
Multi-Trait Genome-Wide Association Studies of Sorghum bicolor Regarding Resistance to Anthracnose, Downy Mildew, Grain Mold and Head Smut


Abstract
1. Introduction
2. Materials and Methods
2.1. Sorghum Association Panel
2.2. Sorghum Mini Core Collection
2.3. Senegalese Sorghum Population
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ananda, G.K.S.; Myrans, H.; Norton, S.L.; Gleadow, R.; Furtado, A.; Henry, R.J. Wild sorghum as a promising resource for crop improvement. Front. Plant Sci. 2020, 11, 1108. [Google Scholar] [CrossRef] [PubMed]
- Koima, I.N.; Kilalo, D.C.; Orek, C.O.; Wagacha, J.M.; Nyaboga, E.N. Identification and characterization of Colletotrichum species causing sorghum anthracnose in Kenya and screening of sorghum germplasm for resistance to anthracnose. J. Fungi 2023, 9, 100. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Rao, V.P.; Upadhyaya, H.D.; Reddy, V.G.; Thakur, R.P. Resistance to grain mold and downy mildew in a Mini-core collection of sorghum germplasm. Plant Dis. 2010, 94, 439–444. [Google Scholar] [CrossRef]
- Prom, L.K.; Perumal, R.; Erattaimuthu, S.R.; Erpelding, J.E.; Montes, N.; Odvody, G.N.; Greenwald, C.; Jin, Z.; Frederiksen, R.; Magill, C.W. Virulence and molecular genotyping studies of Sporisorium reilianum isolates in sorghum. Plant Dis. 2011, 95, 523–529. [Google Scholar] [CrossRef]
- Ahn, E.; Prom, L.K.; Fall, C.; Magill, C. Response of Senegalese sorghum seedlings to pathotype 5 of Sporisorium reilianum. Crops 2022, 2, 142–153. [Google Scholar] [CrossRef]
- Uffelmann, E.; Huang, Q.Q.; Munung, N.S.; de Vries, J.; Okada, Y.; Martin, A.R.; Martin, H.C.; Lappalainen, T.; Posthuma, D. Genome-wide association studies. Nat. Rev. Methods Primers 2021, 1, 59. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, H.; Yang, W.; Zhao, W.; Tong, C. Multivariate linear mixed model enhanced the power of identifying genome-wide association to poplar tree heights in a randomized complete block design. G3 Genes Genomes Genet. 2021, 11, jkaa053. [Google Scholar] [CrossRef]
- Rice, B.R.; Fernandes, S.B.; Lipka, A.E. Multi-trait genome-wide association studies reveal loci associated with maize inflorescence and leaf architecture. Plant Cell Physiol. 2020, 61, 1427–1437. [Google Scholar] [CrossRef]
- Aschard, H.; Vilhjálmsson, B.J.; Greliche, N.; Morange, P.E.; Trégouët, D.A.; Kraft, P. Maximizing the power of principal-component analysis of correlated phenotypes in genome-wide association studies. Am. J. Hum. Genet. 2014, 94, 662–676. [Google Scholar] [CrossRef]
- Ahn, E.; Fall, C.; Prom, L.K.; Magill, C. Genome-wide association study of Senegalese sorghum seedlings responding to a Texas isolate of Colletotrichum sublineola. Sci. Rep. 2022, 12, 13025. [Google Scholar] [CrossRef] [PubMed]
