
Differential Protein Expression of Taenia crassiceps ORF Strain in the Murine Cysticercosis Model Using Resistant (C57BL/6) Mice

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Resistant Mice Infection with the T. crassiceps Cysticerci ORF Strain
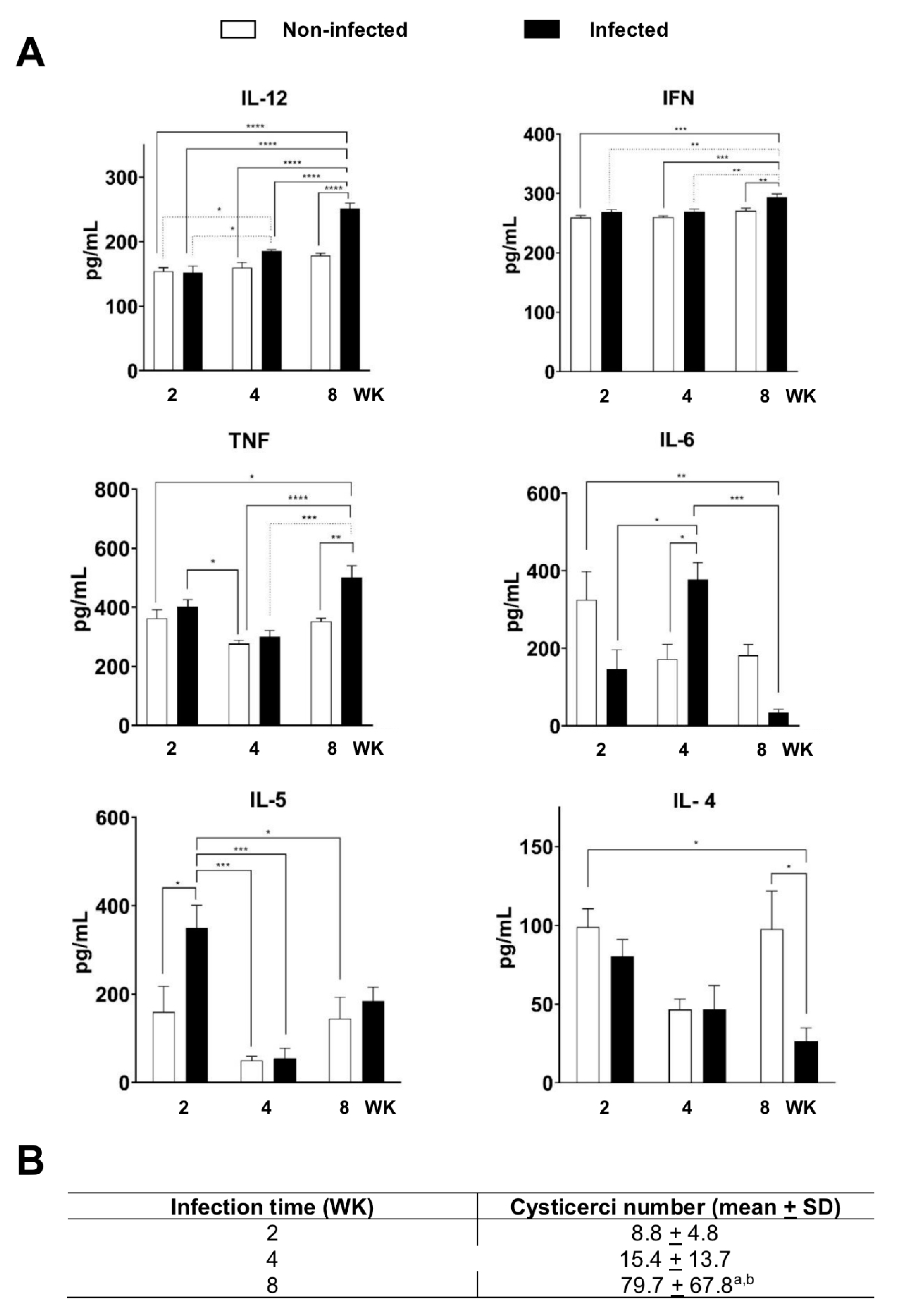
2.3. Spleen Cells Isolation and Cytokine Detection
