
High Prevalence of Rickettsia raoultii Found in Dermacentor Ticks Collected in Barnaul, Altai Krai, Western Siberia

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Ticks
2.2. DNA Extraction and Quantitative PCR
2.3. Sequence Analysis
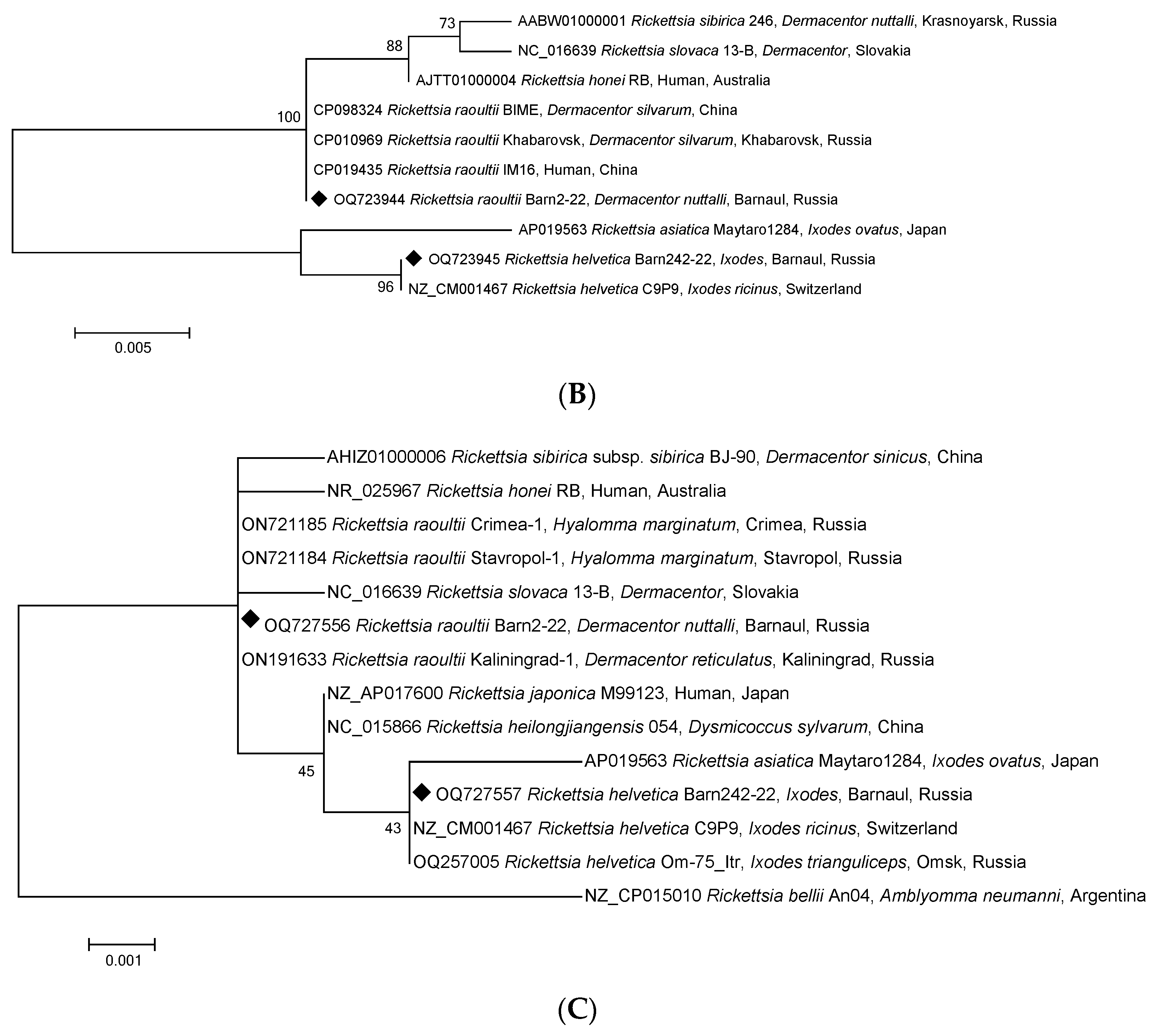
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rudakov, N.V.; Pen’evskaya, N.A.; Kumpan, L.V.; Blokh, A.I.; Shpynov, S.N.; Trankvilevsky, D.V.; Shtrek, S.V. Epidemiological Situation on Tick-Borne Spotted Fever Group Rickettsioses in the Russian Federation in 2012–2021, Prognosis for 2022–2026. Probl. Part. Danger. Infect. 2022, 1, 54–63. [Google Scholar] [CrossRef]
- Beskhlebova, O.V.; Granitov, V.M.; Shpynov, S.N.; Dedkov, V.G.; Arseneva, I.V.; Pantyukhina, A.N. Rickettsioses of spotted fever group in the Altai region. Infect. Dis. News Opin. Train. 2017, 19, 73–78. [Google Scholar] [CrossRef]
- Igolkina, Y.; Krasnova, E.; Rar, V.; Savelieva, M.; Epikhina, T.; Tikunov, A.; Khokhlova, N.; Provorova, V.; Tikunova, N. Detection of causative agents of tick-borne rickettsioses in Western Siberia, Russia: Identification of Rickettsia raoultii and Rickettsia sibirica DNA in clinical samples. Clin. Microbiol. Infect. 2018, 24, 199.e9–199.e12. [Google Scholar] [CrossRef] [PubMed]
- Rudakov, N.; Samoylenko, I.; Shtrek, S.; Igolkina, Y.; Rar, V.; Zhirakovskaia, E.; Tkachev, S.; Kostrykina, Y.; Blokhina, I.; Lentz, P.; et al. A fatal case of tick-borne rickettsiosis caused by mixed Rickettsia sibirica subsp. sibirica and “Candidatus Rickettsia tarasevichiae” infection in Russia. Ticks Tick Borne Dis. 2019, 10, 101278. [Google Scholar] [CrossRef] [PubMed]
- Roux, V.; Rydkyna, E.; Eremeeva, M.; Raoult, D. Citrate synthase gene comparison, a new tool for phylogenetic analysis, and its application for the rickettsiae. Int. J. Syst. Bacteriol. 1997, 47, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Regnery, R.L.; Spruill, C.L.; Plikaytis, B.D. Genotypic identification of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J. Bacteriol. 1991, 173, 1576–1589. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Jang, W.J.; Kim, J.H.; Ryu, J.S.; Lee, S.H.; Park, K.H.; Paik, H.S.; Koh, Y.S.; Choi, M.S.; Kim, I.S. Spotted fever group and typhus group rickettsioses in humans, South Korea. Emerg. Infect. Dis. 2005, 11, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Ishikura, M.; Ando, S.; Shinagawa, Y.; Matsuura, K.; Hasegawa, S.; Nakayama, T.; Fujita, H.; Watanabe, M. Phylogenetic analysis of spotted fever group rickettsiae based on gltA, 17-kDa, and rOmpA genes amplified by nested PCR from ticks in Japan. Microbiol. Immunol. 2003, 47, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Comparison of Infectious Disease Rates. Available online: https://www.iminfin.ru/areas-of-analysis/health/perechen-zabolevanij/sravnenie?territory=01000000 (accessed on 29 June 2023).