- Ahn, E.; Fall, C.; Prom, L.K.; Magill, C. A genome-wide association study of Senegalese sorghum seedlings responding to pathotype 5 of Sporisorium reilianum. Plants 2022, 11, 2999. [Google Scholar] [CrossRef]
- Prom, L.K.; Cuevas, H.E.; Ahn, E.; Isakeit, T.; Rooney, W.L.; Magill, C. Genome-wide association study of grain mold resistance in sorghum association panel as affected by inoculation with Alternaria alternata alone and Alternaria alternata, Fusarium thapsinum, and Curvularia lunata combined. Eur. J. Plant Pathol. 2020, 157, 783–798. [Google Scholar] [CrossRef]
- Ahn, E.; Hu, Z.; Perumal, R.; Prom, L.K.; Odvody, G.; Upadhyaya, H.D.; Magill, C.W. Genome wide association analysis of sorghum mini core lines regarding anthracnose, downy mildew, and head smut. PLoS ONE 2019, 14, e0216671. [Google Scholar] [CrossRef]
- Ahn, E.; Prom, L.K.; Hu, Z.; Odvody, G.; Magill, C.W. Genome-wide association analysis for response of Senegalese sorghum accessions to Texas isolates of anthracnose. Plant Genome 2021, 14, e20097. [Google Scholar] [CrossRef]
- Cuevas, H.E.; Prom, L.K.; Cruet-Burgos, C.M. Genome-wide association mapping of anthracnose (Colletotrichum sublineolum) resistance in NPGS Ethiopian sorghum germplasm. G3 Genes Genomes Genet. 2019, 9, 2879–2885. [Google Scholar] [CrossRef]
- Cuevas, H.E.; Prom, L.K.; Cooper, E.A.; Knoll, J.E.; Ni, X. Genome-wide association mapping of anthracnose (Colletotrichum sublineolum) resistance in the U.S. sorghum association panel. Plant Genome 2018, 11, 170099. [Google Scholar] [CrossRef]
- Prom, L.K.; Ahn, E.; Magill, C. SNPs that identify alleles with highest effect on grain mold ratings after inoculation with Alternaria alternata or with a mixture of Alternaria alternata, Fusarium thapsinum and Curvularia lunata. J. Agric. Crop Res. 2021, 9, 72–79. [Google Scholar]
- Morris, G.P.; Ramu, P.; Deshpande, S.P.; Hash, C.T.; Shah, T.; Upadhyaya, H.D.; Riera-Lizarazu, O.; Brown, P.J.; Acharya, C.B.; Mitchell, S.E.; et al. Population genomic and genome-wide association studies of agroclimatic traits in sorghum. Proc. Natl. Acad. Sci. USA 2013, 110, 453–458. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Adeyanju, A.; Little, C.; Yu, J.; Tesso, T. Genome-wide association study on resistance to stalk rot diseases in grain sorghum. G3 Genes Genomes Genet. 2015, 5, 1165–1175. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Wang, Y.H.; Gowda, C.L.L.; Sharma, S. Association mapping of maturity and plant height using SNP markers with the sorghum mini core collection. Theor. Appl. Genet. 2013, 126, 2003–2015. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Upadhyaya, H.D.; Burrell, A.M.; Sahraeian, S.M.E.; Klein, R.R.; Klein, P.E. Genetic structure and linkage disequilibrium in a diverse, representative collection of the C4 model plant, Sorghum bicolor. G3 Genes Genomes Genet. 