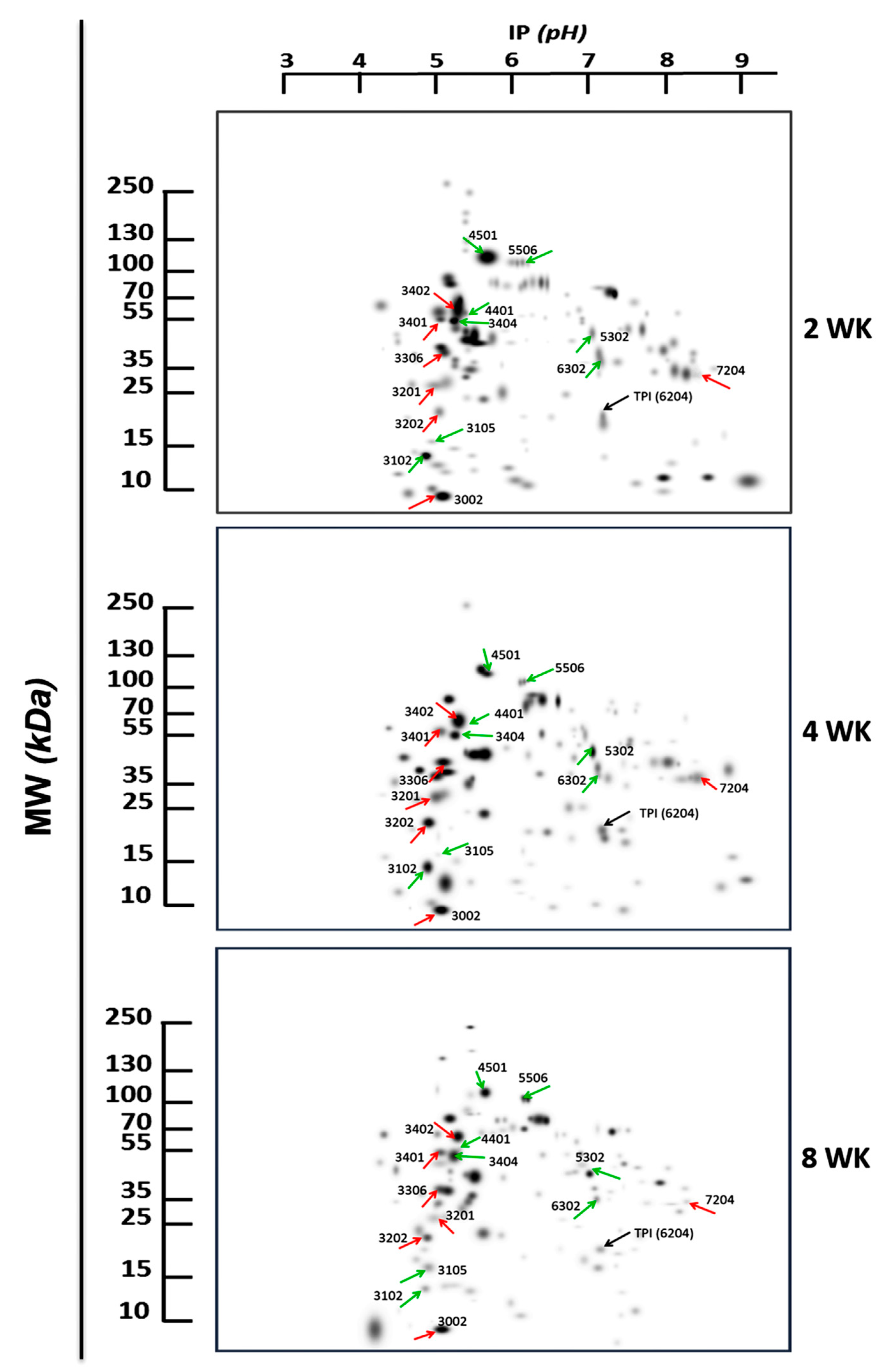
2.4. Two-Dimensional Gel Electrophoresis (2DE)
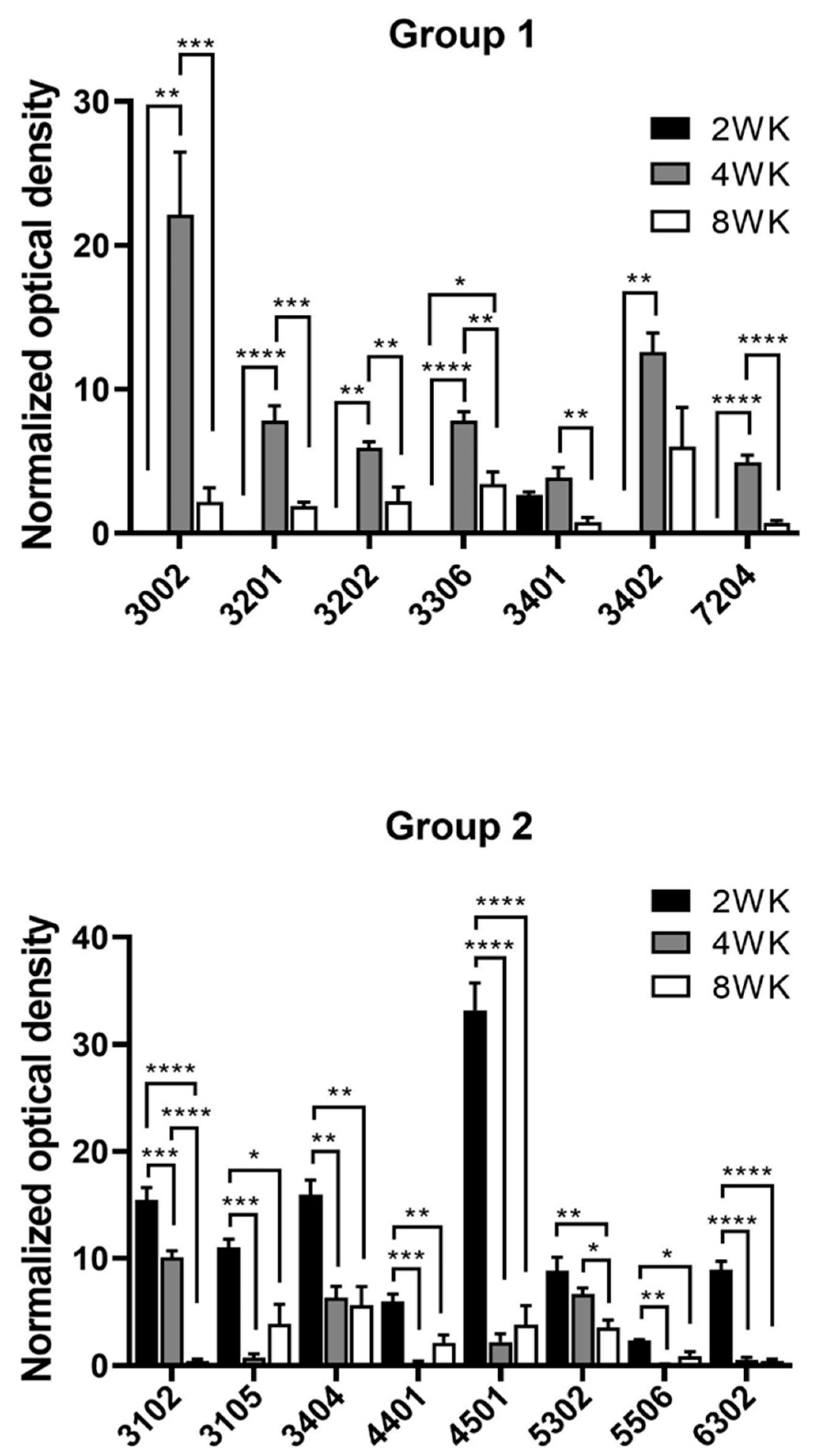
2.5. Proteomics Analysis
2.6. Statistical Analysis
2.7. MALDI-TOF Mass Spectrometry Protein-Identification
3. Results
3.1. Immune Response in the T. crassiceps Cysticercosis Model in C57BL/6 Mouse
3.2. Proteomics Analysis and Mass Spectrometry Protein-Identification
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Working to Overcome the Global Impact of Neglected Tropical Diseases: First WHO Report on Neglected Tropical Diseases; World Health Organization: Geneva, Switzerland, 2010; Available online: https://apps.who.int/iris/handle/10665/44440 (accessed on 1 January 2023).
