
Identification of the Novel Streptococcus equi subsp. zooepidemicus Sequence Type 525 in Donkeys of Abruzzo Region, Italy

 , , , , , ,
, , , , , ,
Abstract
1. Introduction
2. Anamnesis
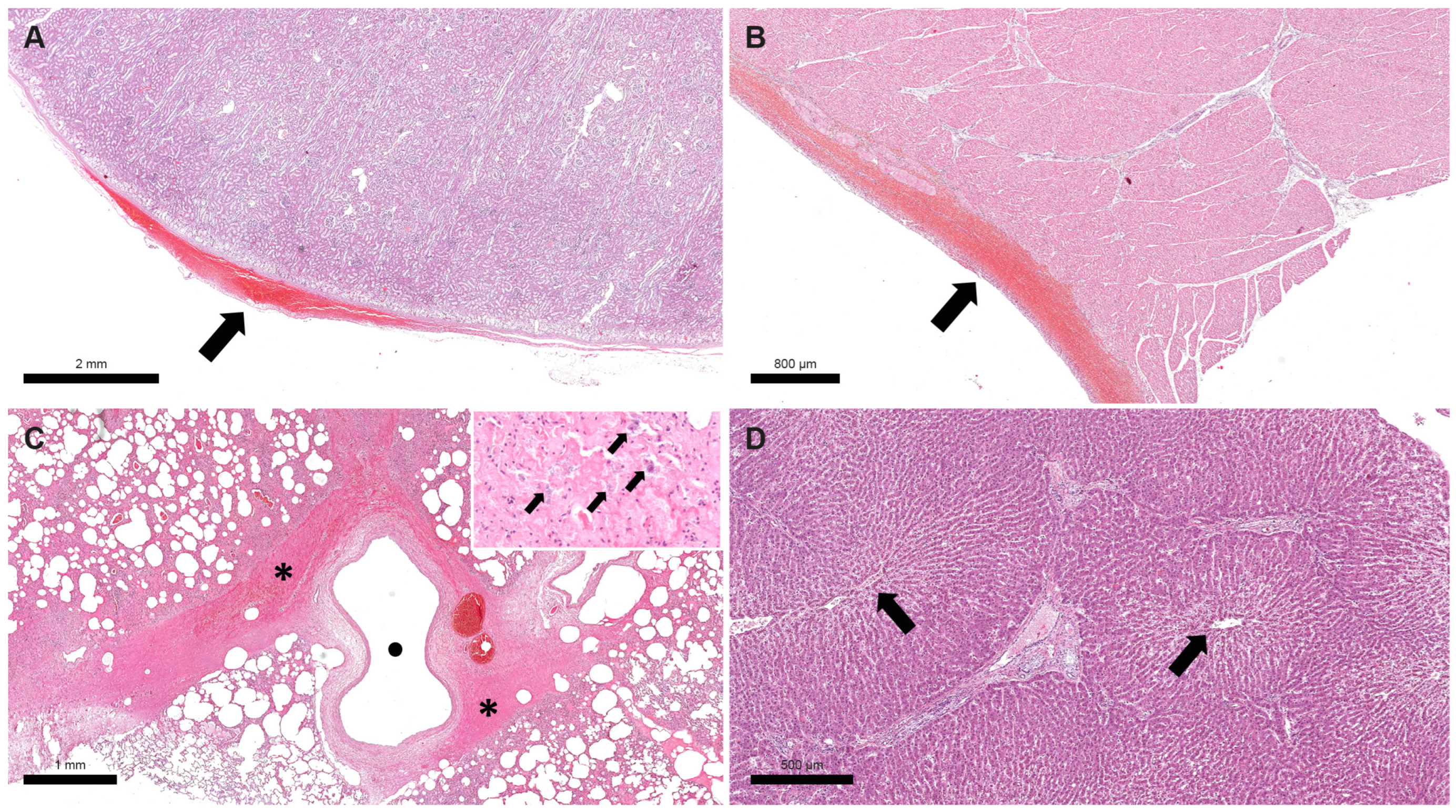
3. Clinical Signs, Necropsy, and Histopathology
4. Bacterial Isolation and Identification
5. Antimicrobial Susceptibility Test
6. Differential Diagnosis
7. Molecular Investigation
8. DNA Extraction and Sequencing
9. Discussion
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Sellon, D.C.; Long, M.T. Equine Infectious Diseases E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2013; ISBN 978-1-4557-5115-0. [Google Scholar]
- Preziuso, S.; Moriconi, M.; Cuteri, V. Genetic Diversity of Streptococcus Equi Subsp. Zooepidemicus Isolated from Horses. Comp. Immunol. Microbiol. Infect Dis. 2019, 65, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Båverud, V.; Johansson, S.K.; Aspan, A. Real-Time PCR for Detection and Differentiation of Streptococcus Equi Subsp. Equi and Streptococcus Equi Subsp. Zooepidemicus. Vet. Microbiol. 2007, 124, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Kudirkiene, E.; Welker, M.; Knudsen, N.R.; Bojesen, A.M. Rapid and Accurate Identification of Streptococcus Equi Subspecies by MALDI-TOF MS. Syst. Appl. Microbiol. 2015, 38, 315–322. [Google Scholar] [CrossRef]
- Mani, R.J.; Thachil, A.J.; Ramachandran, A. Discrimination of Streptococcus Equi Subsp. Equi and Streptococcus Equi Subsp. Zooepidemicus Using Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry. J. Vet. Diagn. Investig. 2017, 29, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Las Heras, A.; Vela, A.I.; Fernández, E.; Legaz, E.; Domínguez, L.; Fernández-Garayzábal, J.F. Unusual Outbreak of Clinical Mastitis in Dairy Sheep Caused by Streptococcus Equi Subsp. Zooepidemicus. J. Clin. Microbiol. 2002, 40, 1106–1108. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Jee, H.; Shin, S.W.; Lee, B.C.; Pakhrin, B.; Yoo, H.S.; Yoon, J.H.; Kim, D.Y. Outbreak and Control of Haemorrhagic Pneumonia Due to Streptococcus Equi Subspecies Zooepidemicus in Dogs. Vet. Rec. 2007, 161, 528–530. [Google Scholar] [CrossRef]
- Eyre, D.W.; Kenkre, J.S.; Bowler, I.C.J.W.; McBride, S.J. Streptococcus Equi Subspecies Zooepidemicus Meningitis--a Case Report and Review of the Literature. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 1459–1463. [Google Scholar] [CrossRef] [PubMed]
- Priestnall, S.; Erles, K. Streptococcus Zooepidemicus: An Emerging Canine Pathogen. Vet. J. 2011, 188, 142–148. [Google Scholar] [CrossRef]
- Minces, L.R.; Brown, P.J.; Veldkamp, P.J. Human Meningitis from Streptococcus equi subsp. Zooepidemicus Acquired as Zoonoses. Epidemiol. Infect. 2011, 139, 406–410. [Google Scholar] [CrossRef]
