M. Seasonal and Pandemic Influenza: 100 Years of Progress, Still Much to Learn. Mucosal Immunol. 2020, 13, 566–573. [Google Scholar] [CrossRef]
- Writing Committee of the WHO Consultation on Clinical Aspects of Pandemic (H1N1) 2009 Influenza; Bautista, E.; Chotpitayasunondh, T.; Gao, Z.; Harper, S.A.; Shaw, M.; Uyeki, T.M.; Zaki, S.R.; Hayden, F.G.; Hui, D.S.; et al. Clinical Aspects of Pandemic 2009 Influenza A (H1N1) Virus Infection. N. Engl. J. Med. 2010, 362, 1708–1719. [Google Scholar] [CrossRef] [PubMed]
- McAuley, J.L.; Kedzierska, K.; Brown, L.E.; Shanks, G.D. Host Immunological Factors Enhancing Mortality of Young Adults during the 1918 Influenza Pandemic. Front. Immunol. 2015, 6, 419. [Google Scholar] [CrossRef]
- Hers, J.F.P.; Masurel, N.; Mulder, J. Bacteriology and histopathology of the respiratory tract and lungs in fatal Asian influenza. Lancet 1958, 272, 1141–1143. [Google Scholar] [CrossRef]
- Shieh, W.-J.; Blau, D.M.; Denison, A.M.; DeLeon-Carnes, M.; Adem, P.; Bhatnagar, J.; Sumner, J.; Liu, L.; Patel, M.; Batten, B.; et al. 2009 Pandemic Influenza A (H1N1). Am. J. Pathol. 2010, 177, 166–175. [Google Scholar] [CrossRef]
- Mauad, T.; Hajjar, L.A.; Callegari, G.D.; Da Silva, L.F.F.; Schout, D.; Galas, F.R.B.G.; Alves, V.A.F.; Malheiros, D.M.A.C.; Auler, J.O.C.; Ferreira, A.F.; et al. Lung Pathology in Fatal Novel Human Influenza A (H1N1) Infection. Am. J. Respir. Crit. Care Med. 2010, 181, 72–79. [Google Scholar] [CrossRef]
- Gao, H.-N.; Lu, H.-Z.; Cao, B.; Du, B.; Shang, H.; Gan, J.-H.; Lu, S.-H.; Yang, Y.-D.; Fang, Q.; Shen, Y.-Z.; et al. Clinical Findings in 111 Cases of Influenza A (H7N9) Virus Infection. N. Engl. J. Med. 2013, 368, 2277–2285. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.H.; Nguyen, T.L.; Nguyen, T.D.; Luong, T.S.; Pham, P.M.; Van Vinh Chau, N.; Pham, T.S.; Vo, C.D.; Le, T.Q.M.; Ngo, T.T.; et al. Avian Influenza A (H5N1) in 10 Patients in Vietnam. N. Engl. J. Med. 2004, 350, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Yuen, K.; Chan, P.; Peiris, M.; Tsang, D.; Que, T.; Shortridge, K.; Cheung, P.; To, W.; Ho, E.; Sung, R.; et al. Clinical Features and Rapid Viral Diagnosis of Human Disease Associated with Avian Influenza A H5N1 Virus. Lancet 1998, 351, 467–471. [Google Scholar] [CrossRef] [PubMed]
- Oner, A.F.; Bay, A.; Arslan, S.; Akdeniz, H.; Sahin, H.A.; Cesur, Y.; Epcacan, S.; Yilmaz, N.; Deger, I.; Kizilyildiz, B.; et al. Avian Influenza A (H5N1) Infection in Eastern Turkey in 2006. N. Engl. J. Med. 2006, 355, 2179–2185. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Pu, J. Influence of Host Sialic Acid Receptors Structure on the Host Specificity of Influenza Viruses. Viruses 2022, 14, 2141. [Google Scholar] [CrossRef] [PubMed]
- Rogers, G.N.; Paulson, J.C.; Daniels, R.S.; Skehel, J.J.; Wilson, I.A.; Wiley, D.C. Single Amino Acid Substitutions in Influenza Haemagglutinin Change Receptor Binding Specificity. Nature 1983, 304, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Shinya, K.; Ebina, M.; Yamada, S.; Ono, M.; Kasai, N.; Kawaoka, Y. Influenza Virus Receptors in the Human Airway. Nature 2006, 440, 435–436. [Google Scholar] [CrossRef]
- Pillai, S.P.; Lee, C.W. Species and Age Related Differences in the Type and Distribution of Influenza Virus Receptors in Different Tissues of Chickens, Ducks and Turkeys. Virol. J. 2010, 7, 5. [Google Scholar] [CrossRef]
- Knipe, D.M.; Howley, P. Fields Virology; Wolters Kluwer Health: Philadelphia, PA, USA, 2013; ISBN 978-1-4698-7422-7. [Google Scholar]
- Boyd, D.F.; Wilson, T.L.; Thomas, P.G. Chapter Seven—One Hundred Years of (Influenza) Immunopathology. In Advances in Virus Research; Carr, J.P., Roossinck, M.J., Eds.; Immunopathology; Academic Press: Cambridge, MA, USA, 2020; Volume 107, pp. 247–284. [Google Scholar]
- Tang, B.M.; Shojaei, M.; Teoh, S.; Meyers, A.; Ho, J.; Ball, T.B.; Keynan, Y.; Pisipati, A.; Kumar, A.; Eisen, D.P.; et al. Neutrophils-Related Host Factors Associated with Severe Disease and Fatality in Patients with Influenza Infection. Nat. Commun. 2019, 10, 3422. [Google Scholar] [CrossRef]
- Guan, W.; Yang, Z.; Wu, N.C.; Lee, H.H.Y.; Li, Y.; Jiang, W.; Shen, L.; Wu, D.C.; Chen, R.; Zhong, N.; et al. Clinical Correlations of Transcriptional Profile in Patients Infected with Avian Influenza H7N9 Virus. J. Infect. Dis. 2018, 218, 1238–1248. [Google Scholar] [CrossRef]
- Fiore-Gartland, A.; Panoskaltsis-Mortari, A.; Agan, A.A.; Mistry, A.J.; Thomas, P.G.; Matthay, M.A.; PALISI PICFlu Investigators; Hertz, T.; Randolph, A.G. Cytokine Profiles of Severe Influenza Virus-Related Complications in Children. Front. Immunol. 2017, 8, 1423. [Google Scholar] [CrossRef] [PubMed]
- Bradley-Stewart, A.; Jolly, L.; Adamson, W.; Gunson, R.; Frew-Gillespie, C.; Templeton, K.; Aitken, C.; Carman, W.; Cameron, S.; McSharry, C. Cytokine Responses in Patients with Mild or Severe Influenza A(H1N1)Pdm09. J. Clin. Virol. 2013, 58, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Oshansky, C.M.; Gartland, A.J.; Wong, S.-S.; Jeevan, T.; Wang, D.; Roddam, P.L.; Caniza, M.A.; Hertz, T.; Devincenzo, J.P.; Webby, R.J.; et al. Mucosal Immune Responses Predict Clinical Outcomes during Influenza Infection Independently of Age and Viral Load. Am. J. Respir. Crit. Care Med. 2014, 189, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Guarner, J.; Falcón-Escobedo, R. Comparison of the Pathology Caused by H1N1, H5N1, and H3N2 Influenza Viruses. Arch. Med. Res. 2009, 40, 655–661. [Google Scholar] [CrossRef] [PubMed]
- To, K.-F.; Chan, P.K.S.; Chan, K.-F.; Lee, W.-K.; Lam, W.-Y.; Wong, K.-F.; Tang, N.L.S.; Tsang, D.N.C.; Sung, R.Y.T.; Buckley, T.A.; et al. Pathology of Fatal Human Infection Associated with Avian Influenza A H5N1 Virus. J. Med. Virol. 2001, 63, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Guarner, J.; Paddock, C.D.; Shieh, W.-J.; Packard, M.M.; Patel, M.; Montague, J.L.; Uyeki, T.M.; Bhat, N.; Balish, A.; Lindstrom, S.; et al. Histopathologic and Immunohistochemical Features of Fatal Influenza Virus Infection in Children during the 2003-2004 Season. Clin. Infect. Dis. 2006, 43, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Lucke, B.; Kime, E. PATHOLOGIC ANATOMY AND BACTERIOLOGY OF INFLUENZA: EPIDEMIC OF AUTUMN, 1918. Arch. Intern. Med. 1919, 24, 154–237. [Google Scholar] [CrossRef]
- Lindsay, M.I.; Herrmann, E.C.; Morrow, G.W.; Brown, A.L. Hong Kong Influenza: Clinical, Microbiologic, and Pathologic Features in 127 Cases. JAMA 1970, 214, 1825–1832. [Google Scholar] [CrossRef]
- The ARDS Definition Task Force. Acute Respiratory Distress Syndrome: The Berlin Definition. JAMA 2012, 307, 2526–2533. [Google Scholar] [CrossRef]
- Avian Influenza Weekly Update Number 924. Available online: https://cdn.who.int/media/docs/default-source/wpro---documents/emergency/surveillance/avian-influenza/ai_20231203.pdf?sfvrsn=5f006f99_123 (accessed on 6 December 2023).