- Vaz, A.J.; Nunes, C.M.; Piazza, R.M.; Livramento, J.A.; Da Silva, M.V.; Nakamura, P.M.; Ferreira, A.W. Immunoblot with cerebrospinal fluid from patients with neuro-cysticercosis using antigen from cysticerci of Taenia solium and Taenia crassiceps. Am. J. Trop. Med. Hyg. 1997, 57, 354–357. [Google Scholar] [CrossRef]
- Willms, R.; Zurabian, R. Taenia crassiceps: In vivo and in vitro models. Para-sitology 2010, 137, 335–346. [Google Scholar] [CrossRef]
- Bobes, R.J.; Estrada, K.; Rios-Valencia, D.G.; Calderón-Gallegos, A.; de la Torre, P.; Carrero, J.C.; Sanchez-Flores, A.; Laclette, J.P. The genomes of two strains of Taenia crassiceps the animal model for the study of human cysticercosis. Front. Cell. Infect. Microbiol. 2022, 12, 876839. [Google Scholar] [CrossRef] [PubMed]
- Fragoso, G.; Lamoyi, E.; Mellor, A.; Lomelí, C.; Hernández, M.; Sciutto, E. Increased resistance to Taenia crassiceps murine cysticercosis in Qa-2 transgenic mice. Infect. Immun. 1998, 66, 760–764. [Google Scholar] [CrossRef]
- Togno-Peirce, C.; Nava-Castro, K.; Terrazas, L.I.; Morales-Montor, J. Sex-associated expression of co-stimulatory molecules CD80, CD86, and accessory molecules, PDL-1, PDL-2 and MHC-II, in F480+ macrophages during murine cysticercosis. BioMed Res. Int. 2013, 2013, 570158. [Google Scholar] [CrossRef] [PubMed]
- Adalid-Peralta, L.; Lopez-Roblero, A.; Camacho-Vázquez, C.; Nájera-Ocampo, M.; Guevara-Salinas, A.; Ruiz-Monroy, N.; Melo-Salas, M.; Morales-Ruiz, V.; López-Recinos, D.; Ortiz-Hernández, E.; et al. Regulatory T cells as an escape mecanismo to the inmune response in Taenia crassiceps infección. Front. Cell. Infect. Microbiol. 2021, 11, 630583. [Google Scholar] [CrossRef]
- Dissanayake, S.; Amith, R.S.; Shahin, A. Taenia crassiceps carbohydrates stimulate IL-6 expression in naïve murine macrophages via Toll-like receptors (TLRs). Mol. Immunol. 2004, 41, 391–398. [Google Scholar] [CrossRef]
- Reyes, J.L.; Terrazas, C.A.; Vera-Arias, L.; Terrazas, L.I. Differential response of antigen presenting cells from susceptible and resistant strains of mice to Taenia crassiceps infection. Infect. Genet. Evol. 2009, 9, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Freitas, A.A.; Moura, V.B.L.; Gonçalves, S.F.; Rodrigues, A.A.; Félix, R.M.; Soares, T.P.; Irusta, V.C.R.; Vinaud, M.C.; Oliveira, M.A.P.; Lino-Junior, R.S. Kinetics of the inflammatory response in subcutaneous cysticercosis induced in mice by Taenia crassiceps. J. Comp. Pathol. 2012, 147, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, E.B.; Sakai, Y.I.; De Gaspari, E. A mouse air pouch model for evaluating the immune response to Taenia crassiceps infection. Exp. Parasitol. 2014, 137, 66–73. [Google Scholar] [CrossRef]
- Spolski, R.S.; Corson, J.; Thomas, P.G.; Kuhn, R.E. Parasite-secreted products regulate the host response to larval Taenia crassiceps. Parasite Immunol. 2000, 22, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Gómez-García, L.; Rivera-Montoya, I.; Rodríguez-Sosa, M.; Terrazas, L.I. Carbohydrate components of Taenia crassiceps metacestodes display Th2-adjuvant and anti-inflammatory properties when co-injected with bystander antigen. Parasitol. Res. 2006, 99, 440–448. [Google Scholar] [CrossRef]