2013, 3, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high siversity apecies. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Olatoye, M.O.; Marla, S.; Morris, G.P. An integrated genotyping-by-sequencing polymorphism map for over 10,000 sorghum genotypes. Plant Genome 2019, 12, 180044. [Google Scholar] [CrossRef]
- Yu, J.; Pressoir, G.; Briggs, W.H.; Vroh Bi, I.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S.; Nielsen, D.M.; Holland, J.B.; et al. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nat. Genet 2006, 38, 203–208. [Google Scholar] [CrossRef]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef]
- McCormick, R.F.; Truong, S.K.; Sreedasyam, A.; Jenkins, J.; Shu, S.; Sims, D.; Kennedy, M.; Amirebrahimi, M.; Weers, B.D.; McKinley, B.; et al. The Sorghum bicolor reference genome: Improved assembly, gene annotations, a transcriptome atlas, and signatures of genome organization. Plant J. 2018, 93, 338–354. [Google Scholar] [CrossRef]
- Prom, L.K.; Ahn, E.; Isakeit, T.; Magill, C. GWAS analysis of sorghum association panel lines identifies SNPs associated with disease response to Texas isolates of Colletotrichum sublineola. Theor. Appl. Genet. 2019, 132, 1389–1396. [Google Scholar] [CrossRef]
- Hwang, I.S.; Hwang, B.K. The pepper mannose-binding lectin gene CaMBL1 is required to regulate cell death and defense responses to microbial pathogens. Plant Physiol. 2011, 155, 447–463. [Google Scholar] [CrossRef]
- Liu, Y.; Khan, A.R.; Gan, Y. C2H2 zinc finger proteins response to abiotic stress in plants. Int. J. Mol. Sci. 2022, 23, 2730. [Google Scholar] [CrossRef]
- Xu, R.; Guo, Y.; Peng, S.; Liu, J.; Li, P.; Jia, W.; Zhao, J. Molecular targets and biological functions of cAMP signaling in Arabidopsis. Biomolecules 2021, 11, 688. [Google Scholar] [CrossRef]
- Mo, B.; Bewley, J.D. The relationship between beta-mannosidase and endo-beta-mannanase activities in tomato seeds during and following germination: A comparison of seed populations and individual seeds. J. Exp. Bot. 2003, 54, 2503–2510. [Google Scholar] [CrossRef]
- Garcia-Seco, D.; Chiapello, M.; Bracale, M.; Pesce, C.; Bagnaresi, P.; Dubois, E.; Moulin, L.; Vannini, C.; Koebnik, R. Transcriptome and proteome analysis reveal new insight into proximal and distal responses of wheat to foliar infection by Xanthomonas translucens. Sci. Rep. 2017, 7, 10157. [Google Scholar] [CrossRef]
- Dong, M.; Gu, J.; Zhang, L.; Chen, P.; Liu, T.; Deng, J.; Lu, H.; Han, L.; Zhao, B. Comparative proteomics analysis of superior and inferior spikelets in hybrid rice during grain filling and response of inferior spikelets to drought stress using isobaric tags for relative and absolute quantification. J. Proteom. 2014, 109, 382–399. [Google Scholar] [CrossRef]