- Peiris, J.S.M.; Yu, W.C.; Leung, C.W.; Cheung, C.Y.; Ng, W.F.; Nicholls, J.M.; Ng, T.K.; Chan, K.H.; Lai, S.T.; Lim, W.L.; et al. Re-Emergence of Fatal Human Influenza A Subtype H5N1 Disease. Lancet 2004, 363, 617–619. [Google Scholar] [CrossRef]
- Ungchusak, K.; Auewarakul, P.; Dowell, S.F.; Kitphati, R.; Auwanit, W.; Puthavathana, P.; Uiprasertkul, M.; Boonnak, K.; Pittayawonganon, C.; Cox, N.J.; et al. Probable Person-to-Person Transmission of Avian Influenza A (H5N1). N. Engl. J. Med. 2005, 352, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Chotpitayasunondh, T.; Ungchusak, K.; Hanshaoworakul, W.; Chunsuthiwat, S.; Sawanpanyalert, P.; Kijphati, R.; Lochindarat, S.; Srisan, P.; Suwan, P.; Osotthanakorn, Y.; et al. Human Disease from Influenza A (H5N1), Thailand, 2004. Emerg. Infect. Dis. 2005, 11, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.K.S. Outbreak of Avian Influenza A(H5N1) Virus Infection in Hong Kong in 1997. Clin. Infect. Dis. 2002, 34 (Suppl. S2), S58–S64. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Xie, Z.; Gao, Z.; Liu, J.; Korteweg, C.; Ye, J.; Lau, L.T.; Lu, J.; Gao, Z.; Zhang, B.; et al. H5N1 Infection of the Respiratory Tract and beyond: A Molecular Pathology Study. Lancet 2007, 370, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Uiprasertkul, M.; Puthavathana, P.; Sangsiriwut, K.; Pooruk, P.; Srisook, K.; Peiris, M.; Nicholls, J.M.; Chokephaibulkit, K.; Vanprapar, N.; Auewarakul, P. Influenza A H5N1 Replication Sites in Humans. Emerg. Infect. Dis. 2005, 11, 1036–1041. [Google Scholar] [CrossRef]
- Uiprasertkul, M.; Kitphati, R.; Puthavathana, P.; Kriwong, R.; Kongchanagul, A.; Ungchusak, K.; Angkasekwinai, S.; Chokephaibulkit, K.; Srisook, K.; Vanprapar, N.; et al. Apoptosis and Pathogenesis of Avian Influenza A (H5N1) Virus in Humans. Emerg. Infect. Dis. 2007, 13, 708–712. [Google Scholar] [CrossRef] [PubMed]
- Chokephaibulkit, K.; Uiprasertkul, M.; Puthavathana, P.; Chearskul, P.; Auewarakul, P.; Dowell, S.F.; Vanprapar, N. A Child with Avian Influenza A (H5N1) Infection. Pediatr. Infect. Dis. J. 2005, 24, 162–166. [Google Scholar] [CrossRef]
- Ku, A.S.; Chan, L.T. The First Case of H5N1 Avian Influenza Infection in a Human with Complications of Adult Respiratory Distress Syndrome and Reye’s Syndrome. J. Paediatr. Child Health 1999, 35, 207–209. [Google Scholar] [CrossRef]
- De Jong, M.D.; Cam, B.V.; Qui, P.T.; Hien, V.M.; Thanh, T.T.; Hue, N.B.; Beld, M.; Phuong, L.T.; Khanh, T.H.; Chau, N.V.V.; et al. Fatal Avian Influenza A (H5N1) in a Child Presenting with Diarrhea Followed by Coma. N. Engl. J. Med. 2005, 352, 686–691. [Google Scholar] [CrossRef]
- De Jong, M.D.; Simmons, C.P.; Thanh, T.T.; Hien, V.M.; Smith, G.J.D.; Chau, T.N.B.; Hoang, D.M.; Van Vinh Chau, N.; Khanh, T.H.; Dong, V.C.; et al. Fatal Outcome of Human Influenza A (H5N1) Is Associated with High Viral Load and Hypercytokinemia. Nat. Med. 2006, 12, 1203–1207. [Google Scholar] [CrossRef]
- Cheung, C.Y.; Poon, L.L.M.; Lau, A.S.; Luk, W.; Lau, Y.L.; Shortridge, K.F.; Gordon, S.; Guan, Y.; Peiris, J.S.M. Induction of Proinflammatory Cytokines in Human Macrophages by Influenza A (H5N1) Viruses: A Mechanism for the Unusual Severity of Human Disease? Lancet 2002, 360, 1831–1837. [Google Scholar] [CrossRef] [PubMed]
- Korteweg, C.; Gu, J. Pathology, Molecular Biology, and Pathogenesis of Avian Influenza A (H5N1) Infection in Humans. Am. J. Pathol. 2008, 172, 1155–1170. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Wang, Z.; Chen, Y.; Ding, W.; Jia, H.; Chan, J.F.-W.; To, K.K.-W.; Chen, H.; Yang, Y.; Liang, W.; et al. Clinical, Virological, and Histopathological Manifestations of Fatal Human Infections by Avian Influenza A(H7N9) Virus. Clin. Infect. Dis. 2013, 57, 1449–1457. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-B.; Li, H.-Y.; Liu, J.-F.; Lan, C.-Q.; Lin, Q.-H.; Chen, S.-X.; Zhang, H.-Y.; Wang, X.-H.; Lin, X.; Pan, J.-G.; et al. Histopathological Findings in a Critically Ill Patient with Avian Influenza A (H7N9). J. Thorac. Dis. 2015, 7, E672–E677. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, B.; Błazejewska, P.; Hessmann, M.; Bruder, D.; Geffers, R.; Mauel, S.; Gruber, A.D.; Schughart, K. Host Genetic Background Strongly Influences the Response to Influenza a Virus Infections. PLoS ONE 2009, 4, e4857. [Google Scholar] [CrossRef]
- Pica, N.; Iyer, A.; Ramos, I.; Bouvier, N.M.; Fernandez-Sesma, A.; García-Sastre, A.; Lowen, A.C.; Palese, P.; Steel, J. The DBA.2 Mouse Is Susceptible to Disease Following Infection with a Broad, but Limited, Range of Influenza A and B Viruses. J. Virol. 2011, 85, 12825–12829. [Google Scholar] [CrossRef] [PubMed]
- Fukushi, M.; Ito, T.; Oka, T.; Kitazawa, T.; Miyoshi-Akiyama, T.; Kirikae, T.; Yamashita, M.; Kudo, K. Serial Histopathological Examination of the Lungs of Mice Infected with Influenza A Virus PR8 Strain. PLoS ONE 2011, 6, e21207. [Google Scholar] [CrossRef]
- Chen, J.; Fang, F.; Li, X.; Chang, H.; Chen, Z. Protection against Influenza Virus Infection in BALB/c Mice Immunized with a Single Dose of Neuraminidase-Expressing DNAs by Electroporation. Vaccine 2005, 23, 4322–4328. [Google Scholar] [CrossRef]
