Phylogeographic Aspects of Bat Lyssaviruses in Europe: A Review

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
| Bat Species | IUCN Status | Bat Identification | Virus Detection | European Bat Lyssaviruses | References | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EBLV-1 | EBLV-2 | BBLV | KBLV | DBLV | LLEBV | WCBV | ||||||||||||
| Spill over Infection | Host Species | Spill over Infection | Host Species | Spill over Infection | Host Species | Spill over Infection | Host Species | Spill over Infection | Host Species | Spill over Infection | Host Species | Spill over Infection | Host Species | |||||
| family Rhinolophidae | ||||||||||||||||||
| Rhinolophus ferrumequinum | NT | morphology | FAT, RT-PCR, RFFIT, mFAVNt | + | [36,37] | |||||||||||||
| family Vespertilionidae | ||||||||||||||||||
| Barbastella barbastellus | VU | morphology | mFAVNt | + | [37] | |||||||||||||
| Eptesicus isabellinus | N/A | morphology | RT-PCR | + | [38,39] | |||||||||||||
| Eptesicus serotinus | LC | cyt b | FAT, RT-PCR, RFFIT | + | + | [36,38,39,40,41,42,43,44,45,46,47,48,49] | ||||||||||||
| Myotis blythii | NT | morphology | mFAVNt | + | [37] | |||||||||||||
| Myotis brandtii | LC | nd1, cyt b | FAT, RT-PCR | + | [24,45] | |||||||||||||
| Myotis capaccinii | VU | morphology | FAT, RFFIT, RTCIT, RT-PCR | + | [35] | |||||||||||||
| Myotis dasycneme | NT | morphology | RT-PCR | + | [41,45,49] | |||||||||||||
| Myotis daubentonii | LC | cyt b | RT-PCR | + | + | [41,45,46,47,48,49] | ||||||||||||
| Myotis myotis | LC | cyt b | FAT, RT-PCR, RFFIT | + | [36,45,48] | |||||||||||||
| Myotis nattereri | LC | cyt b | FAT, RT-PCR, RFFIT | + | + | [18,45,50,51,52] | ||||||||||||
| Nyctalus noctula | LC | cyt b | FAT, RT-PCR | + | [45,48] | |||||||||||||
| Pipistrellus nathusii | LC | cyt b | FAT, RT-qPCR, RT-PCR, RTCIT | + | [45,48] | |||||||||||||
| Pipistrellus pipistrellus | LC | cyt b | FAT, RT-qPCR, RT-PCR, RTCIT | + | [45,48] | |||||||||||||
| Plecotus auritus | LC | cyt b | FAT, RT-qPCR, RT-PCR, RTCIT | + | [45,48] | |||||||||||||