- Gruszynski, K.; Young, A.; Levine, S.J.; Garvin, J.P.; Brown, S.; Turner, L.; Fritzinger, A.; Gertz, R.E.; Murphy, J.M.; Vogt, M.; et al. Streptococcus Equi Subsp. Zooepidemicus Infections Associated with Guinea Pigs. Emerg. Infect. Dis. 2015, 21, 156–158. [Google Scholar] [CrossRef]
- Pelkonen, S.; Lindahl, S.B.; Suomala, P.; Karhukorpi, J.; Vuorinen, S.; Koivula, I.; Väisänen, T.; Pentikäinen, J.; Autio, T.; Tuuminen, T. Transmission of Streptococcus Equi Subspecies Zooepidemicus Infection from Horses to Humans. Emerg. Infect. Dis. 2013, 19, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Bosica, S.; Chiaverini, A.; Angelis, M.E.D.; Petrini, A.; Averaimo, D.; Martino, M.; Rulli, M.; Saletti, M.A.; Cantelmi, M.C.; Ruggeri, F.; et al. Early Release—Severe Streptococcus Equi Subspecies Zooepidemicus Outbreak from Unpasteurized Dairy Product Consumption, Italy. Emerg. Infect. Dis. J. 2023, 29, 5. [Google Scholar] [CrossRef]
- Gao, N. Endometritis in Donkeys Associated with Streptococcus Equi Subspecies Zooepidemicus Infection. PVJ 2020, 40, 537–539. [Google Scholar] [CrossRef] [PubMed]
- Bazzano, M.; McLean, A.; Tesei, B.; Gallina, E.; Laus, F. Selenium and Vitamin E Concentrations in a Healthy Donkey Population in Central Italy. J. Equine Vet. Sci. 2019, 78, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Papademas, P.; Mousikos, P.; Aspri, M. Valorization of Donkey Milk: Technology, Functionality, and Future Prospects. JDS Commun. 2022, 3, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Barrandeguy, M.E.; Carossino, M. Infectious Diseases in Donkeys and Mules: An Overview and Update. J. Equine Vet. Sci. 2018, 65, 98–105. [Google Scholar] [CrossRef]
- Câmara, R.J.F.; Bueno, B.L.; Resende, C.F.; Balasuriya, U.B.R.; Sakamoto, S.M.; dos Reis, J.K.P. Viral Diseases That Affect Donkeys and Mules. Animals 2020, 10, 2203. [Google Scholar] [CrossRef]
- Wilson, W.D.; Magdesian, K.G. Antimicrobial Selection for the Equine Practitioner. Vet. Clin. N. Am. Equine Pract. 2021, 37, 461–494. [Google Scholar] [CrossRef]
- Portmann, A.-C.; Fournier, C.; Gimonet, J.; Ngom-Bru, C.; Barretto, C.; Baert, L. A Validation Approach of an End-to-End Whole Genome Sequencing Workflow for Source Tracking of Listeria Monocytogenes and Salmonella Enterica. Front. Microbiol. 2018, 9, 446. [Google Scholar] [CrossRef]
- Cito, F.; Di Pasquale, A.; Cammà, C.; Cito, P. The Italian Information System for the Collection and Analysis of Complete Genome Sequence of Pathogens Isolated from Animal, Food and Environment. Int. J. Infect. Dis. 2018, 73, 296–297. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Lukjancenko, O.; Saputra, D.; Rasmussen, S.; Hasman, H.; Sicheritz-Pontén, T.; Aarestrup, F.M.; Ussery, D.W.; Lund, O. Benchmarking of Methods for Genomic Taxonomy. J. Clin. Microbiol. 2014, 52, 1529–1539. [Google Scholar] [CrossRef] [PubMed]