- Belser, J.A.; Wadford, D.A.; Pappas, C.; Gustin, K.M.; Maines, T.R.; Pearce, M.B.; Zeng, H.; Swayne, D.E.; Pantin-Jackwood, M.; Katz, J.M.; et al. Pathogenesis of Pandemic Influenza A (H1N1) and Triple-Reassortant Swine Influenza A (H1) Viruses in Mice. J. Virol. 2010, 84, 4194–4203. [Google Scholar] [CrossRef]
- Maines, T.R.; Jayaraman, A.; Belser, J.A.; Wadford, D.A.; Pappas, C.; Zeng, H.; Gustin, K.M.; Pearce, M.B.; Viswanathan, K.; Shriver, Z.H.; et al. Transmission and Pathogenesis of Swine-Origin 2009 A(H1N1) Influenza Viruses in Ferrets and Mice. Science 2009, 325, 484–487. [Google Scholar] [CrossRef]
- Manicassamy, B.; Medina, R.A.; Hai, R.; Tsibane, T.; Stertz, S.; Nistal-Villán, E.; Palese, P.; Basler, C.F.; García-Sastre, A. Protection of Mice against Lethal Challenge with 2009 H1N1 Influenza A Virus by 1918-like and Classical Swine H1N1 Based Vaccines. PLoS Pathog. 2010, 6, e1000745. [Google Scholar] [CrossRef] [PubMed]
- Groves, H.T.; McDonald, J.U.; Langat, P.; Kinnear, E.; Kellam, P.; McCauley, J.; Ellis, J.; Thompson, C.; Elderfield, R.; Parker, L.; et al. Mouse Models of Influenza Infection with Circulating Strains to Test Seasonal Vaccine Efficacy. Front. Immunol. 2018, 9, 126. [Google Scholar] [CrossRef] [PubMed]
- Tumpey, T.M.; Basler, C.F.; Aguilar, P.V.; Zeng, H.; Solórzano, A.; Swayne, D.E.; Cox, N.J.; Katz, J.M.; Taubenberger, J.K.; Palese, P.; et al. Characterization of the Reconstructed 1918 Spanish Influenza Pandemic Virus. Science 2005, 310, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Tumpey, T.M.; Szretter, K.J.; Van Hoeven, N.; Katz, J.M.; Kochs, G.; Haller, O.; García-Sastre, A.; Staeheli, P. The Mx1 Gene Protects Mice against the Pandemic 1918 and Highly Lethal Human H5N1 Influenza Viruses. J. Virol. 2007, 81, 10818–10821. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Tumpey, T.M.; Morken, T.; Zaki, S.R.; Cox, N.J.; Katz, J.M. A Mouse Model for the Evaluation of Pathogenesis and Immunity to Influenza A (H5N1) Viruses Isolated from Humans. J. Virol. 1999, 73, 5903–5911. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Watanabe, S.; Ito, T.; Goto, H.; Wells, K.; McGregor, M.; Cooley, A.J.; Kawaoka, Y. Biological Heterogeneity, Including Systemic Replication in Mice, of H5N1 Influenza A Virus Isolates from Humans in Hong Kong. J. Virol. 1999, 73, 3184–3189. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, H.; Itamura, S.; Iwasaki, T.; Kurata, T.; Tashiro, M. Characterization of Human Influenza A (H5N1) Virus Infection in Mice: Neuro-, Pneumo- and Adipotropic Infection. J. Gen. 2000, 81, 2503–2510. [Google Scholar] [CrossRef]
- Gubareva, L.V.; McCullers, J.A.; Bethell, R.C.; Webster, R.G. Characterization of Influenza A/HongKong/156/97 (H5N1) Virus in a Mouse Model and Protective Effect of Zanamivir on H5N1 Infection in Mice. J. Infect. Dis. 1998, 178, 1592–1596. [Google Scholar] [CrossRef]
- Dybing, J.K.; Schultz-Cherry, S.; Swayne, D.E.; Suarez, D.L.; Perdue, M.L. Distinct Pathogenesis of Hong Kong-Origin H5N1 Viruses in Mice Compared to That of Other Highly Pathogenic H5 Avian Influenza Viruses. J. Virol. 2000, 74, 1443–1450. [Google Scholar] [CrossRef]
- Tanaka, H.; Park, C.-H.; Ninomiya, A.; Ozaki, H.; Takada, A.; Umemura, T.; Kida, H. Neurotropism of the 1997 Hong Kong H5N1 Influenza Virus in Mice. Vet. Microbiol. 2003, 95, 1–13. [Google Scholar] [CrossRef]
- Boon, A.C.M.; Finkelstein, D.; Zheng, M.; Liao, G.; Allard, J.; Klumpp, K.; Webster, R.; Peltz, G.; Webby, R.J. H5N1 Influenza Virus Pathogenesis in Genetically Diverse Mice Is Mediated at the Level of Viral Load. mBio 2011, 2, e00171-11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Liu, C.; Kou, Z.; Gao, T.; Pan, T.; Wu, X.; Yu, H.; Guo, Y.; Zeng, Y.; Du, L.; et al. Differences in the Pathogenicity and Inflammatory Responses Induced by Avian Influenza A/H7N9 Virus Infection in BALB/c and C57BL/6 Mouse Models. PLoS ONE 2014, 9, e92987. [Google Scholar] [CrossRef] [PubMed]
- Mok, C.K.P.; Lee, H.H.Y.; Chan, M.C.W.; Sia, S.F.; Lestra, M.; Nicholls, J.M.; Zhu, H.; Guan, Y.; Peiris, J.M.S. Pathogenicity of the Novel A/H7N9 Influenza Virus in Mice. mBio 2013, 4, e00362-13. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Gustin, K.M.; Pearce, M.B.; Maines, T.R.; Zeng, H.; Pappas, C.; Sun, X.; Carney, P.J.; Villanueva, J.M.; Stevens, J.; et al. Pathogenesis and Transmission of Avian Influenza A (H7N9) Virus in Ferrets and Mice. Nature 2013, 501, 556–559. [Google Scholar] [CrossRef] [PubMed]
- Meliopoulos, V.A.; Karlsson, E.A.; Kercher, L.; Cline, T.; Freiden, P.; Duan, S.; Vogel, P.; Webby, R.J.; Guan, Y.; Peiris, M.; et al. Human H7N9 and H5N1 Influenza Viruses Differ in Induction of Cytokines and Tissue Tropism. J. Virol. 2014, 88, 12982–12991. [Google Scholar] [CrossRef]
- Lee, Y.-N.; Lee, D.-H.; Cheon, S.-H.; Park, Y.-R.; Baek, Y.-G.; Si, Y.-J.; Kye, S.-J.; Lee, E.-K.; Heo, G.-B.; Bae, Y.-C.; et al. Genetic Characteristics and Pathogenesis of H5 Low Pathogenic Avian Influenza Viruses from Wild Birds and Domestic Ducks in South Korea. Sci. Rep. 2020, 10, 12151. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Lu, X.; Maines, T.R.; Smith, C.; Li, Y.; Donis, R.O.; Katz, J.M.; Tumpey, T.M. Pathogenesis of Avian Influenza (H7) Virus Infection in Mice and Ferrets: Enhanced Virulence of Eurasian H7N7 Viruses Isolated from Humans. J. Virol. 2007, 81, 11139–11147. [Google Scholar] [CrossRef]