| Vespertilio murinus | LC | cyt b | RT-PCR | + | [41,45] | |||||||||||||
| family Miniopteridae | ||||||||||||||||||
| Miniopterus schreibersii | NT | morphology | FAT, RT-PCR, RFFIT, mFAVNt | + | + | + | + | [36,37,53] | ||||||||||
| family Molossidae | ||||||||||||||||||
| Tadarida teniotis | LC | morphology | FAT+ | + | [36] | |||||||||||||
| family Pteropodidae | ||||||||||||||||||
| Rousettus aegyptiacus | N/A (EN?) | morphology | FAT, RT-PCR, RFFIT | + | [54] | |||||||||||||
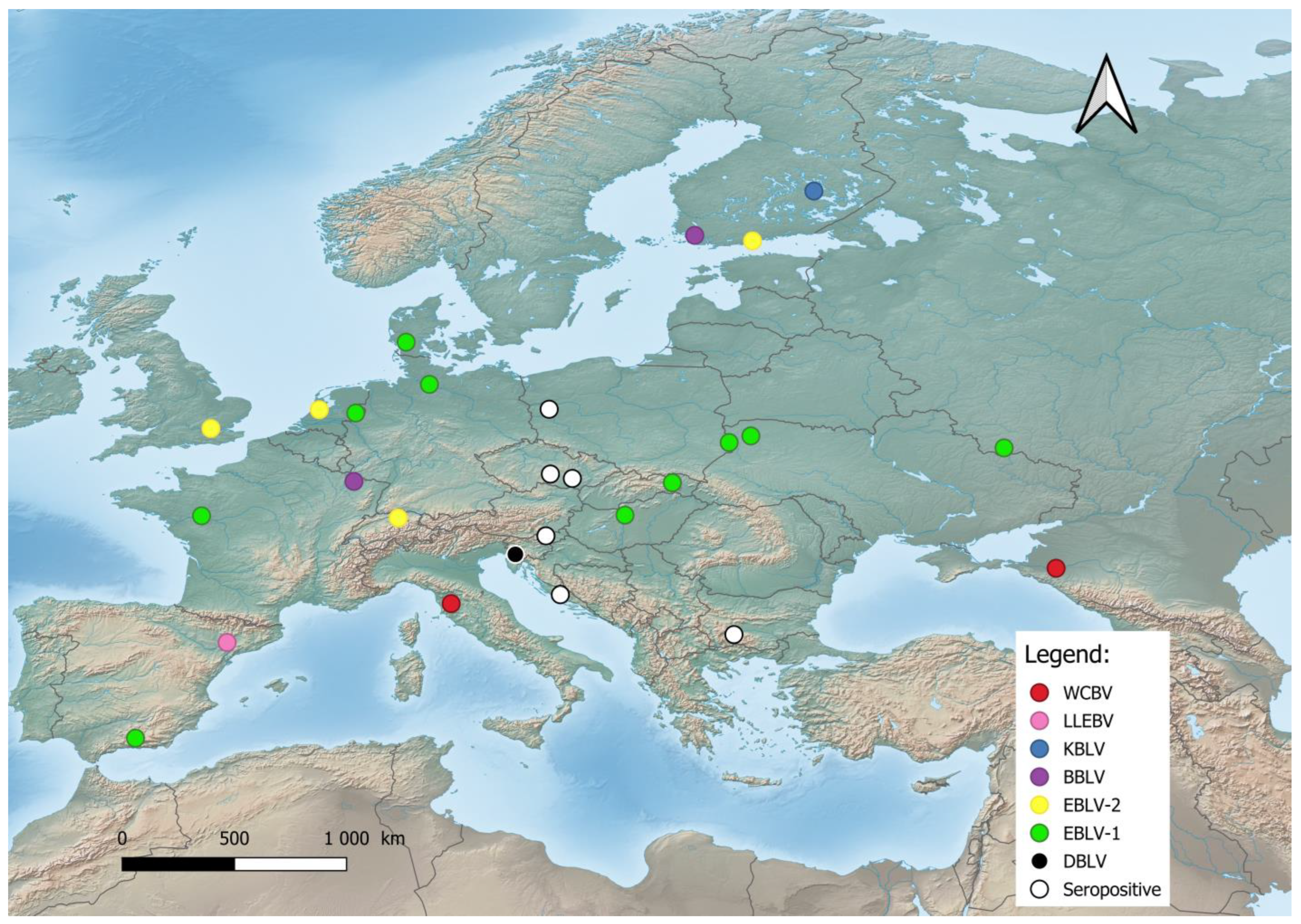
2. Origin, Evolution, and Geographic Distribution of Bat Lyssaviruses
3. Virion Structure and Genome
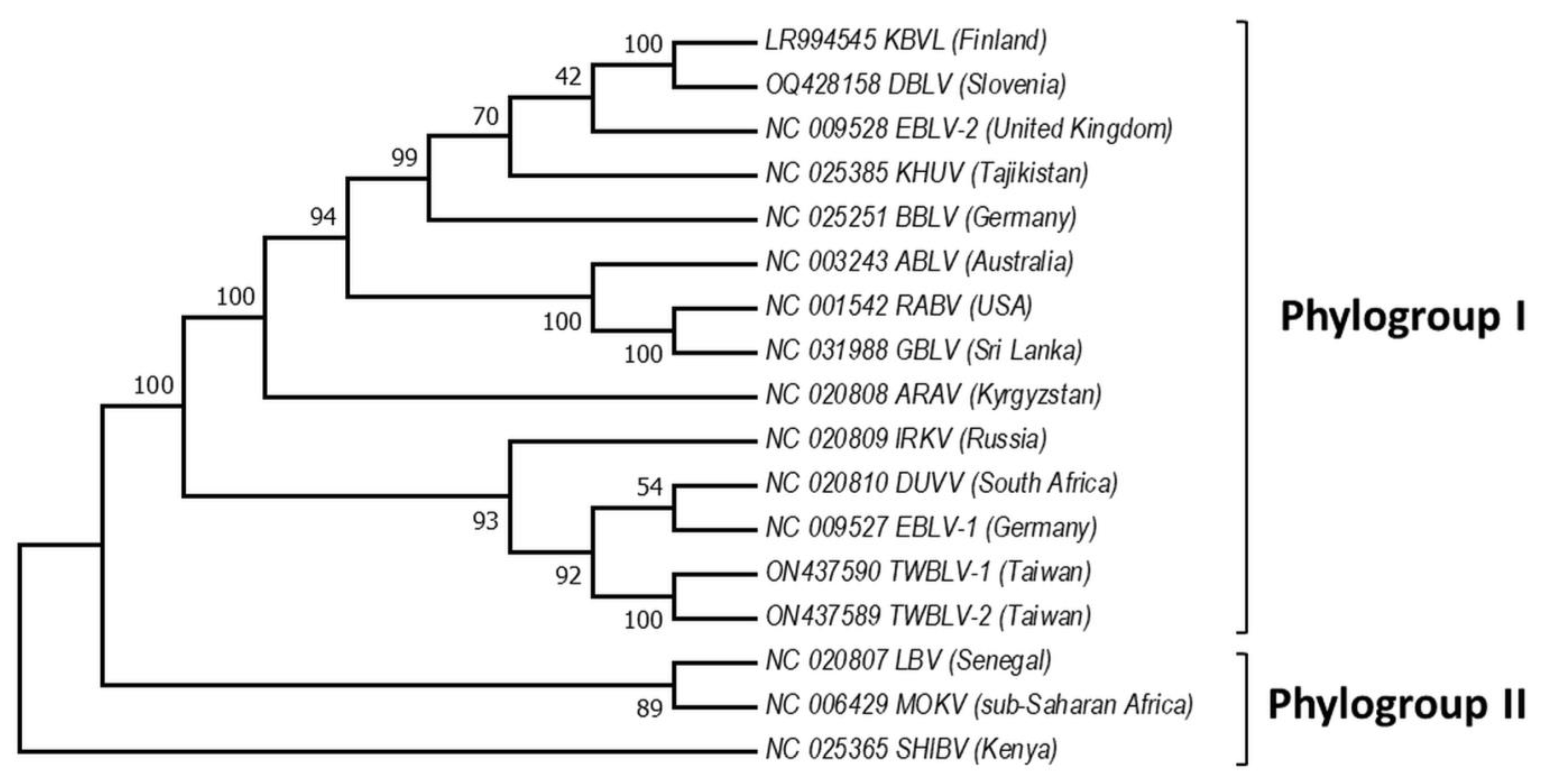
4. Phylogeny of Bat Lyssaviruses
5. Transmission Routes of Bat Lyssaviruses
5.1. Bat Intra- and Cross-Species Transmission
5.2. Other Vertebrates Cross-Species Transmission
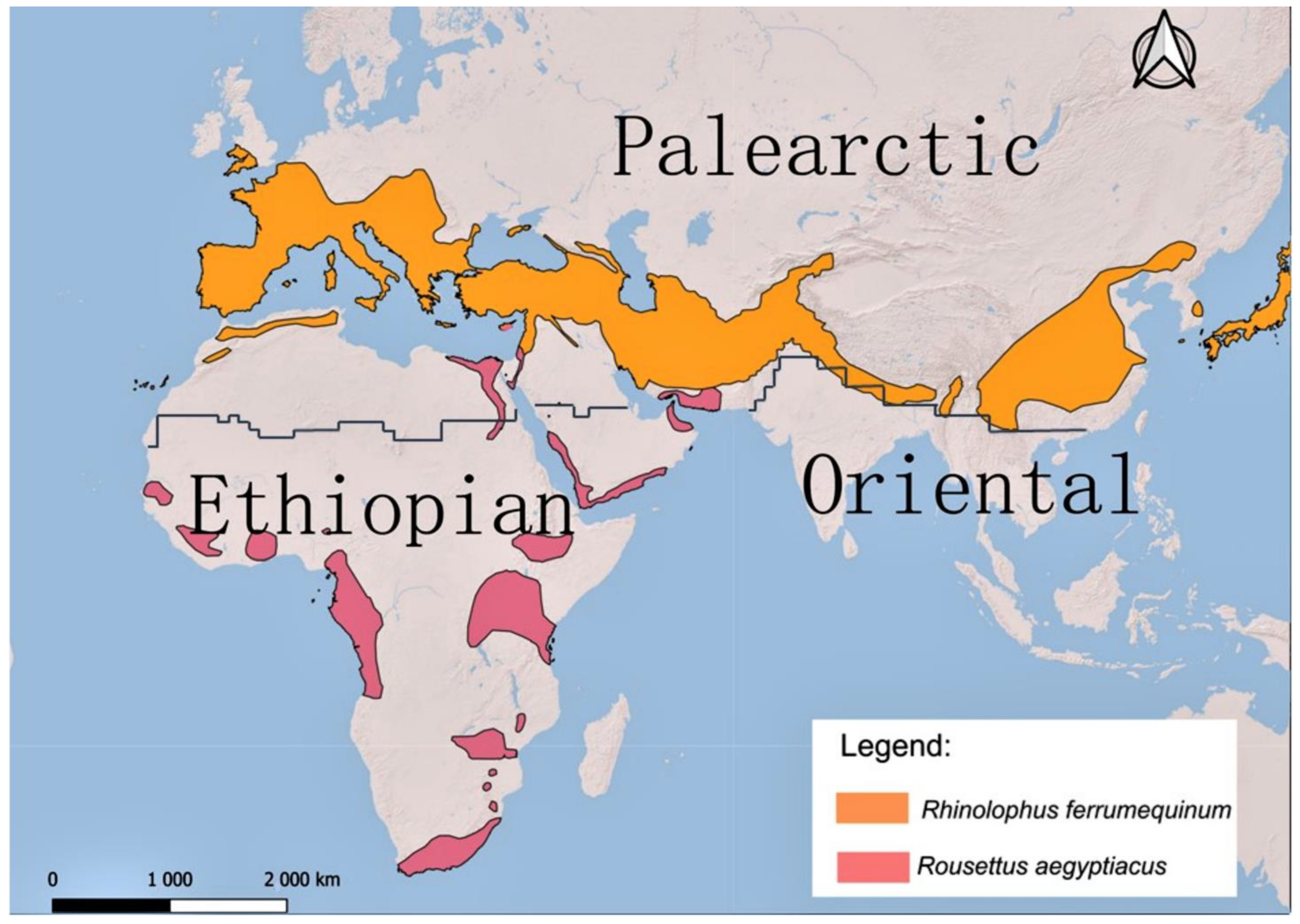
5.3. Within the Contact Zone of the Palearctic and Ethiopian Realms—Potential Scenarios
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ruedi, M.; Stadelmann, B.; Gager, Y.; Douzery, E.J.; Francis, C.M.; Lin, L.K.; Guillén-Servent, A.; Cibois, A. Molecular phylogenetic reconstructions identify East Asia as the cradle for the evolution of the cosmopolitan genus Myotis (Mammalia, Chiroptera). Mol. Phylogenet. Evol. 2013, 69, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Teeling, E.C.; Vernes, S.C.; Dávalos, L.M.; Ray, D.A.; Gilbert, M.T.P.; Myers, E. Bat1K Consortium. Bat Biology, Genomes, and the Bat1K Project: To Generate Chromosome-Level Genomes for All Living Bat Species. Annu. Rev. Anim. Biosci 2018, 6, 23–46. [Google Scholar] [CrossRef] [PubMed]
- Teeling, E.C.; Jones, G.; Rossiter, S.J. Phylogeny, genes, and hearing: Implications for the evolution of echolocation in bats. In Bat Bioacoustics; Springer: New York, NY, USA, 2016; pp. 25–54. [Google Scholar]
- Simmons, N.B.; Cirranello, A.L. Bat Species of the World: A Taxonomic and Geographic Database, Version 1.3. 2023. 2023. Available online: https://batnames.org/ (accessed on 8 August 2023).