- Florensa, A.F.; Kaas, R.S.; Clausen, P.T.L.C.; Aytan-Aktug, D.; Aarestrup, F.M. ResFinder—An Open Online Resource for Identification of Antimicrobial Resistance Genes in next-Generation Sequencing Data and Prediction of Phenotypes from Genotypes. Microb. Genom. 2022, 8, 000748. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A Better, Faster Version of the PHAST Phage Search Tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.; Pettengill, J.B.; Luo, Y.; Payne, J.; Shpuntoff, A.; Rand, H.; Strain, E. CFSAN SNP Pipeline: An automated method for constructing SNP matrices from next-generation sequence data. PeerJ Comput. Sci. 2015, 1, e20. [Google Scholar] [CrossRef]
- Racklyeft, D.; Love, D. Bacterial Infection of the Lower Respiratory Tract in 34 Horses. Aust. Vet. J. 2000, 78, 549–559. [Google Scholar] [CrossRef]
- Laing, G.; Christley, R.; Stringer, A.; Ashine, T.; Cian, F.; Aklilu, N.; Newton, R.; Radford, A.; Pinchbeck, G. Pathology, Infectious Agents and Horse- and Management-Level Risk Factors Associated with Signs of Respiratory Disease in Ethiopian Working Horses. Equine Vet. J. 2021, 53, 670–681. [Google Scholar] [CrossRef]
- Kuchipudi, S.V.; Surendran Nair, M.; Yon, M.; Gontu, A.; Nissly, R.H.; Barry, R.; Greenawalt, D.; Pierre, T.; Li, L.; Thirumalapura, N.; et al. A Novel Real-Time PCR Assay for the Rapid Detection of Virulent Streptococcus Equi Subspecies Zooepidemicus—An Emerging Pathogen of Swine. Front. Vet. Sci. 2021, 8, 604675. [Google Scholar] [CrossRef]
- Steward, K.F.; Robinson, C.; Holden, M.T.G.; Harris, S.R.; Ros, A.F.; Pérez, G.C.; Baselga, R.; Waller, A.S. Diversity of Streptococcus Equi Subsp. Zooepidemicus Strains Isolated from the Spanish Sheep and Goat Population and the Identification, Function and Prevalence of a Novel Arbutin Utilisation System. Vet. Microbiol. 2017, 207, 231–238. [Google Scholar] [CrossRef]
- Pisoni, G.; Zadoks, R.N.; Vimercati, C.; Locatelli, C.; Zanoni, M.G.; Moroni, P. Epidemiological Investigation of Streptococcus Equi Subspecies Zooepidemicus Involved in Clinical Mastitis in Dairy Goats. J. Dairy Sci. 2009, 92, 943–951. [Google Scholar] [CrossRef]
- Hasegawa, T.; Torii, K.; Hashikawa, S.; Iinuma, Y.; Ohta, M. Cloning and Characterization of Two Novel DNases from Streptococcus pyogenes. Arch. Microbiol. 2002, 177, 451–456. [Google Scholar] [CrossRef]
- Meurer, M.; Öhlmann, S.; Bonilla, M.C.; Valentin-Weigand, P.; Beineke, A.; Hennig-Pauka, I.; Schwerk, C.; Schroten, H.; Baums, C.G.; von Köckritz-Blickwede, M.; et al. Role of Bacterial and Host DNases on Host-Pathogen Interaction during Streptococcus Suis Meningitis. Int. J. Mol. Sci. 2020, 21, 5289. [Google Scholar] [CrossRef] [PubMed]