- Peón, A.N.; Espinoza-Jiménez, A.; Terrazas, L.I. Immunoregulation by Taenia crassiceps and its antigens. BioMed Res. Int. 2013, 2013, 498583. [Google Scholar] [CrossRef]
- Toledo, A.; Fragoso, G.; Rosas, G.; Hernández, M.; Gevorkian, G.; López-Casillas, F.; Hernández, B.; Acero, G.; Huerta, M.; Larralde, C.; et al. Two epitopes shared by Taenia crassiceps and Taenia solium confer protection against murine T. crassiceps cysticercosis along with a prominent T1 response. Infect. Immun. 2001, 69, 1766–1773. [Google Scholar] [CrossRef]
- Vega-Angeles, V.; Terrazas, L.; Ledesma-Soto, Y.; Jiménez, L.; Landa, A. Taenia solium glutathione transferase fraction activates macrophages and favors the development of Th1-type response. Biosci. Rep. 2019, 39, BSR20181132. [Google Scholar] [CrossRef] [PubMed]
- Vaca-Paniagua, F.; Torres-Rivera, A.; Parra-Unda, R.; Landa, A. Taenia solium: Antioxidant metabolism enzymes as targets for cestocidal drugs and vaccines. Curr. Top. Med. Chem. 2008, 8, 393–399. [Google Scholar] [CrossRef]
- Vibanco-Pérez, N.; Jiménez, L.; Mendoza-Hernández, G.; Landa, A. Characterization of a recombinant mu-class glutathione S-transferase from Taenia solium. Parasitol. Res. 2022, 88, 398–404. [Google Scholar]
- Díaz-Zaragoza, M.; Jiménez, L.; Hernández, M.; Hernández-Ávila, R.; Navarro, L.; Ochoa-Sánchez, A.; Encarnación-Guevara, S.; Ostoa-Saloma, P.; Landa, A. Protein expression profile of Taenia crassiceps cysticerci related to Th1- and Th2-type responses in the mouse cysticercosis model. Acta Trop. 2020, 212, 105696. [Google Scholar] [CrossRef] [PubMed]
- Kruger, N.J. The Bradford method for protein quantitation. In The Protein Protocols Handbook. Springer Protocols Handbooks; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2009. [Google Scholar] [CrossRef]
- Villarreal, J.M.; Becerra-Lobato, N.; Rebollar-Flores, J.E.; Medina-Aparicio, L.; Carbajal-Gómez, E.; Zavala-García, M.L.; Vázquez, A.; Gutiérrez-Ríos, R.M.; Olvera, L.; Encarnación, S.; et al. The Salmonella enterica serovar Typhi ltrR-ompR-ompC-ompF genes are involved in resistance to the bile salt sodium deoxycholate and in bacterial transformation. Mol. Microbiol. 2014, 92, 1005–1024. [Google Scholar] [CrossRef]
- Aitken, A.; Collinge, D.B.; van Heusden, B.P.; Isobe, T.; Roseboom, P.H.; Rosenfeld, G.; Soll, J. 14-3-3 proteins: A highly conserved, widespread family of eukaryotic proteins. Trends Biochem. Sci. 1992, 17, 498–501. [Google Scholar] [CrossRef]
- Jones, D.H.; Ley, S.; Aitken, A. Isoforms of 14-3-3 protein can form homo- and heterodimers in vivo and in vitro: Implications for function as adapter proteins. FEBS Lett. 1995, 368, 55. [Google Scholar] [CrossRef] [PubMed]
- McGonigle SMelissa JBeall, M.J.; Pearce, E.J. Eukaryotic initiation factor 2a subunit associates with TGFb receptors and 14-3-31 and acts as a modulator of the TGFb response. Biochemistry 2002, 41, 579587. [Google Scholar] [CrossRef]