- Wackermannová, M.; Pinc, L.; Jebavý, L. Olfactory sensitivity in mammalian species. Physiol. Res. 2016, 65, 369. [Google Scholar] [CrossRef]
- Wilkinson, G.S.; Adams, D.M.; Haghani, A.; Lu, A.T.; Zoller, J.; Breeze, C.E.; Arnold, B.D.; Ball, H.C.; Carter, G.G.; Cooper, L.N.; et al. Genome Methylation Predicts Age and Longevity of Bats. Nat. Commun. 2021, 12, 1615. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.-Y.; Liang, L.; Zhu, Z.-H.; Zhou, W.-P.; Irwin, D.M.; Zhang, Y.-P.; Hillis, D.M. Adaptive evolution of energy metabolism genes and the origin of flight in bats. PNAS 2010, 107, 8666–8671. [Google Scholar] [CrossRef] [PubMed]
- Subudhi, S.; Rapin, N.; Misra, V. Immune System Modulation and Viral Persistence in Bats: Understanding Viral Spillover. Viruses 2019, 11, 192. [Google Scholar] [CrossRef]
- Banerjee, A.; Baker, M.L.; Kulcsar, K.; Misra, V.; Plowright, R.; Mossman, K. Novel Insights into Immune Systems of Bats. Front. Immunol. 2020, 11, 26. [Google Scholar] [CrossRef]
- Begeman, L.; Suu-Ire, R.; Banyard, A.C.; Drosten, C.; Eggerbauer, E.; Freuling, C.M.; Gibson, l.; Goharriz, H.; Horton, D.L.; Jennings, D.; et al. Experimental Lagos bat virus infection in straw-colored fruit bats: A suitable model for bat rabies in a natural reservoir species. PLoS Negl. Trop. Dis. 2020, 14, e0008898. [Google Scholar] [CrossRef] [PubMed]
- Irving, A.T.; Ahn, M.; Goh, G.; Anderson, D.E.; Wang, L.F. Lessons from the host defences of bats, a unique viral reservoir. Nature 2021, 589, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Brook, C.E.; Dobson, A.P. Bats as ‘special’ reservoirs for emerging zoonotic pathogens. Trends Microbiol. 2015, 23, 172–180. [Google Scholar] [CrossRef]
- Hayman, D.T.S. Bats as viral reservoirs. Annu. Rev. Virol. 2016, 3, 77–99. [Google Scholar] [CrossRef]
- Latinne, A.; Hu, B.; Olival, K.J.; Zhu, G.; Zhang, L.; Li, H.; Chmura, A.A.; Field, H.E.; Zambrana-Torrelio, C.; Epstein, J.H.; et al. Origin and cross-species transmission of bat coronaviruses in China. Nat. Commun. 2020, 11, 4235. [Google Scholar] [CrossRef] [PubMed]
- EUROBATS. Action Plan for the Conservation of Bat Species in the European Union 2016–2021; Inf.EUROBATS.AC21.5; EUROBATS: Bonn, Germany, 2006. [Google Scholar]
- Teeling, E.C.; Springer, M.S.; Madsen, O.; Bates, P.; O’Brien, S.J.; Murphy, W.J. A molecular phylogeny for bats illuminates biogeography and the fossil record. Science 2005, 307, 580–584. [Google Scholar] [CrossRef]
- Kohl, C.; Kurth, A. European bats as carriers of viruses with zoonotic potential. Viruses 2014, 6, 3110–3128. [Google Scholar] [CrossRef]
- Smreczak, M.; Orłowska, A.; Marzec, A.; Trębas, P.; Müller, T.; Freuling, C.M.; Żmudziński, J.F. Bokeloh bat lyssavirus isolation in a Natterer’s bat, Poland. Zoonoses Public Health 2018, 65, 1015–1019. [Google Scholar] [CrossRef]
- Vos, A.; Kaipf, I.; Denzinger, A.; Fooks, A.R.; Johnson, N.; Müller, T. European bat lyssaviruses—An ecological enigma. Acta Chiropt. 2007, 9, 283–296. [Google Scholar] [CrossRef]
- Banyard, A.C.; Hayman, D.; Johnson, N.; McElhinney, L.; Fooks, A.R. Bats and lyssaviruses. Adv. Virus Res. 2011, 79, 239–289. [Google Scholar] [CrossRef]
- Banyard, A.C.; Davis, A.; Gilbert, A.; Markotter, W. Bat Rabies. In Rabies: Scientific Basis of the Disease and Its Management, 4th ed.; Fooks, A.R., Jackson, A.C., Eds.; Chapter 7; Elsevier: Amsterdam, The Netherlands, 2020; pp. 231–276. ISBN 978-0-12-818705-0. [Google Scholar]
- Afonso, C.L.; Amarasinghe, G.K.; Bányai, K.; Bào, Y.; Basler, C.F.; Bavari, S.; Bejerman, N.; Blasdell, K.R.; Briand, F.X.; Briese, T.; et al. Taxonomy of the order Mononegavirales: Update 2016. Arch Virol. 2016, 161, 2351–2360. [Google Scholar] [CrossRef]
- Nokireki, T.; Tammiranta, N.; Kokkonen, U.M.; Kantala, T.; Gadd, T. Tentative novel lyssavirus in a bat in Finland. Transbound. Emerg. Dis. J. 2018, 65, 593–596. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, J.H.; Adkins, S.; Agwanda, B.R.; Al Kubrusli, R.; Alkhovsky, S.V.; Amarasinghe, G.K.; Avšič-Županc, T.; Ayllón, M.A.; Bahl, J.; Balkema-Buschmann, A.; et al. Taxonomic update of phylum Negarnaviricota (Riboviria: Orthornavirae), including the large orders Bunyavirales and Mononegavirales. Arch Virol. 2021, 166, 3513–3566. [Google Scholar] [CrossRef] [PubMed]
- Coertse, J.; Markotter, W.; Le Roux, K.; Stewart, D.; Sabeta, C.T.; Nel, L.H. New isolations of the rabies related Mokola virus from South Africa. BMC Vet. Res. 2017, 13, 37. [Google Scholar] [CrossRef]
- Klein, A.; Calvelage, S.; Schlottau, K.; Hoffmann, B.; Eggerbauer, E.; Müller, T.; Freuling, C.M. Retrospective Enhanced Bat Lyssavirus Surveillance in Germany between 2018–2020. Viruses 2021, 13, 1538. [Google Scholar] [CrossRef] [PubMed]
- Marston, D.A.; Horton, D.L.; Ngeleja, C.; Hampson, K.; McElhinney, L.M.; Banyard, A.C.; Haydon, D.; Cleaveland, S.; Rupprecht, C.E.; Bigambo, M.; et al. Ikoma lyssavirus, highly divergent novel lyssavirus in an African civet. Emerg. Infect. Dis. 2012, 18, 664–667. [Google Scholar] [CrossRef]
- Markotter, W.; Kgaladi, J.; Nel, L.H.; Marston, D.; Wright, N.; Coertse, J.; Müller, T.F.; Sabeta, C.T.; Fooks, A.R.; Freuling, C.M. Diversity and Epidemiology of Mokola Virus. PLoS Negl. Trop. Dis. 2013, 7, e2511. [Google Scholar]