- Driskell, E.A.; Jones, C.A.; Stallknecht, D.E.; Howerth, E.W.; Tompkins, S.M. Avian Influenza Virus Isolates from Wild Birds Replicate and Cause Disease in a Mouse Model of Infection. Virology 2010, 399, 280–289. [Google Scholar] [CrossRef]
- Gillim-Ross, L.; Santos, C.; Chen, Z.; Aspelund, A.; Yang, C.-F.; Ye, D.; Jin, H.; Kemble, G.; Subbarao, K. Avian Influenza H6 Viruses Productively Infect and Cause Illness in Mice and Ferrets. J. Virol. 2008, 82, 10854–10863. [Google Scholar] [CrossRef]
- Joseph, T.; McAuliffe, J.; Lu, B.; Jin, H.; Kemble, G.; Subbarao, K. Evaluation of Replication and Pathogenicity of Avian Influenza a H7 Subtype Viruses in a Mouse Model. J. Virol. 2007, 81, 10558–10566. [Google Scholar] [CrossRef]
- Heath, A.W.; Addison, C.; Ali, M.; Teale, D.; Potter, C.W. In Vivo and in Vitro Hamster Models in the Assessment of Virulence of Recombinant Influenza Viruses. Antiviral Res. 1983, 3, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.J.; Teh, C.Z.; Jennings, R.; Potter, C.W. Transmissibility of Influenza Viruses in Hamsters. Arch. Virol. 1982, 72, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Jennings, R.; Denton, M.D.; Potter, C.W. The Hamster as an Experimental Animal for the Study of Influenza. I. The Role of Antibody in Protection. Med. Microbiol. Immunol. 1976, 162, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Iwatsuki-Horimoto, K.; Nakajima, N.; Ichiko, Y.; Sakai-Tagawa, Y.; Noda, T.; Hasegawa, H.; Kawaoka, Y. Syrian Hamster as an Animal Model for the Study of Human Influenza Virus Infection. J. Virol. 2018, 92, e01693-17. [Google Scholar] [CrossRef] [PubMed]
- Paterson, J.; Ryan, K.A.; Morley, D.; Jones, N.J.; Yeates, P.; Hall, Y.; Whittaker, C.J.; Salguero, F.J.; Marriott, A.C. Infection with Seasonal H1N1 Influenza Results in Comparable Disease Kinetics and Host Immune Responses in Ferrets and Golden Syrian Hamsters. Pathogens 2023, 12, 668. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.J.; Lee, A.C.-Y.; Chan, J.F.-W.; Liu, F.; Li, C.; Chen, Y.; Chu, H.; Lau, S.-Y.; Wang, P.; Chan, C.C.-S.; et al. Coinfection by Severe Acute Respiratory Syndrome Coronavirus 2 and Influenza A(H1N1)Pdm09 Virus Enhances the Severity of Pneumonia in Golden Syrian Hamsters. Clin. Infect. Dis. 2021, 72, e978–e992. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Kang, J.-A.; Lyoo, K.-S.; Le, T.B.; Yeo, Y.H.; Wong, S.-S.; Na, W.; Song, D.; Webby, R.J.; Zanin, M.; et al. Severe Acute Respiratory Syndrome Coronavirus 2 and Influenza A Virus Co-Infection Alters Viral Tropism and Haematological Composition in Syrian Hamsters. Transbound. Emerg. Dis. 2022, 69, e3297–e3304. [Google Scholar] [CrossRef] [PubMed]
- Oishi, K.; Horiuchi, S.; Minkoff, J.M.; Ten Oever, B.R. The Host Response to Influenza A Virus Interferes with SARS-CoV-2 Replication during Coinfection. J. Virol. 2022, 96, e0076522. [Google Scholar] [CrossRef]
- Shinya, K.; Makino, A.; Tanaka, H.; Hatta, M.; Watanabe, T.; Le, M.Q.; Imai, H.; Kawaoka, Y. Systemic Dissemination of H5N1 Influenza A Viruses in Ferrets and Hamsters after Direct Intragastric Inoculation. J. Virol. 2011, 85, 4673–4678. [Google Scholar] [CrossRef]
- Pappas, C.; Yang, H.; Carney, P.J.; Pearce, M.B.; Katz, J.M.; Stevens, J.; Tumpey, T.M. Assessment of Transmission, Pathogenesis and Adaptation of H2 Subtype Influenza Viruses in Ferrets. Virology 2015, 477, 61–71. [Google Scholar] [CrossRef]
- Huang, S.S.H.; Banner, D.; Fang, Y.; Ng, D.C.K.; Kanagasabai, T.; Kelvin, D.J.; Kelvin, A.A. Comparative Analyses of Pandemic H1N1 and Seasonal H1N1, H3N2, and Influenza B Infections Depict Distinct Clinical Pictures in Ferrets. PLoS ONE 2011, 6, e27512. [Google Scholar] [CrossRef] [PubMed]
- Svitek, N.; Rudd, P.A.; Obojes, K.; Pillet, S.; Von Messling, V. Severe Seasonal Influenza in Ferrets Correlates with Reduced Interferon and Increased IL-6 Induction. Virology 2008, 376, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Munster, V.J.; De Wit, E.; Van den Brand, J.M.A.; Herfst, S.; Schrauwen, E.J.A.; Bestebroer, T.M.; Van de Vijver, D.; Boucher, C.A.; Koopmans, M.; Rimmelzwaan, G.F.; et al. Pathogenesis and Transmission of Swine-Origin 2009 A(H1N1) Influenza Virus in Ferrets. Science 2009, 325, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Van den Brand, J.M.A.; Stittelaar, K.J.; Van Amerongen, G.; Rimmelzwaan, G.F.; Simon, J.; De Wit, E.; Munster, V.; Bestebroer, T.; Fouchier, R.A.M.; Kuiken, T.; et al. Severity of Pneumonia Due to New H1N1 Influenza Virus in Ferrets Is Intermediate between That Due to Seasonal H1N1 Virus and Highly Pathogenic Avian Influenza H5N1 Virus. J. Infect. Dis. 2010, 201, 993–999. [Google Scholar] [CrossRef]
- Van den Brand, J.M.A.; Stittelaar, K.J.; Van Amerongen, G.; Reperant, L.; De Waal, L.; Osterhaus, A.D.M.E.; Kuiken, T. Comparison of Temporal and Spatial Dynamics of Seasonal H3N2, Pandemic H1N1 and Highly Pathogenic Avian Influenza H5N1 Virus Infections in Ferrets. PLoS ONE 2012, 7, e42343. [Google Scholar] [CrossRef] [PubMed]
- Tumpey, T.M.; Maines, T.R.; Van Hoeven, N.; Glaser, L.; Solórzano, A.; Pappas, C.; Cox, N.J.; Swayne, D.E.; Palese, P.; Katz, J.M.; et al. A Two-Amino Acid Change in the Hemagglutinin of the 1918 Influenza Virus Abolishes Transmission. Science 2007, 315, 655–659. [Google Scholar] [CrossRef]