- Obsilova, V.; Silhan, J.; Boura, E.; Teisinger, J.; Obsil, T. 14-3-3 proteins: A family of versatile molecular regulators. Physiol. Res. 2008, 57 (Suppl. S3), S11–S21. [Google Scholar] [CrossRef]
- Ye, Y.; Yang, Y.; Cai, X.; Liu, L.; Wu, K.; Yu, M. Down-regulation of 14-3-3 Zeta inhibits TGF-β1–induced actomyosin contraction in human trabecular meshwork cells through RhoA signaling pathway. Investig. Ophthalmol. Vis. Sci. 2016, 57, 719–730. [Google Scholar] [CrossRef]
- Siles-Lucas, M.; Merli, M.; Mackenstedt, U.; Gottstein, B. The Echinococcus multilocularis 14-3-3 protein protects mice against primary but not secondary alveolar Echinococcosis. Vaccine 2003, 21, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Virginio, V.G.; Monteiro, K.M.; Drumond, F.; de Carvalho, M.O.; Vargas, D.M.; Zaha, A.; Ferreira, H.B. Excretory/secretory products from in vitro-cultured Echinococcus granulosus protoscoleces. Mol. Biochem. Parasitol. 2012, 183, 15–22. [Google Scholar] [CrossRef]
- Park, I.; Han, C.; Jin, S.; Lee, B.; Choi, H.; Kwon, J.T.; Kim, D.; Kim, J.; Lifirsu, E.; Park, W.J.; et al. Myosin regulatory light chains are required to maintain the stability of myosin II and cellular integrity. Biochem. J. 2011, 434, 171–180. [Google Scholar] [CrossRef]
- Buffoni, L.; Piva, M.M.; Baska, P.; Januszkiewicz, K.; Norbury, L.J.; Prior, K.C.; Dezen, D.; Silva, A.S.; Wedrychowicz, H.; Mendes, R.E. Immunization with the recombinant myosin regulatory light chain (FhrMRLC) in Adjuplex® adjuvant elicits a Th1-biased immune response and a reduction of parasite burden in Fasciola hepatica infected rats. Parasitol. Int. 2020, 75, 102037. [Google Scholar] [CrossRef]
- Gnanasekar, M.; Salunkhe, A.M.; Mallia, A.K.; He, Y.X.; Kalyanasundaram, R. Praziquantel affects the regulatory myosin light chain of Schistosoma mansoni. Antimicrob. Agents Chemother 2009, 53, 1054–1060. [Google Scholar] [CrossRef]
- Yokoyama, M.; Kimura, M.Y.; Ito, T.; Hayashizaki, K.; Endo, Y.; Wang, Y.; Yagi, R.; Nakagawa, T.; Kato, N.; Matsubara, H.; et al. Myosin Light Chain 9/12 regulates the pathogenesis of inflammatory Bowel disease. Front. Immunol. 2021, 11, 594297. [Google Scholar] [CrossRef]
- Zhang, S.; Yu, M.; Guo, Q.; Li, R.; Li, G.; Tan, S.; Li, X.; Wei, Y.; Wu, M. Annexin A2 binds to endosomes and negatively regulates TLR4-triggered inflammatory responses via the TRAM-TRIF pathway. Sci. Rep. 2015, 5, 15859. [Google Scholar] [CrossRef]
- Liu, N.; Jiang, Y.; Chung, J.Y.; Li, Y.; Yu, Z.; Kim, J.W.; Lok, J.M.; Whalen, M.J.; Wang, X. Annexin A2 deficiency exacerbates neuroinflammation and long-term neurological deficits after traumatic brain injury in mice. Int. J. Mol. Sci. 2019, 20, 6125. [Google Scholar] [CrossRef] [PubMed]
- Sønder, S.L.; Boye, T.L.; Tölle, R.; Dengjel, J.; Maeda, K.; Jäättelä, M.; Simonsen, A.C.; Jaiswal, J.K.; Nylandsted, J. Annexin A7 is required for ESCRT III-mediated plasma membrane repair. Sci. Rep. 2019, 9, 6726. [Google Scholar] [CrossRef] [PubMed]
- Ammendolia, D.A.; Bement, W.M.; Brumell, J.H. Plasma membrane integrity: Implications for health and disease. BMC Biol. 2021, 19, 71. [Google Scholar] [CrossRef]