- Sabeta, C.T.; Markotter, W.; Mohale, D.K.; Shumba, W.; Wandeler, A.I.; Nel, L.H. Mokola virus in domestic mammals, South Africa. Emerg. Infect. Dis. 2007, 13, 1371–1373. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, C.; Kuzmin, I.; Meslin, F. Lyssaviruses and rabies: Current conundrums, concerns, contradictions and controversies. F1000Research 2017, 6, 184. [Google Scholar] [CrossRef]
- Hu, S.; Hsu, C.; Lee, M.; Tu, Y.; Chang, J.; Wu, C.; Lee, S.-H.; Ting, L.-J.; Tsai, K.-R.; Cheng, M.-C.; et al. Lyssavirus in Japanese Pipistrelle, Taiwan. Emerg. Infect. Dis. 2018, 24, 782–785. [Google Scholar] [CrossRef]
- Calvelage, S.; Tammiranta, N.; Nokireki, T.; Gadd, T.; Eggerbauer, E.; Zaeck, L.M.; Potratz, M.; Wylezich, C.; Höper, D.; Müller, T.; et al. Genetic and Antigenetic Characterization of the Novel Kotalahti Bat Lyssavirus (KBLV). Viruses 2021, 13, 69. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.-C.; Hsu, C.-L.; Lee, F.; Tu, Y.-C.; Chen, Y.-W.; Chang, J.-C.; Hsu, W.-C. Novel Bat Lyssaviruses Identified by Nationwide Passive Surveillance in Taiwan, 2018–2021. Viruses 2022, 14, 1562. [Google Scholar] [CrossRef]
- Coertse, J.; Grobler, C.S.; Sabeta, C.T.; Seamark, E.C.J.; Kearney, T.; Paweska, J.T.; Markotter, W. Lyssaviruses in Insectivorous Bats, South Africa, 2003-2018. Emerg. Infect. Dis. 2020, 26, 3056–3060. [Google Scholar] [CrossRef] [PubMed]
- Černe, D.; Hostnik, P.; Toplak, I.; Presetnik, P.; Maurer-Wernig, J.; Kuhar, U. Discovery of a novel bat lyssavirus in a Long-fingered bat (Myotis capaccinii) from Slovenia. PLoS Negl Trop Dis. 2023, 17, e0011420. [Google Scholar] [CrossRef]
- Serra-Cobo, J.; Amengual, B.; Abellan, C.; Bourhy, H. European bat Lyssavirus infection in Spanish bat populations. Emerg. Infect. Dis. 2002, 8, 413–420. [Google Scholar] [CrossRef]
- Picard-Meyer, E.; Dubourg-Savage, M.J.; Arthur, L.; Barataud, M.; Bécu, D.; Bracco, S.; Borel, C.; Larcher, G.; Meme-Lafond, B.; Moinet, M.; et al. Active surveillance of bat rabies in France: A 5-year study (2004–2009). Vet Microbiol. 2011, 151, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Schatz, J.; Fooks, A.R.; McElhinney, L.; Horton, D.; Echevarria, J.; Vázquez-Moron, S.; Kooi, E.A.; Rasmussen, T.B.; Müller, T.; Freuling, C.M. Bat rabies surveillance in Europe. Zoonoses Public Health 2012, 60, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Mingo-Casas, P.; Sandonis, V.; Obón, E.; Berciano, J.M.; Vázquez-Morón, S.; Juste, J.; Echevarria, J.E. First cases of European bat lyssavirus type 1 in Iberian serotine bats: Implications for the molecular epidemiology of bat rabies in Europe. PLoS Negl Trop Dis. 2018, 12, e0006290. [Google Scholar] [CrossRef] [PubMed]
- Echevarria, J.E.; Avellon, A.; Juste, J.; Vera, M.; Ibayez, C. Screening of active lyssavirus infection in wild bat populations by viral RNA on oropharyngeal swabs. J. Clin. Microbiol. 2001, 39, 3678–3683. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.L.; Holmes, E.C.; Larrous, F.; Van der Poel, W.H.; Tjørnehøj, K.; Alonso, W.J.; Bourhy, H. Phylogeography, population dynamics, and molecular evolution of European bat lyssaviruses. Virol. J. 2005, 79, 10487–10497. [Google Scholar] [CrossRef]
- Dacheux, L.; Larrous, F.; Mailles, A.; Boisseleau, D.; Delmas, O.; Biron, C.; Bouchier, C.; Capek, I.; Muller, M.; Ilari, F.; et al. European Bat Lyssavirus Transmission among Cats, Europe. Emerg. Infect. Dis. 2009, 15, 280–284. [Google Scholar] [CrossRef]
- Muller, T.; Cox, J.; Peter, W.; Schafer, R.; Johnson, N.; McElhinney, L.M.; Geue, J.L.; Tjornehoj, K.; Fooks, A.R. Spill-over of European bat lyssavirus type 1 into a stone marten (Martes foina) in Germany. J. Vet. Med. Ser. B 2004, 51, 49–54. [Google Scholar] [CrossRef]
- Tjørnehøj, K.; Fooks, A.R.; Agerholm, J.S.; Rønsholt, L. Natural and experimental infection of sheep with European bat lyssavirus type-1 of Danish bat origin. J. Comp. Pathol. 2006, 134, 190–201. [Google Scholar] [CrossRef]
- Schatz, J.; Freuling, C.M.; Auer, E.; Goharriz, H.; Harbusch, C.; Johnson, N.; Kaipf, I.; Mettenleiter, T.C.; Mühldorfer, K.; Mühle, R.-U.; et al. Enhanced Passive Bat Rabies Surveillance in Indigenous Bat Species from Germany—A Retrospective Study. PLoS Negl. Trop Dis. 2014, 8, e2835. [Google Scholar] [CrossRef]
- Megali, A.; Yannic, G.; Zahno, M.L.; Brügger, D.; Bertoni, G.; Christe, P.; Zanoni, R. Surveillance for European bat lyssavirus in Swiss bats. Arch Virol. 2010, 155, 1655–1662. [Google Scholar] [CrossRef] [PubMed]
- Fooks, A.R.; McElhinney, L.M.; Pounder, D.J.; Finnegan, C.J.; Mansfield, K.; Johnson, N.; Brookes, S.M.; Parsons, G.; White, K.; McIntyre, P.G.; et al. Case Report: Isolation of a European Bat Lyssavirus Type 2a From a Fatal Human Case of Rabies Encephalitis. J. Med. Virol. 2003, 71, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.; Johnson, N.; Freuling, C.M.; Fooks, A.R.; Selhorst, T.; Vos, A. Epidemiology of bat rabies in Germany. Arch Virol. 2007, 152, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Amengual, B.; Whitby, J.E.; King, A.; Cobo, J.S.; Bourhy, H. Evolution of European bat lyssaviruses. J. Gen. Virol. 1997, 78, 2319–2328. [Google Scholar] [CrossRef]
- Freuling, C.M.; Beer, M.; Conraths, F.J.; Finke, S.; Hoffmann, B.; Keller, B.; Kliemt, J.; Mettenleiter, T.C.; Mühlbach, E.; Teifke, J.P.; et al. Novel lyssavirus in natterer’s bat, Germany. Emerg. Infect. Dis. 2011, 17, 1519–1522. [Google Scholar] [CrossRef]
- Picard-Meyer, E.; Servat, A.; Robardet, E.; Moinet, M.; Borel, C.; Cliquet, F. Isolation of Bokeloh bat lyssavirus in Myotis nattereri in France. Arch. Virol. 2013, 158, 2333–2340. [Google Scholar] [CrossRef] [PubMed]