- Butt, H.; Bazin, J.; Prasad, K.; Awad, N.; Crespi, M.; Reddy, A.S.N.; Mahfouz, M.M. The rice serine/arginine splicing factor RS33 regulates pre-mRNA splicing during abiotic stress responses. Cells 2022, 11, 1796. [Google Scholar] [CrossRef]
- Yang, S.; Tang, F.; Zhu, H. Alternative splicing in plant immunity. Int. J. Mol. Sci. 2014, 15, 10424–10445. [Google Scholar] [CrossRef]
- Kim, M.; Lim, J.H.; Ahn, C.S.; Park, K.; Kim, G.T.; Kim, W.T.; Pai, H.S. Mitochondria-associated hexokinases play a role in the control of programmed cell death in Nicotiana benthamiana. Plant Cell 2006, 18, 2341–2355. [Google Scholar] [CrossRef]
- Fernández-González, M.; González-Fernández, E.; Fernández-González, D.; Rodríguez-Rajo, F.J. Secondary outcomes of the ole e 1 proteins involved in pollen tube development: Impact on allergies. Front. Plant Sci. 2020, 11, 974. [Google Scholar] [CrossRef]
- Jeandet, P.; Formela-Luboińska, M.; Labudda, M.; Morkunas, I. The role of sugars in plant responses to stress and their regulatory function during development. Int. J. Mol. Sci. 2022, 23, 5161. [Google Scholar] [CrossRef]
- Girma, G.; Nida, H.; Seyoum, A.; Mekonen, M.; Nega, A.; Lule, D.; Dessalegn, K.; Bekele, A.; Gebreyohannes, A.; Adeyanju, A.; et al. A large-scale genome-wide association analyses of Ethiopian sorghum landrace collection reveal loci associated with important traits. Front. Plant Sci. 2019, 10, 691. [Google Scholar] [CrossRef]
- Kumar, P.; He, Y.; Singh, R.; Davis, R.F.; Guo, H.; Paterson, A.H.; Peterson, D.G.; Shen, X.; Nichols, R.L.; Chee, P.W. Fine mapping and identification of candidate genes for a QTL affecting Meloidogyne incognita reproduction in Upland cotton. BMC Genom. 2016, 17, 567. [Google Scholar] [CrossRef] [PubMed]
- Casarrubia, S.; Martino, E.; Daghino, S.; Kohler, A.; Morin, E.; Khouja, H.R.; Murat, C.; Barry, K.W.; Lindquist, E.A.; Martin, F.M.; et al. Modulation of plant and fungal gene expression upon Cd exposure and symbiosis in ericoid mycorrhizal Vaccinium myrtillus. Front. Microbiol. 2020, 11, 341. [Google Scholar] [CrossRef] [PubMed]
- Venezia, M.; Creasey Krainer, K.M. Current advancements and limitations of gene editing current advancements and limitations of gene editing in orphan crops. Front. Plant Sci. 2021, 12, 742932. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
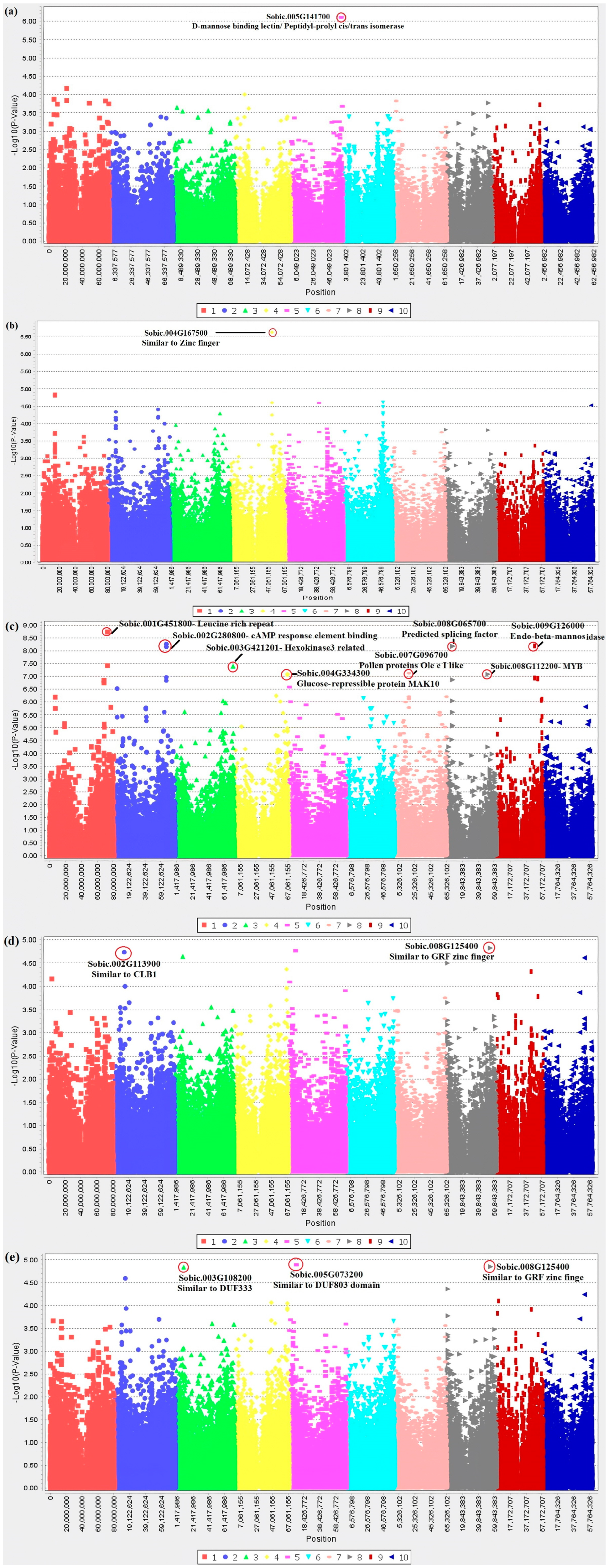
| Population | Traits | Chr | Location | Candidate Gene and Function | Base Pairs | SNP Base % | TASSEL p-Value | SNPs with High Linkage Disequilibrium (LD) (0.7 < R2), Closest Gene and Function |
|---|---|---|---|---|---|---|---|---|