- Hammond, J.W.; Cai, D.; Verhey, K.J. Tubulin modifications and their cellular functions. Curr. Opin. Cell Biol. 2008, 20, 71–76. [Google Scholar] [CrossRef]
- Márquez-Navarro, A.; Pérez-Reyes, A.; Zepeda-Rodríguez, A.; Reynoso-Ducoing, O.; Hernández-Campos, A.; Hernández-Luis, F.; Castillo, R.; Yépez-Mulia, L.; Ambrosio, J.R. RCB20, an experimental benzimidazole derivative, affects tubulin expression and induces gross anatomical changes in Taenia crassiceps cysticerci. Parasitol. Res. 2013, 112, 2215–2226. [Google Scholar] [CrossRef]
- Reynoso-Ducoing, O.; Valverde-Islas, L.; Paredes-Salomon, C.; Pérez-Reyes, A.; Landa, A.; Robert, L.; Mendoza, G.; Ambrosio, J.R. Analysis of the expression of cytoskeletal proteins of Taenia crassiceps ORF strain cysticerci (Cestoda). Parasitol. Res. 2014, 113, 1955–1969. [Google Scholar] [CrossRef]
- De Lima, N.F.; Picanço, G.A.; Valencia, D.G.R.; Villegas, E.O.L.; Mellado, M.D.R.E.; Ambrosio, J.R.; Vinaud, M.C. Alterations in Taenia crassiceps cysticerci cytoskeleton induced by nitazoxanide and flubendazole. Acta Trop. 2021, 221, 106027. [Google Scholar] [CrossRef] [PubMed]
- Kinnunen, A.; Surcel, H.M.; Halttunen, M.; Tiitinen, A.; Morrison, R.P.; Morrison, S.G.; Koskela, P.; Lehtinen, M.; Paavonen, J. Chlamydia trachomatis heat shock protein-60 induced interferon-gamma and interleukin-10 production in infertile women: Chlamydial HSP60-induced cytokines in infertile women. Clin. Exp. Immunol. 2003, 131, 299–303. [Google Scholar] [CrossRef]
- Zanin-Zhorov, A.; Bruck, R.; Tal, G.; Oren, S.; Aeed, H.; Hershkoviz, R.; Cohen, I.R.; Lider, O. Heat shock protein 60 inhibits Th1-mediated hepatitis model via innate regulation of Th1/Th2 transcription factors and cytokines. J. Immunol. 2005, 174, 3227–3236. [Google Scholar] [CrossRef]
- Zhou, S.; Jin, X.; Chen, X.; Zhu, J.; Xu, Z.; Wang, X.; Liu, F.; Hu, W.; Zhou, L.; Su, C. Heat shock protein 60 in eggs specifically induces Tregs and reduces liver Immunopathology in mice with Schistosomiasis Japonica. PLoS ONE 2015, 10, e0139133. [Google Scholar] [CrossRef]
- Zhao, Y.; Coulson, E.J.; Su, X.; Zhang, J.; Sha, B.; Xu, H.; Deng, Y.; Chen, Y.; Cao, J.; Wang, Y.; et al. Identification of 14-3-3 epsilon as a regulator of the neural apoptotic pathway for chronic-stress-induced depression. iScience 2021, 24, 102043. [Google Scholar] [CrossRef]
- Fu, W.; Hu, W.; Yi, Y.S.; Hettinghouse, A.; Sun, G.; Bi, Y.; He, W.; Zhang, L.; Gao, G.; Liu, J.; et al. TNFR2/14-3-3ε signaling complex instructs macrophage plasticity in inflammation and autoimmunity. J. Clin. Investig. 2021, 131, e144016. [Google Scholar] [CrossRef] [PubMed]
- Köhnke, R.; Mei, J.; Park, M.; York, D.A.; Erlanson-Albertsson, C. Fatty acids and glucose in high concentration down-regulates ATP synthase beta-subunit protein expression in INS-1 cells. Nutr. Neurosci. 2007, 10, 273–278. [Google Scholar] [CrossRef]
- Santivañez, S.J.; Hernández-González, A.; Chile, N.; Oleaga, A.; Arana, Y.; Palma, S.; Verastegui, M.; Gonzalez, A.E.; Gilman, R.; Garcia, H.H.; et al. Proteomic study of activated Taenia solium oncospheres. Mol. Biochem. Parasitol. 2010, 171, 32–39. [Google Scholar] [CrossRef]