- Puechmaille, S.J.; Allegrini, B.; Boston, E.S.; Dubourg-Savage, M.-J.; Evin, A.; Knochel, A.; Le Bris, Y.; Lecoq, V.; Lemaire, M.; Rist, D.; et al. Genetic analyses reveal further cryptic lineages within the Myotis nattereri species complex. Mamm. Biol. 2012, 77, 224–228. [Google Scholar] [CrossRef]
- Coxon, C.; McElhinney, L.; Pacey, A.; Gauntlett, F.; Holland, S. Preliminary Outbreak Assessment: Rabies in a Cat in Italy. Department for Environment, Food and Rural Affairs, Animal and Plant Health Agency, Advice Services—International Disease Monitoring. 2020. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/897070/rabies-cat-italy-poa.pdf (accessed on 8 August 2023).
- Van Der Poel, W.H.M.; Van Der Heide, R.; Van Amerongen, G.; Van Keulen, L.J.M.; Wellenberg, G.J.; Bourhy, H.; Schaftenaar, W.; Groen, J.; Osterhaus, A.D.M.E. Characterisation of a recently isolated lyssavirus in frugivorous zoo bats. Arch. Virol. 2000, 145, 1919–1931. [Google Scholar] [CrossRef]
- Nel, L.H.; Rupprecht, C.E. Emergence of lyssaviruses in the Old World: The case of Africa. Curr. Top. Microbiol. Immunol. 2007, 315, 161–193. [Google Scholar] [CrossRef]
- Hayman, D.T.S.; Fooks, A.R.; Marston, D.A.; Garcia-R, J.C. The Global Phylogeography of Lyssaviruses—Challenging the ‘Out of Africa’ Hypothesis. PLoS Negl. Trop. Dis. 2016, 10, e0005266. [Google Scholar] [CrossRef] [PubMed]
- Longdon, B.; Murray, G.G.; Palmer, W.J.; Day, J.P.; Parker, D.J.; Welch, J.J.; Obbard, D.J.; Jiggins, F.M. The evolution, diversity, and host associations of rhabdoviruses. Virus Evol. 2015, 1, vev014. [Google Scholar] [CrossRef]
- Li, C.X.; Shi, M.; Tian, J.H.; Lin, X.D.; Kang, Y.J.; Chen, L.; Qin, X.C.; Xu, J.; Holmes, E.C.; Zhang, Y.Z. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. Elife 2015, 29, e05378. [Google Scholar] [CrossRef]
- Walker, P.J.; Firth, C.; Widen, S.G.; Blasdell, K.R.; Guzman, H.; Wood, T.G.; Paradkar, P.N.; Holmes, E.; Tesh, R.B.; Vasilakis, N. Evolution of genome size and complexity in the Rhabdoviridae. PLoS Pathog. 2015, 11, e1004664. [Google Scholar] [CrossRef]
- Velasco-Villa, A.; Mauldin, M.R.; Shi, M.; Escobar, L.E.; Gallardo-Romero, N.F.; Damon, I.; Olson, V.A.; Streicker, D.G.; Emerson, G. The history of rabies in the Western Hemisphere. Antivir. Res. 2017, 146, 221–232. [Google Scholar] [CrossRef]
- Caraballo, D.A.; Lema, C.; Novaro, L.; Gury-Dohmen, F.; Russo, S.; Beltrán, F.J.; Palacios, G.; Cisterna, D.M. A Novel Terrestrial Rabies Virus Lineage Occurring in South America: Origin, Diversification, and Evidence of Contact between Wild and Domestic Cycles. Viruses 2021, 13, 2484. [Google Scholar] [CrossRef]
- Singh, R.; Singh, K.P.; Cherian, S.; Saminathan, M.; Kapoor, S.; Manjunatha Reddy, G.B.; Panda, S.; Dhama, K. Rabies–epidemiology, pathogenesis, public health concerns and advances in diagnosis and control: A comprehensive review. Vet. Q. 2017, 37, 212–251. [Google Scholar] [CrossRef]
- Potratz, M.; Zaeck, L.M.; Weigel, C.; Klein, A.; Freuling, C.M.; Müller, T.; Finke, S. Neuroglia infection by rabies virus after anterograde virus spread in peripheral neurons. ANC 2020, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Baghi, H.B.; Rupprecht, C. Notes on three periods of rabies focus in the Middle East: From progress during the cradle of civilization to neglected current history. Zoonoses Public Health 2021, 68, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Badrane, H.; Tordo, N. Host switching in Lyssavirus history from the chiroptera to the carnivora orders. J. Virol. 2001, 75, 8096–8104. [Google Scholar] [CrossRef]
- Rupprecht, C.E.; Turmelle, A.; Kuzmin, I.V. A perspective on lyssavirus emergence and perpetuation. COVIRO 2011, 1, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Badrane, H.; Bahloul, C.; Perrin, P.; Tordo, N. Evidence of two Lyssavirus phylogroups with distinct pathogenicity and immunogenicity. Virol. J. 2001, 75, 3268–3276. [Google Scholar] [CrossRef] [PubMed]
- Fooks, A.; Brookes, S.; Johnson, N.; McElhinney, L.; Hutson, A. European bat lyssaviruses: An emerging zoonosis. Epidemiol. Infect. 2003, 131, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- McElhinney, L.M.; Marston, D.A.; Wise, E.L.; Freuling, C.M.; Bourhy, H.; Zanoni, R.; Moldal, T.; Kooi, E.A.; Neubauer-Juric, A.; Nokireki, T.; et al. Molecular Epidemiology and Evolution of European Bat Lyssavirus 2. Int. J. Mol. Sci. 2018, 19, 156. [Google Scholar] [CrossRef] [PubMed]
- Šimić, I.; Lojkić, I.; Krešić, N.; Cliquet, F.; Picard-Meyer, E.; Wasniewski, M.; Ćukušić, A.; Zrnčić, V.; Bedeković, T. Molecular and serological survey of lyssaviruses in Croatian bat populations. BMC Vet. Res. 2018, 14, 274. [Google Scholar] [CrossRef]
- Seidlova, V.; Zukal, J.; Brichta, J.; Anisimov, N.; Apoznański, G.; Bandouchova, H.; Bartonička, T.; Berková, H.; Botvinkin, A.D.; Heger, T.; et al. Active surveillance for antibodies confirms circulation of lyssaviruses in Palearctic bats. BMC Vet. Res. 2020, 16, 482. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Hughes, G.J.; Botvinkin, A.D.; Orciari, L.A.; Rupprecht, C.E. Phylogenetic relationships of Irkut and West Caucasian bat viruses within the Lyssavirus genus and suggested quantitative criteria based on the N gene sequence for lyssavirus genotype definition. Virus Res. 2005, 111, 28–43. [Google Scholar] [CrossRef]