- De Wit, E.; Siegers, J.Y.; Cronin, J.M.; Weatherman, S.; Van den Brand, J.M.; Leijten, L.M.; Van Run, P.; Begeman, L.; Van den Ham, H.-J.; Andeweg, A.C.; et al. 1918 H1N1 Influenza Virus Replicates and Induces Proinflammatory Cytokine Responses in Extrarespiratory Tissues of Ferrets. J. Infect. Dis. 2018, 217, 1237–1246. [Google Scholar] [CrossRef]
- Watanabe, T.; Watanabe, S.; Shinya, K.; Kim, J.H.; Hatta, M.; Kawaoka, Y. Viral RNA Polymerase Complex Promotes Optimal Growth of 1918 Virus in the Lower Respiratory Tract of Ferrets. Proc. Natl. Acad. Sci. USA 2009, 106, 588–592. [Google Scholar] [CrossRef]
- Maines, T.R.; Chen, L.-M.; Matsuoka, Y.; Chen, H.; Rowe, T.; Ortin, J.; Falcón, A.; Hien, N.T.; Mai, L.Q.; Sedyaningsih, E.R.; et al. Lack of Transmission of H5N1 Avian–Human Reassortant Influenza Viruses in a Ferret Model. Proc. Natl. Acad. Sci. USA 2006, 103, 12121–12126. [Google Scholar] [CrossRef]
- Imai, M.; Watanabe, T.; Kiso, M.; Nakajima, N.; Yamayoshi, S.; Iwatsuki-Horimoto, K.; Hatta, M.; Yamada, S.; Ito, M.; Sakai-Tagawa, Y.; et al. A Highly Pathogenic Avian H7N9 Influenza Virus Isolated from A Human Is Lethal in Some Ferrets Infected via Respiratory Droplets. Cell Host Microbe 2017, 22, 615–626.e8. [Google Scholar] [CrossRef]
- Pearce, M.B.; Pappas, C.; Gustin, K.M.; Davis, C.T.; Pantin-Jackwood, M.J.; Swayne, D.E.; Maines, T.R.; Belser, J.A.; Tumpey, T.M. Enhanced Virulence of Clade 2.3.2.1 Highly Pathogenic Avian Influenza A H5N1 Viruses in Ferrets. Virology 2017, 502, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Zitzow, L.A.; Rowe, T.; Morken, T.; Shieh, W.-J.; Zaki, S.; Katz, J.M. Pathogenesis of Avian Influenza A (H5N1) Viruses in Ferrets. J. Virol. 2002, 76, 4420–4429. [Google Scholar] [CrossRef] [PubMed]
- Richard, M.; Schrauwen, E.J.A.; De Graaf, M.; Bestebroer, T.M.; Spronken, M.I.J.; Van Boheemen, S.; De Meulder, D.; Lexmond, P.; Linster, M.; Herfst, S.; et al. Limited Airborne Transmission of H7N9 Influenza A Virus between Ferrets. Nature 2013, 501, 560–563. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Pulit-Penaloza, J.A.; Sun, X.; Brock, N.; Pappas, C.; Creager, H.M.; Zeng, H.; Tumpey, T.M.; Maines, T.R. A Novel A(H7N2) Influenza Virus Isolated from a Veterinarian Caring for Cats in a New York City Animal Shelter Causes Mild Disease and Transmits Poorly in the Ferret Model. J. Virol. 2017, 91, e00672-17. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.D.; Olsen, R.J.; LaCasse, R.A.; Safronetz, D.; Ashraf, M.; Porter, A.R.; Braughton, K.R.; Feldmann, F.; Clifton, D.R.; Kash, J.C.; et al. Seasonal H3N2 Influenza A Virus Fails to Enhance Staphylococcus Aureus Co-Infection in a Non-Human Primate Respiratory Tract Infection Model. Virulence 2013, 4, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Kobasa, D.; Jones, S.M.; Shinya, K.; Kash, J.C.; Copps, J.; Ebihara, H.; Hatta, Y.; Hyun Kim, J.; Halfmann, P.; Hatta, M.; et al. Aberrant Innate Immune Response in Lethal Infection of Macaques with the 1918 Influenza Virus. Nature 2007, 445, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Skinner, J.A.; Zurawski, S.M.; Sugimoto, C.; Vinet-Oliphant, H.; Vinod, P.; Xue, Y.; Russell-Lodrigue, K.; Albrecht, R.A.; García-Sastre, A.; Salazar, A.M.; et al. Immunologic Characterization of a Rhesus Macaque H1N1 Challenge Model for Candidate Influenza Virus Vaccine Assessment. Clin. Vaccine Immunol. CVI 2014, 21, 1668–1680. [Google Scholar] [CrossRef] [PubMed]
- Josset, L.; Engelmann, F.; Haberthur, K.; Kelly, S.; Park, B.; Kawoaka, Y.; García-Sastre, A.; Katze, M.G.; Messaoudi, I. Increased Viral Loads and Exacerbated Innate Host Responses in Aged Macaques Infected with the 2009 Pandemic H1N1 Influenza A Virus. J. Virol. 2012, 86, 11115–11127. [Google Scholar] [CrossRef]
- Marriott, A.C.; Dennis, M.; Kane, J.A.; Gooch, K.E.; Hatch, G.; Sharpe, S.; Prevosto, C.; Leeming, G.; Zekeng, E.-G.; Staples, K.J.; et al. Influenza A Virus Challenge Models in Cynomolgus Macaques Using the Authentic Inhaled Aerosol and Intra-Nasal Routes of Infection. PLoS ONE 2016, 11, e0157887. [Google Scholar] [CrossRef]
- Safronetz, D.; Rockx, B.; Feldmann, F.; Belisle, S.E.; Palermo, R.E.; Brining, D.; Gardner, D.; Proll, S.C.; Marzi, A.; Tsuda, Y.; et al. Pandemic Swine-Origin H1N1 Influenza A Virus Isolates Show Heterogeneous Virulence in Macaques. J. Virol. 2011, 85, 1214–1223. [Google Scholar] [CrossRef]
- Moncla, L.H.; Ross, T.M.; Dinis, J.M.; Weinfurter, J.T.; Mortimer, T.D.; Schultz-Darken, N.; Brunner, K.; Capuano, S.V., III; Boettcher, C.; Post, J.; et al. A Novel Nonhuman Primate Model for Influenza Transmission. PLoS ONE 2013, 8, e78750. [Google Scholar] [CrossRef] [PubMed]
- Mooij, P.; Koopman, G.; Mortier, D.; Van Heteren, M.; Oostermeijer, H.; Fagrouch, Z.; De Laat, R.; Kobinger, G.; Li, Y.; Remarque, E.J.; et al. Pandemic Swine-Origin H1N1 Influenza Virus Replicates to Higher Levels and Induces More Fever and Acute Inflammatory Cytokines in Cynomolgus versus Rhesus Monkeys and Can Replicate in Common Marmosets. PLoS ONE 2015, 10, e0126132. [Google Scholar] [CrossRef] [PubMed]