- Waller, A.S.; Robinson, C. Streptococcus Zooepidemicus and Streptococcus Equi Evolution: The Role of CRISPRs. Biochem. Soc. Trans. 2013, 41, 1437–1443. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, R.; Artiushin, S.; Timoney, J.F. P9, a Temperate Bacteriophage of Streptococcus Equi. Int. Congr. Ser. 2006, 1289, 165–168. [Google Scholar] [CrossRef]
- McShan, W.M.; McCullor, K.A.; Nguyen, S.V. The Bacteriophages of Streptococcus pyogenes. Microbiol. Spectr. 2019, 7, 3. [Google Scholar] [CrossRef]
- Nocera, F.P.; D’Eletto, E.; Ambrosio, M.; Fiorito, F.; Pagnini, U.; De Martino, L. Occurrence and Antimicrobial Susceptibility Profiles of Streptococcus Equi Subsp. Zooepidemicus Strains Isolated from Mares with Fertility Problems. Antibiotics 2021, 11, 25. [Google Scholar] [CrossRef]
- Firew, S.; Pal, M. Clinical and Microbiological Observations on Strangles in Donkeys. Haryana Vet. 2015, 54, 64–66. [Google Scholar]
- Dong, J.; Gao, N.; Waller, A.S.; Cook, F.R.; Fan, S.; Yuan, D.; Du, Y.; Li, F.; Norimine, J.; Zhu, W. An Outbreak of Strangles Associated with a Novel Genotype of Streptococcus Equi Subspecies Equi in Donkeys in China during 2018. Equine Vet. J. 2019, 51, 743–748. [Google Scholar] [CrossRef]
- Zhu, Y.; Chen, S.; Yi, Z.; Holyoak, R.; Wang, T.; Ding, Z.; Li, J. Nasopharyngeal Microbiomes in Donkeys Shedding Streptococcus Equi Subspecies Equi in Comparison to Healthy Donkeys. Front. Vet. Sci. 2021, 8, 645627. [Google Scholar] [CrossRef]
- Allen, G.P.; Kydd, J.H.; Slater, J.D.; Smith, K.C. Advances in Understanding of the Pathogenesis, Epidemiology, and Immunological Control of Equid Herpesvirus Abortion. In Infectious Diseases VIII, Proceedings of the Eighth International Conference, Dubai, United Arab Emirates, 23–26 March 1998; R & W Publications: Suffolk, UK, 1999. [Google Scholar]
- Rickards, K.J.; Thiemann, A.K. Respiratory Disorders of the Donkey. Vet. Clin. N. Am. Equine Pract. 2019, 35, 561–573. [Google Scholar] [CrossRef]
- Burden, F.; Thiemann, A. Donkeys Are Different. J. Equine Vet. Sci. 2015, 35, 376–382. [Google Scholar] [CrossRef]
- Thiemann, A. Clinical Approach to the Dull Donkey. Practice 2013, 35, 470–476. [Google Scholar] [CrossRef]
- Oliveira, F.G.; Cook, R.F.; Naves, J.H.F.; Oliveira, C.H.S.; Diniz, R.S.; Freitas, F.J.C.; Lima, J.M.; Sakamoto, S.M.; Leite, R.C.; Issel, C.J.; et al. Equine Infectious Anemia Prevalence in Feral Donkeys from Northeast Brazil. Prev. Vet. Med. 2017, 140, 30–37. [Google Scholar] [CrossRef]
- Jerele, S.; Davis, E.; Mapes, S.; Pusterla, N.; Navas González, F.J.; Iglesias Pastrana, C.; Abdelfattah, E.M.; McLean, A. Survey of Serum Amyloid A and Bacterial and Viral Frequency Using QPCR Levels in Recently Captured Feral Donkeys from Death Valley National Park (California). Animals 2020, 10, 1086. [Google Scholar] [CrossRef] [PubMed]