- Ceballos, N.; Morón, S.; Berciano, J.M.; Nicolás, O.; López, C.; Juste, J.; Nevado, C.; Setién, A.A.; Echevarría, J.E. Novel Lyssavirus in Bat, Spain. Emerg Infect Dis. 2013, 19, 793–795. [Google Scholar] [CrossRef]
- Jakava-Viljanen, M.; Nokireki, T.; Sironen, T.; Vapalahti, O.; Sihvonen, L.; Liisa, S.; Huovilainen, A. Evolutionary trends of European bat lyssavirus type 2 including genetic characterization of Finnish strains of human and bat origin 24 years apart. Arch. Virol. 2015, 160, 1489–1498. [Google Scholar] [CrossRef]
- Harris, S.L.; Aegerter, J.N.; Brookes, S.M.; McElhinney, L.M.; Jones, G.; Smith, G.C.; Fooks, A.R. Targeted surveillance for European bat lyssaviruses in English bats (2003-06). J. Wildl. Dis. 2009, 45, 1030–1041. [Google Scholar] [CrossRef]
- Dietzgen, R.G.; Kondo, H.; Goodin, M.M.; Kurath, G.; Vasilakis, N. The family Rhabdoviridae: Mono- and bipartite negative-sense RNA viruses with diverse genome organization and common evolutionary origins. Virus Res. 2017, 227, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Lelli, D.; Prosperi, A.; Moreno, A.; Chiapponi, C.; Gibellini, A.M.; De Benedictis, P.; Leopardi, S.; Sozzi, E.; Lavazza, A. Isolation of a novel Rhabdovirus from an insectivorous bat (Pipistrellus kuhlii) in Italy. Virol. J. 2018, 15, 1–7. [Google Scholar] [CrossRef]
- Walker, P.J.; Dietzgen, R.G.; Joubert, D.A.; Blasdell, K.R. Rhabdovirus accessory genes. Virus Res. 2011, 162, 110–125. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, I.V.; Wu, X.; Tordo, N.; Rupprecht, C.E. Complete genomes of Aravan, Khujand, Irkut and West Caucasian bat viruses, with special attention to the polymerase gene and non-coding regions. Virus Res. 2008, 136, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, I.V.; Orciari, L.A.; Arai, Y.T.; Smith, J.S.; Hanlon, C.A.; Kameoka, Y.; Rupprecht, C.E. Bat lyssaviruses (Aravan and Khujand) from Central Asia: Phylogenetic relationships according to N, P and G gene sequences. Virus Res. 2003, 97, 65–79. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Novella, I.S.; Dietzgen, R.G.; Padhi, A.; Rupprecht, C.E. The rhabdoviruses: Biodiversity, phylogenetics, and evolution. Infect Genet Evol. 2009, 9, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Delmas, O.; Holmes, E.C.; Talbi, C.; Larrous, F.; Dacheux, L.; Bouchier, C.; Bourhy, H. Genomic diversity and evolution of the lyssaviruses. PLoS ONE 2008, 3, e2057. [Google Scholar] [CrossRef]
- Fooks, A.R.; Cliquet, F.; Finke, S.; Freuling, C.; Hemachudha, T.; Mani, R.S.; Müller, T.; Nadin-Davis, S.; Picard-Meyer, E.; Wilde, H.; et al. Rabies. Nat. Rev. Dis. Prim. 2017, 3, 17091. [Google Scholar] [CrossRef]
- Vega, S.; Lorenzo-Rebenaque, L.; Marin, C.; Domingo, R.; Fariñas, F. Tackling the Threat of Rabies Reintroduction in Europe. Front. Vet. Sci. 2021, 7, 613712. [Google Scholar] [CrossRef]
- Gunawardena, P.S.; Marston, D.A.; Ellis, R.J.; Wise, E.L.; Karawita, A.C.; Breed, A.C.; Fooks, A.R. Lyssavirus in Indian flying foxes Sri Lanka. Emerg. Infect. Dis. 2016, 22, 1456–1459. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Mayer, A.E.; Niezgoda, M.; Markotter, W.; Agwanda, B.; Breiman, R.F.; Rupprecht, C.E. Shimoni bat virus, a new representative of the Lyssavirus genus. Virus Res. 2010, 149, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Tordo, N.; Poch, O.; Ermine, A.; Keith, G.; Rougeon, F. Completion of the rabies virus genome sequence determi-nation: Highly conserved domains among the L (polymerase) proteins of unsegmented negative-strand RNA viruses. Virology 1988, 165, 565–576. [Google Scholar] [CrossRef]
- Gould, A.R.; Kattenbelt, J.A.; Gumley, S.G.; Lunt, R.A. Characterisation of an Australian bat lyssavirus variant isolated from an insectivorous bat. Virus Res. 2002, 89, 1–28. [Google Scholar] [CrossRef]
- Marston, D.A.; McElhinney, L.M.; Johnson, N.; Müller, T.; Conzelmann, K.K.; Tordo, N.; Fooks, A.R. Comparative analysis of the full genome sequence of European bat lyssavirus type 1 and type 2 with other lyssaviruses and evidence for a conserved transcription termination and polyadenylation motif in the G-L 3’ non-translated region. J. Gen. Virol. 2007, 88, 1302–1314. [Google Scholar] [CrossRef]
- Le Mercier, P.; Jacob, Y.; Tordo, N. The complete Mokola virus genome sequence: Structure of the RNA-dependent RNA polymerase. J. Gen. Virol. 1997, 78, 1571–1576. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, I.V.; Bozick, B.; Guagliardo, S.A.; Kunkel, R.; Shak, J.R.; Tong, S.; Rupprecht, C.E. Bats, emerging infectious diseases, and the rabies paradigm revisited. Emerg. Health. Threats. J. 2011, 4, 1. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Rupprecht, C.E. Bat Lyssaviruses. In Bats and Viruses: A New Frontier of Emerging Infectious Diseases; John Wiley and Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 47–97. [Google Scholar] [CrossRef]
- Dietz, C.; von Helversen, O.; Nill, D. Bats of Britain. Europe and Northwest Africa; A and C Black: London, UK, 2009; 400p. [Google Scholar]
- Arnaout, Y.; Djelouadji, Z.; Robardet, E.; Cappelle, J.; Cliquet, F.; Touzalin, F.; Jimenez, G.; Hurstel, S.; Borel, C.; Picard-Meyer, E. Genetic identification of bat species for pathogen surveillance across France. PLoS ONE 2022, 4, e0261344. [Google Scholar] [CrossRef]
- Çoraman, E.; Dundarova, H.; Dietz, C.; Mayer, F. Patterns of mtDNA introgression suggest population replacement in Palaearctic whiskered bat species. R. Soc. Open Sci. 2020, 7, 191805. [Google Scholar] [CrossRef]