- Cillóniz, C.; Shinya, K.; Peng, X.; Korth, M.J.; Proll, S.C.; Aicher, L.D.; Carter, V.S.; Chang, J.H.; Kobasa, D.; Feldmann, F.; et al. Lethal Influenza Virus Infection in Macaques Is Associated with Early Dysregulation of Inflammatory Related Genes. PLoS Pathog. 2009, 5, e1000604. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, F.; Kobasa, D.; Embury-Hyatt, C.; Grolla, A.; Taylor, T.; Kiso, M.; Kakugawa, S.; Gren, J.; Jones, S.M.; Kawaoka, Y.; et al. Oseltamivir Is Effective against 1918 Influenza Virus Infection of Macaques but Vulnerable to Escape. mBio 2019, 10, e02059-19. [Google Scholar] [CrossRef] [PubMed]
- Baskin, C.R.; Bielefeldt-Ohmann, H.; Tumpey, T.M.; Sabourin, P.J.; Long, J.P.; García-Sastre, A.; Tolnay, A.-E.; Albrecht, R.; Pyles, J.A.; Olson, P.H.; et al. Early and Sustained Innate Immune Response Defines Pathology and Death in Nonhuman Primates Infected by Highly Pathogenic Influenza Virus. Proc. Natl. Acad. Sci. USA 2009, 106, 3455–3460. [Google Scholar] [CrossRef] [PubMed]
- Rimmelzwaan, G.F.; Kuiken, T.; Van Amerongen, G.; Bestebroer, T.M.; Fouchier, R.A.; Osterhaus, A.D. Pathogenesis of Influenza A (H5N1) Virus Infection in a Primate Model. J. Virol. 2001, 75, 6687–6691. [Google Scholar] [CrossRef] [PubMed]
- Kuiken, T.; Rimmelzwaan, G.F.; Van Amerongen, G.; Osterhaus, A.D.M.E. Pathology of Human Influenza A (H5N1) Virus Infection in Cynomolgus Macaques (Macaca Fascicularis). Vet. Pathol. 2003, 40, 304–310. [Google Scholar] [CrossRef]
- Hirst, G.K. Studies on the mechanism of adaptation of influenza virus to mice. J. Exp. Med. 1947, 86, 357–366. [Google Scholar] [CrossRef]
- Wyde, P.R.; Couch, R.B.; Mackler, B.F.; Cate, T.R.; Levy, B.M. Effects of Low- and High-Passage Influenza Virus Infection in Normal and Nude Mice. Infect. Immun. 1977, 15, 221–229. [Google Scholar] [CrossRef]
- Staeheli, P.; Grob, R.; Meier, E.; Sutcliffe, J.G.; Haller, O. Influenza Virus-Susceptible Mice Carry Mx Genes with a Large Deletion or a Nonsense Mutation. Mol. Cell. Biol. 1988, 8, 4518–4523. [Google Scholar] [CrossRef]
- Haller, O.; Acklin, M.; Staeheli, P. Influenza Virus Resistance of Wild Mice: Wild-Type and Mutant Mx Alleles Occur at Comparable Frequencies. J. Interferon Res. 1987, 7, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Dhanwani, R.; Zhou, Y.; Huang, Q.; Verma, V.; Dileepan, M.; Ly, H.; Liang, Y. A Novel Live Pichinde Virus-Based Vaccine Vector Induces Enhanced Humoral and Cellular Immunity after a Booster Dose. J. Virol. 2016, 90, 2551–2560. [Google Scholar] [CrossRef] [PubMed]
- Mozdzanowska, K.; Maiese, K.; Gerhard, W. Th Cell-Deficient Mice Control Influenza Virus Infection More Effectively Than Th- and B Cell-Deficient Mice: Evidence for a Th-Independent Contribution by B Cells to Virus Clearance1. J. Immunol. 2000, 164, 2635–2643. [Google Scholar] [CrossRef] [PubMed]
- Kamal, R.P.; Katz, J.M.; York, I.A. Molecular Determinants of Influenza Virus Pathogenesis in Mice. Curr. Top. Microbiol. Immunol. 2014, 385, 243–274. [Google Scholar] [CrossRef] [PubMed]
- Ibricevic, A.; Pekosz, A.; Walter, M.J.; Newby, C.; Battaile, J.T.; Brown, E.G.; Holtzman, M.J.; Brody, S.L. Influenza Virus Receptor Specificity and Cell Tropism in Mouse and Human Airway Epithelial Cells. J. Virol. 2006, 80, 7469–7480. [Google Scholar] [CrossRef] [PubMed]
- Van Riel, D.; Munster, V.J.; De Wit, E.; Rimmelzwaan, G.F.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Kuiken, T. Human and Avian Influenza Viruses Target Different Cells in the Lower Respiratory Tract of Humans and Other Mammals. Am. J. Pathol. 2007, 171, 1215–1223. [Google Scholar] [CrossRef]
- Van Riel, D.; Den Bakker, M.A.; Leijten, L.M.E.; Chutinimitkul, S.; Munster, V.J.; De Wit, E.; Rimmelzwaan, G.F.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Kuiken, T. Seasonal and Pandemic Human Influenza Viruses Attach Better to Human Upper Respiratory Tract Epithelium than Avian Influenza Viruses. Am. J. Pathol. 2010, 176, 1614–1618. [Google Scholar] [CrossRef]
- Meijer, W.J.; Linn, F.H.H.; Wensing, A.M.J.; Leavis, H.L.; Van Riel, D.; GeurtsvanKessel, C.H.; Wattjes, M.P.; Murk, J.-L. Acute Influenza Virus-Associated Encephalitis and Encephalopathy in Adults: A Challenging Diagnosis. JMM Case Rep. 2016, 3, e005076. [Google Scholar] [CrossRef]
- Roubidoux, E.K.; Schultz-Cherry, S. Animal Models Utilized for the Development of Influenza Virus Vaccines. Vaccines 2021, 9, 787. [Google Scholar] [CrossRef]
- Fukao, K.; Ando, Y.; Noshi, T.; Kitano, M.; Noda, T.; Kawai, M.; Yoshida, R.; Sato, A.; Shishido, T.; Naito, A. Baloxavir Marboxil, a Novel Cap-Dependent Endonuclease Inhibitor Potently Suppresses Influenza Virus Replication and Represents Therapeutic Effects in Both Immunocompetent and Immunocompromised Mouse Models. PLoS ONE 2019, 14, e0217307. [Google Scholar] [CrossRef]
- Honce, R.; Jones, J.; Meliopoulos, V.A.; Livingston, B.; Sharp, B.; Estrada, L.D.; Wang, L.; Caulfield, W.; Freeman, B.; Govorkova, E.; et al. Efficacy of Oseltamivir Treatment in Influenza Virus-Infected Obese Mice. mBio 2023, 14, e00887-23. [Google Scholar] [CrossRef] [PubMed]