- Osthaus, B.; Proops, L.; Long, S.; Bell, N.; Hayday, K.; Burden, F. Hair Coat Properties of Donkeys, Mules and Horses in a Temperate Climate. Equine Vet. J. 2018, 50, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Avadhanula, V.; Rodriguez, C.A.; Devincenzo, J.P.; Wang, Y.; Webby, R.J.; Ulett, G.C.; Adderson, E.E. Respiratory Viruses Augment the Adhesion of Bacterial Pathogens to Respiratory Epithelium in a Viral Species- and Cell Type-Dependent Manner. J. Virol. 2006, 80, 1629–1636. [Google Scholar] [CrossRef] [PubMed]
- Timoney, J.F. The Pathogenic Equine Streptococci. Vet. Res. 2004, 35, 397–409. [Google Scholar] [CrossRef]


{kind=link}
| Antibiotic | Results |
|---|---|
| Ceftiofur | S |
| Tylosin | S |
| Rifampicin | S |
| Ampicillin | S |
| Lincomycin | S |
| Rifamixin | I |
| Spiramycin | I |
| Penicillin | I |
| Gentamicin | I |
| Neomycin | R |
| Amikacin | R |
| Sulfamethoxazole and Trimethoprim | R |
| Amoxicillin | R |
| Tetracycline | R |
| Streptomycin | R |
| Aminosidine | R |
| Case n° | Isolate ID | Matrix | Virulence-Associated Genes | PHASTER | ResFinder |
|---|---|---|---|---|---|
| 1 | 2022.TE.33840.1.2 | Lung | - | - | - |
| 1 | 2022.TE.33841.1.2 | Liver | mf2 | - | - |
| 1 | 2022.TE.33842.1.2 | Spleen | mf2 | - | - |
| 2 | 2023.TE.1927.1.2 | Retropharyngeal lymph nodes | mf2 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cantelmi, M.C.; Merola, C.; Averaimo, D.; Chiaverini, A.; Cito, F.; Cocco, A.; Di Teodoro, G.; De Angelis, M.E.; Di Bernardo, D.; Auzino, D.; et al. Identification of the Novel Streptococcus equi subsp. zooepidemicus Sequence Type 525 in Donkeys of Abruzzo Region, Italy. Pathogens 2023, 12, 750. https://doi.org/10.3390/pathogens12060750
Cantelmi MC, Merola C, Averaimo D, Chiaverini A, Cito F, Cocco A, Di Teodoro G, De Angelis ME, Di Bernardo D, Auzino D, et al. Identification of the Novel Streptococcus equi subsp. zooepidemicus Sequence Type 525 in Donkeys of Abruzzo Region, Italy. Pathogens. 2023; 12(6):750. https://doi.org/10.3390/pathogens12060750
Chicago/Turabian StyleCantelmi, Maria Chiara, Carmine Merola, Daniela Averaimo, Alexandra Chiaverini, Francesca Cito, Antonio Cocco, Giovanni Di Teodoro, Maria Elisabetta De Angelis, Daniela Di Bernardo, Davide Auzino, and et al. 2023. "Identification of the Novel Streptococcus equi subsp. zooepidemicus Sequence Type 525 in Donkeys of Abruzzo Region, Italy" Pathogens 12, no. 6: 750. https://doi.org/10.3390/pathogens12060750
APA StyleCantelmi, M. C., Merola, C., Averaimo, D., Chiaverini, A., Cito, F., Cocco, A., Di Teodoro, G., De Angelis, M. E., Di Bernardo, D., Auzino, D., & Petrini, A. (2023). Identification of the Novel Streptococcus equi subsp. zooepidemicus Sequence Type 525 in Donkeys of Abruzzo Region, Italy. Pathogens, 12(6), 750. https://doi.org/10.3390/pathogens12060750