- Mediannikov, O.; Matsumoto, K.; Samoylenko, I.; Drancourt, M.; Roux, V.; Rydkina, E.; Davoust, B.; Tarasevich, I.; Brouqui, P.; Fournier, P.E. Rickettsia raoultii sp. nov., a spotted fever group rickettsia associated with Dermacentor ticks in Europe and Russia. Int. J. Syst. Evol. Microbiol. 2008, 58, 1635–1639. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.G.; Kwon, O.D.; Kwak, D. High prevalence of Rickettsia raoultii and associated pathogens in canine ticks, South Korea. Emerg. Infect. Dis. 2020, 26, 2532. [Google Scholar] [CrossRef] [PubMed]
- Balážová, A.; Földvári, G.; Bilbija, B.; Nosková, E.; Široký, P. High prevalence and low diversity of Rickettsia in Dermacentor reticulatus ticks, Central Europe. Emerg. Infect. Dis. 2022, 28, 893–895. [Google Scholar] [CrossRef] [PubMed]
- Dedkov, V.G.; Simonova, E.G.; Beshlebova, O.V.; Safonova, M.V.; Stukolova, O.A.; Verigina, E.V.; Savinov, G.V.; Karaseva, I.P.; Blinova, E.A.; Granitov, V.M.; et al. The burden of tick-borne diseases in the Altai region of Russia. Ticks Tick Borne Dis. 2017, 8, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, K.; Elfving, K.; Pahlson, C. Rickettsia helvetica in patient with meningitis, Sweden, 2006. Emerg. Infect. Dis. 2010, 16, 490–492. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, K. Septicaemia with Rickettsia helvetica in a patient with acute febrile illness, rash and myasthenia. J. Infect. 2009, 58, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, K.; Lindquist, O.; Påhlson, C. Association of Rickettsia helvetica with chronic perimyocarditis in sudden cardiac death. Lancet 1999, 9185, 1169–1173. [Google Scholar] [CrossRef] [PubMed]
- Speck, S.; Kern, T.; Aistleitner, K.; Dilcher, M.; Dobler, G.; Essbauer, S. In vitro studies of Rickettsia-host cell interactions: Confocal laser scanning microscopy of Rickettsia helvetica-infected eukaryotic cell lines. PLoS Negl. Trop. Dis. 2018, 12, e0006151. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) | Annealing Temperature (°C) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| Rp877p | GGGGACCTGCTCACGGCGG | 58 | 382 | [5] |
| Rp1258n | ATTGCAAAAAGTACCGTGAACA | |||
| Rr.190.70p | ATGGCGAATATTTCTCCAAAA | 55 | 532 | [6] |