- De Benedictis, P.; Leopardi, S.; Markotter, W.; Velasco-Villa, A. The Importance of Accurate Host Species Identification in the Framework of Rabies Surveillance, Control and Elimination. Viruses 2022, 14, 492. [Google Scholar] [CrossRef]
- Parize, P.; Travecedo Robledo, I.C.; Cervantes-Gonzalez, M.; Kergoat, L.; Larrous, F.; Serra-Cobo, J.; Dacheux, L.; Bourhy, H. Circumstances of Human–Bat interactions and risk of lyssavirus transmission in metropolitan France. Zoonoses Public Health 2020, 67, 774–784. [Google Scholar] [CrossRef]
- Salicini, I.; Ibáñez, C.; Juste, J. Multilocus phylogeny and species delimitation within the Natterer’s bat species complex in the Western Palearctic. Mol. Phylogenet. Evol. 2011, 61, 888–898. [Google Scholar] [CrossRef] [PubMed]
- Salicini, I.; Ibáñez, C.; Juste, J. Deep differentiation between and within Mediterranean glacial refugia in a flying mammal, the Myotis nattereri bat complex. J. Biogeogr. 2013, 40, 1182–1193. [Google Scholar] [CrossRef]
- Eggerbauer, E.; Troupin, C.; Passior, K.; Pfaff, F.; Höper, D.; Neubauer-Juric, A.; Haberl, S.; Bouchier, C.; Mettenleiter, T.C.; Bourhy, H.; et al. The Recently Discovered Bokeloh Bat Lyssavirus: Insights Into Its Genetic Heterogeneity and Spatial Distribution in Europe and the Population Genetics of Its Primary Host. Adv. Virus Res. 2017, 99, 199–232. [Google Scholar] [CrossRef]
- Çoraman, E.; Dietz, C.; Hempel, E.; Ghazaryan, A.; Levin, E.; Presetnik, P.; Zagmajster, M.; Mayer, F. Reticulate evolutionary history of a Western Palaearctic Bat Complex explained by multiple mtDNA introgressions in secondary contacts. J. Biogeogr. 2019, 46, 343–354. [Google Scholar] [CrossRef]
- Fooks, A.R.; Shipley, R.; Markotter, W.; Tordo, N.; Freuling, C.M.; Müller, T.; McElhinney, L.M.; Banyard, A.C.; Rupprecht, C.E. Renewed Public Health Threat from Emerging Lyssaviruses. Viruses 2021, 13, 1769. [Google Scholar] [CrossRef] [PubMed]
- Wright, P.G.R.; Newton, J.; Agnelli, P.; Budinski, I.; Salvo, I.D.; Flaquer, C.; Fulco, A.; Georgiakakis, P.; Martinoli, A.; Mas, M.; et al. Hydrogen isotopes reveal evidence of migration of Miniopterus schreibersii in Europe. BMC Ecolol. 2020, 20, 52. [Google Scholar] [CrossRef] [PubMed]
- Freuling, C.; Vos, A.; Johnson, N.; Kaipf, I.; Denzinger, A.; Neubert, L.; Mansfield, K.; Hicks, D.; Nuñez, A.; Tordo, N.; et al. Experimental infection of serotine bats (Eptesicus serotinus) with European bat lyssavirus type 1a. J. Gen. Virol. 2009, 90, 2493–2502. [Google Scholar] [CrossRef]
- de Thoisy, B.; Bourhy, H.; Delaval, M.; Pontier, D.; Dacheux, L.; Darcissac, E.; Donato, D.; Guidez, A.; Larrous, F.; Lavenir, R.; et al. Bioecological drivers of rabies virus circulation in a neotropical bat community. PLoS Negl. Trop. Dis. 2016, 10, e0004378. [Google Scholar] [CrossRef]
- Bonnaud, E.M.; Troupin, C.; Dacheux, L.; Holmes, E.C.; Monchatre-Leroy, E.; Tanguy, M.; Bouchier, C.; Cliquet, F.; Barrat, J.; Bourhy, H. Comparison of intra-and inter-host genetic diversity in rabies virus during experimental cross-species transmission. PLoS Pathog. 2019, 15, e1007799. [Google Scholar] [CrossRef]
- Colombi, D.; Serra-Cobo, J.; Métras, R.; Apolloni, A.; López-Roig, M.; Bourhy, H.; Colizza, V. Mechanisms for lyssavirus persistence in non-synanthropic bats in Europe: Insights from a modelling study. Sci. Rep. 2019, 9, 537. [Google Scholar] [CrossRef] [PubMed]
- Horton, D.L.; Breed, A.C.; Arnold, M.E.; Smith, G.C.; Aegerter, J.N.; McElhinney, L.M.; Johnson, N.; Banyard, A.C.; Raynor, R.; Mackie, I.; et al. Between roost contact is essential for maintenance of European bat lyssavirus type-2 in Myotis daubentonii bat reservoir: ‘The Swarming Hypothesis’. Sci. Rep. 2020, 10, 1740. [Google Scholar] [CrossRef]
- Tájek, P.; Tájková, P. A long distance migration in Nyctalus leisleri from the Czech Republic to southern France (Chiroptera: Vespertilionidae). Lynx Ser. Nova 2020, 51, 223–226. [Google Scholar] [CrossRef]
- Dundarova, H.; Michev, B.; Pandourski, I. Bats over the Western Black Sea open water area. Acta Zool. Bulg. 2021, 73, 543–546. [Google Scholar]
- Bouma, H.R.; Carey, H.V.; Kroese, F.G. Hibernation: The immune system at rest? J. Leukoc. Biol. 2010, 88, 619–624. [Google Scholar] [CrossRef]
- Field, K.A.; Sewall, B.J.; Prokkola, J.M.; Turner, G.G.; Gagnon, M.F.; Liley, T.M.; White, J.P.; Johnson, J.J.; Hauer, C.L.; Reeder, D.M. Effect of torpor on host transcriptomic responses to a fungal pathogen in hibernating bats. Mol. Ecol. 2018, 27, 3727–3743. [Google Scholar] [CrossRef]
- Constantine, D.G. Bat Rabies and Other Lyssavirus Infections; US Geological Survey: Reston, VA, USA, 2009; 68p. [Google Scholar]
- Harazim, M.; Perrot, J.; Varet, H.; Bourhy, H.; Lannoy, J.; Pikula, J.; Seidlová, V.; Dacheux, M.; Martínková, N. Transcriptomic responses of bat cells to European bat lyssavirus 1 infection under conditions simulating euthermia and hibernation. BMC Immunol. 2023, 24, 7. [Google Scholar] [CrossRef]
- Gilbert, A.T.; McCracken, G.F.; Sheeler, L.L.; Muller, L.I.; O’Rourke, D.; Kelch, W.J.; New, J.C., Jr. Rabies surveillance among bats in Tennessee, USA, 1996–2010. J. Wildl. Dis. 2015, 51, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Robardet, E.; Borel, C.; Moinet, M.; Jouan, D.; Wasniewski, M.; Barrat, J.; Boué, F.; Montchâtre-Leroy, E.; Servat, A.; Gimenez, O.; et al. Longitudinal survey of two serotine bat (Eptesicus serotinus) maternity colonies exposed to EBLV-1 (European Bat Lyssavirus type 1): Assessment of survival and serological status variations using capture-recapture models. PLoS Negl. Trop. Dis. 2017, 11, e0006048. [Google Scholar] [CrossRef] [PubMed]