- Ison, M.G.; Mishin, V.P.; Braciale, T.J.; Hayden, F.G.; Gubareva, L.V. Comparative Activities of Oseltamivir and A-322278 in Immunocompetent and Immunocompromised Murine Models of Influenza Virus Infection. J. Infect. Dis. 2006, 193, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Lowen, A.C.; Mubareka, S.; Tumpey, T.M.; García-Sastre, A.; Palese, P. The Guinea Pig as a Transmission Model for Human Influenza Viruses. Proc. Natl. Acad. Sci. USA 2006, 103, 9988–9992. [Google Scholar] [CrossRef] [PubMed]
- Schulman, J.L.; Kilbourne, E.D. Experimental transmission of influenza virus infection in mice. II. Some factors affecting the incidence of transmitted infection. J. Exp. Med. 1963, 118, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Fong Legaspi, S.L.; Schwartzman, L.M.; Gygli, S.M.; Sheng, Z.-M.; Freeman, A.D.; Matthews, L.M.; Xiao, Y.; Ramuta, M.D.; Batchenkova, N.A.; et al. An Inactivated Multivalent Influenza A Virus Vaccine Is Broadly Protective in Mice and Ferrets. Sci. Transl. Med. 2022, 14, eabo2167. [Google Scholar] [CrossRef]
- Kinoshita, T.; Watanabe, K.; Sakurai, Y.; Nishi, K.; Yoshikawa, R.; Yasuda, J. Co-Infection of SARS-CoV-2 and Influenza Virus Causes More Severe and Prolonged Pneumonia in Hamsters. Sci. Rep. 2021, 11, 21259. [Google Scholar] [CrossRef]
- Buhnerkempe, M.G.; Gostic, K.; Park, M.; Ahsan, P.; Belser, J.A.; Lloyd-Smith, J.O. Mapping Influenza Transmission in the Ferret Model to Transmission in Humans. eLife 2015, 4, e07969. [Google Scholar] [CrossRef]
- Herlocher, M.L.; Elias, S.; Truscon, R.; Harrison, S.; Mindell, D.; Simon, C.; Monto, A.S. Ferrets as a Transmission Model for Influenza: Sequence Changes in HA1 of Type A (H3N2) Virus. J. Infect. Dis. 2001, 184, 542–546. [Google Scholar] [CrossRef]
- Weinfurter, J.T.; Brunner, K.; Capuano, S.V.; Li, C.; Broman, K.W.; Kawaoka, Y.; Friedrich, T.C. Cross-Reactive T Cells Are Involved in Rapid Clearance of 2009 Pandemic H1N1 Influenza Virus in Nonhuman Primates. PLoS Pathog. 2011, 7, e1002381. [Google Scholar] [CrossRef]
- McCarthy, K.R.; Von Holle, T.A.; Sutherland, L.L.; Oguin, T.H.; Sempowski, G.D.; Harrison, S.C.; Moody, M.A. Differential Immune Imprinting by Influenza Virus Vaccination and Infection in Nonhuman Primates. Proc. Natl. Acad. Sci. USA 2021, 118, e2026752118. [Google Scholar] [CrossRef]
- Carroll, T.D.; Matzinger, S.R.; Barry, P.A.; McChesney, M.B.; Fairman, J.; Miller, C.J. Efficacy of Influenza Vaccination of Elderly Rhesus Macaques Is Dramatically Improved by Addition of a Cationic Lipid/DNA Adjuvant. J. Infect. Dis. 2014, 209, 24–33. [Google Scholar] [CrossRef]
- Rimmelzwaan, G.F.; Baars, M.; Van Beek, R.; Van Amerongen, G.; Lövgren-Bengtsson, K.; Claas, E.C.; Osterhaus, A.D. Induction of Protective Immunity against Influenza Virus in a Macaque Model: Comparison of Conventional and Iscom Vaccines. J. Gen. Virol. 1997, 78 Pt 4, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.; Andrewes, C.H.; Laidlaw, P.P. A virus obtained from influenza patients. Lancet 1933, 222, 66–68. [Google Scholar] [CrossRef]
- Maher, J.A.; DeStefano, J. The Ferret: An Animal Model to Study Influenza Virus. Lab Anim. 2004, 33, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Baz, M.; Paskel, M.; Matsuoka, Y.; Zengel, J.; Cheng, X.; Jin, H.; Subbarao, K. Replication and Immunogenicity of Swine, Equine, and Avian H3 Subtype Influenza Viruses in Mice and Ferrets. J. Virol. 2013, 87, 6901–6910. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Eckert, A.M.; Tumpey, T.M.; Maines, T.R. Complexities in Ferret Influenza Virus Pathogenesis and Transmission Models. Microbiol. Mol. Biol. Rev. 2016, 80, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Andrewes, C.H.; Glover, R.E. Spread of Infection from the Respiratory Tract of the Ferret. I. Transmission of Influenza A Virus. Br. J. Exp. Pathol. 1941, 22, 91–97. [Google Scholar]
- Jayaraman, A.; Chandrasekaran, A.; Viswanathan, K.; Raman, R.; Fox, J.G.; Sasisekharan, R. Decoding the Distribution of Glycan Receptors for Human-Adapted Influenza A Viruses in Ferret Respiratory Tract. PLoS ONE 2012, 7, e27517. [Google Scholar] [CrossRef]
- Jia, N.; Barclay, W.S.; Roberts, K.; Yen, H.-L.; Chan, R.W.Y.; Lam, A.K.Y.; Air, G.; Peiris, J.S.M.; Dell, A.; Nicholls, J.M.; et al. Glycomic Characterization of Respiratory Tract Tissues of Ferrets: Implications for Its Use in Influenza Virus Infection Studies. J. Biol. Chem. 2014, 289, 28489–28504. [Google Scholar] [CrossRef]
- Bodewes, R.; Kreijtz, J.H.C.M.; Van Amerongen, G.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Rimmelzwaan, G.F.; Kuiken, T. Pathogenesis of Influenza A/H5N1 Virus Infection in Ferrets Differs between Intranasal and Intratracheal Routes of Inoculation. Am. J. Pathol. 2011, 179, 30–36. [Google Scholar] [CrossRef]
- Van den Brand, J.M.A.; Stittelaar, K.J.; Leijten, L.M.E.; Van Amerongen, G.; Simon, J.H.; Osterhaus, A.D.M.E.; Kuiken, T. Modification of the Ferret Model for Pneumonia from Seasonal Human Influenza A Virus Infection. Vet. Pathol. 2012, 49, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Gary, J.M.; Ritter, J.M.; Sun, X.; Maines, T.R.; Belser, J.A. Influenza A Virus Infection and Pathology in Nasal and Periocular Tissues after Ocular Inoculation in Ferrets. Vet. Pathol. 2022, 59, 1056–1061. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.; Prather, K.A.; Sznitman, J.; Jimenez, J.L.; Lakdawala, S.S.; Tufekci, Z.; Marr, L.C. Airborne Transmission of Respiratory Viruses. Science 2021, 373, eabd9149. [Google Scholar] [CrossRef] [PubMed]