| Rr.190.602n | AGTGCAGCATTCGCTCCCCCT | |||
| rompB SFG IF | GTTTAATACGTGCTGCTAACCAA | 57 | 426 | [7] |
| rompB SFG/TG IR | GGTTTTGCCCATATACCGTAAG | |||
| Rr17k.90p | GCTCTTGCAGCTTCTATGTT | 55 | 450 | [8] |
| Rr17k.539n | TCAATTCACAACTTGCCATT | |||
| 16S3 | GATGGATGAGCCCGCGTCAG | 65 | 772 | [3] |
| 16S4 | GCATCTCTGCGATCCGCGAC |
| Genus | Collecting Zones | Number of Ticks | Number of Ticks Infected by SFGR (%, 95% CI) | |
|---|---|---|---|---|
| R. raoultii | R. helvetica | |||
| Dermacentor | 1 | 71 | 37 (52.1, 40.7–63.3) | 0 |
| 2 | 60 | 33 (55.0, 42.5–66.9) | 0 | |
| 3 | 11 | 11 (100.0, 70.0–100.0) | 0 | |
| 4 | 1 | 0 | 0 | |
| 5 | 57 | 42 (73.7, 60.9–83.4) | 0 | |
| 6 | 2 | 2 (100.0, 29.0–100.0) | 0 | |
| 7 | - | - | - | |
| Subtotal | 202 | 125 (61.9, 55.0–68.3) | 0 | |
| Ixodes | 1 | - | - | - |
| 2 | - | - | - | |
| 3 | - | - | - | |
| 4 | 11 | 0 | 0 | |
| 5 | 5 | 0 | 2 (40.0, 11.6–77.1) | |
| 6 | 39 | 0 | 2 (5.1, 0.5–17.8) | |
| 7 | 42 | 0 | 1 (2.4, <0.01–13.4) | |
| Subtotal | 97 | 0 | 5 (5.1, 1.9–11.8) | |
| Haemaphysalis | 7 | 1 | 0 | 0 |
| Total | 300 | 125 (41.7, 36.2–47.3) | 5 (1.7, 0.6–4.0) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rakov, A.V.; Chekanova, T.A.; Petremgvdlishvili, K.; Timonin, A.V.; Valdokhina, A.V.; Shirokostup, S.V.; Lukyanenko, N.V.; Akimkin, V.G. High Prevalence of Rickettsia raoultii Found in Dermacentor Ticks Collected in Barnaul, Altai Krai, Western Siberia. Pathogens 2023, 12, 914. https://doi.org/10.3390/pathogens12070914
Rakov AV, Chekanova TA, Petremgvdlishvili K, Timonin AV, Valdokhina AV, Shirokostup SV, Lukyanenko NV, Akimkin VG. High Prevalence of Rickettsia raoultii Found in Dermacentor Ticks Collected in Barnaul, Altai Krai, Western Siberia. Pathogens. 2023; 12(7):914. https://doi.org/10.3390/pathogens12070914
Chicago/Turabian StyleRakov, Alexey V., Tatiana A. Chekanova, Ketevan Petremgvdlishvili, Andrey V. Timonin, Anna V. Valdokhina, Sergey V. Shirokostup, Natalia V. Lukyanenko, and Vasiliy G. Akimkin. 2023. "High Prevalence of Rickettsia raoultii Found in Dermacentor Ticks Collected in Barnaul, Altai Krai, Western Siberia" Pathogens 12, no. 7: 914. https://doi.org/10.3390/pathogens12070914
APA StyleRakov, A. V., Chekanova, T. A., Petremgvdlishvili, K., Timonin, A. V., Valdokhina, A. V., Shirokostup, S. V., Lukyanenko, N. V., & Akimkin, V. G. (2023). High Prevalence of Rickettsia raoultii Found in Dermacentor Ticks Collected in Barnaul, Altai Krai, Western Siberia. Pathogens, 12(7), 914. https://doi.org/10.3390/pathogens12070914