- Shipley, R.; Wright, E.; Selden, D.; Wu, G.; Aegerter, J.; Fooks, A.R.; Banyard, A.C. Bats and viruses: Emergence of novel lyssaviruses and association of bats with viral zoonoses in the EU. Infect. Dis. Trop. Med. 2019, 4, 31. [Google Scholar] [CrossRef]
- Constantine, D.G.; Emmons, R.W.; Woodie, J.D. Rabies virus in nasal mucosa of naturally infected bats. Science 1972, 175, 1255–1256. [Google Scholar] [CrossRef]
- Allendorf, S.D.; Cortez, A.; Heinemann, M.B.; Harary, C.M.A.; Antunes, J.M.A.; Peres, M.G.; Vicente, A.F.; Sodré, M.M.; da Rosa, R.; Megid, J. Rabies virus distribution in tissues and molecular characterization of strains from naturally infected non-hematophagous bats. Virus Res. 2012, 165, 119–125. [Google Scholar] [CrossRef]
- Fisher, C.R.; Streicker, D.G.; Schnell, M.J. The spread and evolution of rabies virus: Conquering new frontiers. Nat. Rev. Microbiol. 2018, 16, 241–255. [Google Scholar] [CrossRef]
- Davis, A.; Rudd, R.J.; Bowen, R.A. Effects of Aerosolized Rabies Virus Exposure on Bats and Mice. J. Infect. Dis. 2007, 195, 151144–151150. [Google Scholar] [CrossRef]
- Johnson, N.; Phillpotts, R.; Fooks, A.R. Airborne transmission of lyssaviruses. J. Med. Microbiol. 2006, 55, 785–790. [Google Scholar] [CrossRef]
- Lojkić, I.; Šimić, I.; Bedeković, T.; Krešić, N. Current Status of Rabies and Its Eradication in Eastern and Southeastern Europe. Pathogens 2021, 10, 742. [Google Scholar] [CrossRef]
- Leopardi, S.; Barneschi, E.; Manna, G.; Zecchin, B.; Priori, P.; Drzewnioková, P.; Festa, F.; Lombardo, A.; Parca, F.; Scaravelli, D.; et al. Spillover of West Caucasian Bat Lyssavirus (WCBV) in a Domestic Cat and Westward Expansion in the Palearctic Region. Viruses 2021, 13, 2064. [Google Scholar] [CrossRef]
- Banyard, A.C.; Evans, J.S.; Luo, T.R.; Fooks, A.R. Lyssaviruses and bats: Emergence and zoonotic threat. Viruses 2014, 6, 2974–2990. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Niezgoda, M.; Franka, R.; Agwanda, B.; Markotter, W.; Beagley, J.C.; Urazova, O.Y.; Breiman, R.F.; Rupprecht, C.E. Lagos bat virus in Kenya. J. Clin. Microbio. 2008, 46, 1451–1461. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Turmelle, A.S.; Agwanda, B.; Markotter, W.; Niezgoda, M.; Breiman, R.F.; Rupprecht, C.E. Commerson’s Leaf-Nosed Bat (Hipposideros commersoni) is the Likely Reservoir of Shimoni Bat Virus. Vector-Borne Zoonotic Dis. 2011, 11, 1465–1470. [Google Scholar] [CrossRef]
- Lučan, R.K.; Bartonička, T.; Benda, P.; Bilgin, R.; Jedlička, P.; Nicolaou, H.; Reiter, A.; Shohdi, W.M.; Šálek, M.; Řeřucha, Š.; et al. Reproductive seasonality of the Egyptian fruit bat (Rousettus aegyptiacus) at the northern limits of its distribution. J. Mammal. 2014, 95, 1036–1042. [Google Scholar] [CrossRef]
- Benda, P.; Abi-Said, M.; Bartonička, T.; Bilgin, R.; Faizolahi, K.; Lučan, R.K.; Nicolaou, H.; Raiter, A.; Shohdi, W.M.; Uhrin, M.; et al. Rousettus aegyptiacus (Pteropodidae) in the Palaearctic: List of records and revision of the distribution range. Vespertilio 2011, 15, 3–36. [Google Scholar]
- Benda, P.; Abi Said, M.R.; Bou Jaoude, I.; Karanouh, R.; Lučan, R.K.; Sadek, R.; Ševčik, M.; Uhrin, M.; Horáček, I. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 13. Review of distribution and ectoparasites of bats in Lebanon. Acta Soc. Zool. Bohem. 2016, 80, 207–316. [Google Scholar]
- Willoughby, A.; Phelps, K.; Olival, K. A comparative analysis of viral richness and viral sharing in cave-roosting bats. Diversity 2017, 9, 35. [Google Scholar] [CrossRef]
- Wang, Z.; Huang, G.; Huang, M.; Dai, Q.; Hu, Y.; Zhou, J.; Wei, F. Global patterns of phylogenetic diversity and transmission of bat coronavirus. Sci. China Life Sci. 2022, 66, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Albery, G.F.; Becker, D.J.; Brierley, L.; Brook, C.E.; Christofferson, R.C.; Cohen, L.E.; Dallas, T.A.; Eskew, E.A.; Fagre, A.; Farrell, M.J.; et al. The science of the host-virus network. Nat. Microbiol. 2021, 6, 1483–1492. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dundarova, H.; Ivanova-Aleksandrova, N.; Bednarikova, S.; Georgieva, I.; Kirov, K.; Miteva, K.; Neov, B.; Ostoich, P.; Pikula, J.; Zukal, J.; et al. Phylogeographic Aspects of Bat Lyssaviruses in Europe: A Review. Pathogens 2023, 12, 1089. https://doi.org/10.3390/pathogens12091089
Dundarova H, Ivanova-Aleksandrova N, Bednarikova S, Georgieva I, Kirov K, Miteva K, Neov B, Ostoich P, Pikula J, Zukal J, et al. Phylogeographic Aspects of Bat Lyssaviruses in Europe: A Review. Pathogens. 2023; 12(9):1089. https://doi.org/10.3390/pathogens12091089
Chicago/Turabian StyleDundarova, Heliana, Nadya Ivanova-Aleksandrova, Sarka Bednarikova, Irina Georgieva, Krasimir Kirov, Kalina Miteva, Boyko Neov, Peter Ostoich, Jiri Pikula, Jan Zukal, and et al. 2023. "Phylogeographic Aspects of Bat Lyssaviruses in Europe: A Review" Pathogens 12, no. 9: 1089. https://doi.org/10.3390/pathogens12091089
APA StyleDundarova, H., Ivanova-Aleksandrova, N., Bednarikova, S., Georgieva, I., Kirov, K., Miteva, K., Neov, B., Ostoich, P., Pikula, J., Zukal, J., & Hristov, P. (2023). Phylogeographic Aspects of Bat Lyssaviruses in Europe: A Review. Pathogens, 12(9), 1089. https://doi.org/10.3390/pathogens12091089