| SAP | Grain mold under three treatments | 5 | 60278659 | Sobic.005G141700 Uncharacterized Associated PlantFAMs- D-mannose binding lectin/Peptidyl-prolyl cis/trans isomerase | 3147 | A: 13.1% C: 86.9% | 0.000000774 Bonferroni ≈ 0.00000041 | S05_60274552 S05_60274571 Nearest gene Sobic.005G141700 |
| Mini Core | Anthracnose + Downy mildew + Head smut | 4 | 51771351 | Sobic.004G167500 Uncharacterized Functional annotation- Zinc finger, RING/FYVE/PHD-type | 381 | C: 79.9% T: 20.1% | 0.000000236 * Bonferroni ≈ 0.0000003 | None |
| Senegalese | Anthracnose with three variates | 1 | 72868925 and multiple SNPs nearby | Sobic.001G451800 Protein kinase domain//Leucine-rich repeat N-terminal domain | 0 | G: 79.9% T: 20.1% | 0.00000000188 * Bonferroni ≈ 0.00000017 | S01_72831422 Nearest gene Sobic.001G451400- Vacuole morphology and inheritance protein 14 |
| 2 | 66200847 and multiple SNPs nearby | Sobic.002G280800 cAMP response element binding protein (CREB) Associated PlantFAMs- bZIP transcription factor domain | 2920 | C: 20.1% G: 79.9% | 0.00000000553 * Bonferroni ≈ 0.00000017 | S02_66173353 Nearest gene Sobic.002G280300 PF05553–Cotton fiber expressed protein (DUF761) S02_66179471 S02_66179497 Nearest gene Sobic.002G280400 Acyl carrier protein /Zinc finger protein 593-related S02_66186569 S02_66188204 S02_66188632 S02_66188704 S02_66188732 Nearest gene Sobic.002G280600 Mediator of RNA polyerase II transcription subunit 4 S02_66189758 S02_66189765 S02_66189995 S02_66190001 S02_66190035 S02_66194427 S02_66194452 S02_66196956 S02_66196986 Nearest gene Sobic.002G280700 Histone-binding protein RBBP4 S02_66200592 S02_66200825 Nearest gene Sobic.002G280800 Camp-response element binding protein-related | ||
| 9 | 47938177 and multiple SNPs nearby | Sobic.009G126000 Mannosyl glycoprotein endo-beta-mannosidase | 0 | A: 17.0% G: 83% | 0.00000000645 * Bonferroni ≈ 0.00000017 | None | ||
| 8 | 7370058 and multiple SNPs nearby | Sobic.008G065700 Uncharacterized Associated PlantFAMs- Predicted splicing factor | 1117 | A: 22.8% C: 77.2% | 0.0000000067 * Bonferroni ≈ 0.00000017 | S08_7327804 S08_7345722 Nearest gene Sobic.008G065600 Auxin responsive GH3 gene family S08_7363922 Nearest gene Sobic.008G065700 | ||
| 3 | 72625166 | Sobic.003G421201 Hexokinase-3 related | 0 | A: 21.0% G: 79.0% | 0.0000000404 * Bonferroni ≈ 0.00000017 | S03_72617056 S03_72617102 Nearest gene Sobic.003G421100 Gulonolactone oxidase | ||
| 7 | 17951013 | Sobic.007G096700 Pollen proteins Ole e I like | 0 | G: 85.5% T: 14.5% | 0.0000000752 * Bonferroni ≈ 0.00000017 | S07_17760523 S07_17760525 S07_17760589 S07_17761449 S07_17761552 Nearest gene Sobic.007G096000 Enhancer of polycomb S07_17831891 S07_17831935 Nearest gene Sobic.007G096200 Nucleoporin nup43 S07_17882047 S07_17882072 S07_17882080 Nearest gene Sobic.007G096401 Uncharacterized | ||