- Govorkova, E.A.; Rehg, J.E.; Krauss, S.; Yen, H.-L.; Guan, Y.; Peiris, M.; Nguyen, T.D.; Hanh, T.H.; Puthavathana, P.; Long, H.T.; et al. Lethality to Ferrets of H5N1 Influenza Viruses Isolated from Humans and Poultry in 2004. J. Virol. 2005, 79, 2191–2198. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, J.; Huang, K.; Li, K.-S.; Yuen, K.-Y.; Guan, Y.; Chen, H.; Ng, W.F. Systemic Infection of Avian Influenza A Virus H5N1 Subtype in Humans. Hum. Pathol. 2009, 40, 735–739. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Delaney, C.A.; Orosz, S.E. Ferret Respiratory System: Clinical Anatomy, Physiology, and Disease. Vet. Clin. N. Am. Exot. Anim. Pract. 2011, 14, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Reeve, P.; Pibermann, M.; Gerendas, B. Studies with Some Influenza B Viruses in Cell Cultures, Hamsters and Hamster Tracheal Organ Cultures. Med. Microbiol. Immunol. 1981, 169, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.; Chanock, V.; Chanock, R.M. Temperature-Sensitive Mutants of Influenza Virus. I. Behavior in Tissue Culture and in Experimental Animals. J. Infect. Dis. 1971, 123, 145–157. [Google Scholar] [CrossRef]
- Jennings, R.; Potter, C.W.; McLaren, C. Effect of Preinfection and Preimmunization on the Serum Antibody Response to Subsequent Immunization with Heterotypic Influenza Vaccines. J. Immunol. Baltim. Md 1950 1974, 113, 1834–1843. [Google Scholar] [CrossRef]
- Potter, C.W.; Jennings, R. The Hamster as a Model System for the Study of Influenza Vaccines. Postgrad. Med. J. 1976, 52, 345–351. [Google Scholar] [CrossRef]
- Bouvier, N.M.; Lowen, A.C. Animal Models for Influenza Virus Pathogenesis and Transmission. Viruses 2010, 2, 1530–1563. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.A.; Engel, G.A.; Feeroz, M.M.; San, S.; Rompis, A.; Lee, B.P.Y.-H.; Shaw, E.; Oh, G.; Schillaci, M.A.; Grant, R.; et al. Influenza Virus Infection in Nonhuman Primates. Emerg. Infect. Dis. 2012, 18, 1672–1675. [Google Scholar] [CrossRef] [PubMed]
- Rimmelzwaan, G.F.; Baars, M.; Van Amerongen, G.; Van Beek, R.; Osterhaus, A.D. A Single Dose of an ISCOM Influenza Vaccine Induces Long-Lasting Protective Immunity against Homologous Challenge Infection but Fails to Protect Cynomolgus Macaques against Distant Drift Variants of Influenza A (H3N2) Viruses. Vaccine 2001, 20, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, S.; Iwatsuki-Horimoto, K.; Kiso, M.; Nakajima, N.; Gregg, R.W.; Katsura, H.; Tomita, Y.; Maemura, T.; Da Silva Lopes, T.J.; Watanabe, T.; et al. Pathogenesis of Influenza A(H7N9) Virus in Aged Nonhuman Primates. J. Infect. Dis. 2020, 222, 1155–1164. [Google Scholar] [CrossRef]
- Davis, A.S.; Taubenberger, J.K.; Bray, M. The Use of Nonhuman Primates in Research on Seasonal, Pandemic and Avian Influenza, 1893–2014. Antiviral Res. 2015, 117, 75–98. [Google Scholar] [CrossRef]
- Chan, M.; Tiwary, M.; Wu, H.L.; Tailor, N.; Vendramelli, R.; Audet, J.; Warner, B.M.; Tierney, K.; Albietz, A.; Truong, T.; et al. Pandemic 1918 Influenza Virus Does Not Cause Lethal Infection in Rhesus or Cynomolgus Macaques. J. Virol. 2022, 96, e00728-22. [Google Scholar] [CrossRef]
| Benefits | Drawbacks | Main Applications [Refs] | |
|---|---|---|---|
| Mouse | Low cost Many tools available Genetically malleable Rapid rate of reproduction | Require adapted strains to model seasonal flu Lack influenza restriction factor Mx1 Do not sneeze and thus cannot model airborne transmission | Vaccine and therapeutic testing [75,139,152] Pathogenesis of severe disease [77,80,140,141] |
| Hamster | Low cost Simple management Naturally susceptible to human and animal IAVs Contact transmission Similar sialic acid residue distribution to humans | Fewer reagents Lack of clinical signs like sneezing and nasal discharge Not very friendly | IAV and coronavirus co-infection [103,104,105,153] |
| Ferret | Naturally susceptible to human and animal IAVs Transmission via respiratory droplets Similar sialic acid residue distribution to humans | Fewer reagents Lack of inbred strains Increased cost | Transmission studies [113,116,154,155] Pathogenesis [115] |
| NHP | Anatomic and immunologic similarities to humans Naturally susceptible to human and animal IAVs | High cost Complicated husbandry requirements Ethical concerns Lack of availability | Immunology [123,124,125,130,132,156,157] Vaccine and therapeutic testing [131,133,158,159] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirk, N.M.; Liang, Y.; Ly, H. Comparative Pathology of Animal Models for Influenza A Virus Infection. Pathogens 2024, 13, 35. https://doi.org/10.3390/pathogens13010035
Kirk NM, Liang Y, Ly H. Comparative Pathology of Animal Models for Influenza A Virus Infection. Pathogens. 2024; 13(1):35. https://doi.org/10.3390/pathogens13010035
Chicago/Turabian StyleKirk, Natalie M., Yuying Liang, and Hinh Ly. 2024. "Comparative Pathology of Animal Models for Influenza A Virus Infection" Pathogens 13, no. 1: 35. https://doi.org/10.3390/pathogens13010035
APA StyleKirk, N. M., Liang, Y., & Ly, H. (2024). Comparative Pathology of Animal Models for Influenza A Virus Infection. Pathogens, 13(1), 35. https://doi.org/10.3390/pathogens13010035