| 4 | 66666642 and multiple SNPs nearby | Sobic.004G334300 Similar to Glucose-repressible protein MAK10 | 0 | A: 32.5% G: 67.5% | 0.0000000813 * Bonferroni ≈ 0.00000017 | S04_66651495 Nearest gene Monogalactosyldiacylglycerol synthase 2 | ||
| 8 | 51886715 | Sobic.008G112200 MYB transcription factor | 0 | C: 91.2% T: 8.8% | 0.0000000834 * Bonferroni ≈ 0.00000017 | S08_51822410 S08_51822572 Nearest gene Sobic.008G112000 Oxidoreductase, 2OG-FE II S08_51868277 S08_51868331 Nearest gene Sobic.008G112100 Lysophosphatidic acid acyltransferase / lysophosphatidylinositol acyltransferase S08_51886688 S08_51886691 Nearest gene Sobic.008G112200 | ||
| Head smut with two variates | 8 | 54897663 | Sobic.008G125400 Uncharacterized Associated PlantFAMs- GRF zinc finger (zf-GRF) | 0 | C: 70.8% G: 29.2% | 0.000015 Bonferroni ≈ 0.00000017 | None | |
| 2 | 13906340 | Sobic.002G113900 Uncharacterized Similar to calcium-dependent lipid binding (CLB1) protein | 0 | A: 67.0% G: 33.0% | 0.0000184 Bonferroni ≈ 0.00000017 | None | ||
| Anthracnose + Head smut | 5 | 8867065 | Sobic.005G073200 Uncharacterized Associated PlantFAMs- DUF803 domain containing | 0 | A: 37.8% T: 62.2% | 0.000013 Bonferroni ≈ 0.00000017 | None | |
| 8 | 54897663 | Sobic.008G125400 Uncharacterized Associated PlantFAMs- GRF zinc finger (zf-GRF) | 0 | C: 70.8% G: 29.2% | 0.0000143 Bonferroni ≈ 0.00000017 | Identical SNP above | ||
| 3 | 9787536 | Sobic.003G108200 Uncharacterized Associated PlantFAMs- protein of unknown function (DUF3339) | 0 | C: 48.6% T: 51.4% | 0.0000146 Bonferroni ≈ 0.00000017 | None |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, E.; Prom, L.K.; Magill, C. Multi-Trait Genome-Wide Association Studies of Sorghum bicolor Regarding Resistance to Anthracnose, Downy Mildew, Grain Mold and Head Smut. Pathogens 2023, 12, 779. https://doi.org/10.3390/pathogens12060779
Ahn E, Prom LK, Magill C. Multi-Trait Genome-Wide Association Studies of Sorghum bicolor Regarding Resistance to Anthracnose, Downy Mildew, Grain Mold and Head Smut. Pathogens. 2023; 12(6):779. https://doi.org/10.3390/pathogens12060779
Chicago/Turabian StyleAhn, Ezekiel, Louis K. Prom, and Clint Magill. 2023. "Multi-Trait Genome-Wide Association Studies of Sorghum bicolor Regarding Resistance to Anthracnose, Downy Mildew, Grain Mold and Head Smut" Pathogens 12, no. 6: 779. https://doi.org/10.3390/pathogens12060779
APA StyleAhn, E., Prom, L. K., & Magill, C. (2023). Multi-Trait Genome-Wide Association Studies of Sorghum bicolor Regarding Resistance to Anthracnose, Downy Mildew, Grain Mold and Head Smut. Pathogens, 12(6), 779. https://doi.org/10.3390/pathogens12060779