- Grzelak, S.; Stachyra, A.; Bień-Kalinowska, J. The first analysis of Trichinella spiralis and Trichinella britovi adult worm excretory-secretory proteins by two-dimensional electrophoresis coupled with LC-MS/MS. Vet. Parasitol. 2021, 297, 109096. [Google Scholar] [CrossRef]
- Reamtong, O.; Rujimongkon, K.; Sookrung, N.; Saeung, A.; Thiangtrongjit, T.; Sakol-varee, Y.; Thammapalo, S.; Loymek, S.; Chaicumpa, W. Immunome and immune complex-forming components of Brugia malayi identified by microfilaremic human sera. Exp. Parasitol. 2019, 200, 92–98. [Google Scholar] [CrossRef]
- Stolf, B.S.; Smyrnias, I.; Lopes, L.R.; Vendramin, A.; Goto, H.; Laurindo, F.R.M.; Shah, A.M.; Santos, C.X.C. Protein disulfide isomerase and host-pathogen interaction. Sci. World J. 2011, 11, 1749–1761. [Google Scholar] [CrossRef]
- Meng, Y.; Zhang, Q.; Zhang, M.; Gu, B.; Huang, G.; Wang, Q.; Shan, W. The protein disulfide isomerase 1 of Phytophthora parasitica (PpPDI1) is associated with the haustoria-like structures and contributes to plant infection. Front. Plant Sci. 2015, 6, 632. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, D.M.; Söling, H.D. The protein disulphide-isomerase family: Unravelling a string of folds. Biochem. J. 1999, 339 Pt 1, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Benhnini, F.; Chenik, M.; Laouini, D.; Louzir, H.; Cazenave, P.A.; Dellagi, K. Comparative evaluation of two vaccine candidates against experimental leishmaniasis due to Leishmania major infection in four inbred mouse strains. Clin. Vaccine Immunoly 2009, 16, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Y.; Chen, D.; Ji, S.; Yang, L.T.; Huang, Q.; Guan, I.; Chang, K.; Li, D.; Yuan, R.; et al. Dust-mite-derived protein disulfide isomerase suppresses airway allergy by inducing tolerogenic dendritic cells. J. Biol. Chem. 2021, 296, 100585. [Google Scholar] [CrossRef]
- Kalinna, B.H.; McManus, D.P. A vaccine against the Asian schistosome, Schistosoma japonicum: An update on paramyosin as a target of protective immunity. Int. J. Parasitol. 1997, 27, 1213–1219. [Google Scholar] [CrossRef]
- Vázquez-Talavera, J.; Solís, C.F.; Terrazas, L.I.; Laclette, J.P. Characterization and protective potential of the immune response to Taenia solium paramyosin in a murine model of cysticercosis. Infect. Immun. 2001, 69, 5412–5416. [Google Scholar] [CrossRef]
- Hao, C.; Wang, W.; Zhan, B.; Wang, Z.; Huang, J.; Sun, X.; Zhu, X. Trichinella spiralis Paramyosin Induces Colonic Regulatory T Cells to Mitigate Inflammatory Bowel Disease. Front. Cell Dev. Biol. 2021, 9, 695015. [Google Scholar] [CrossRef] [PubMed]
- Laclette, J.P.; Shoemaker, C.B.; Richter, D.; Arcos, L.; Pante, N.; Cohen, C.; Bing, D.; Nicholson-Weller, A. Paramyosin inhibits complement C1. J. Immunol. 1992, 148, 124–128. [Google Scholar] [CrossRef]
- Sun, R.; Zhao, X.; Wang, Z.; Yang, J.; Zhao, L.; Zhan, B.; Zhu, X. Trichinella spiralis paramyosin binds human complement C1q and inhibits classical complement activation. PLoS Negl. Trop. Dis. 2015, 9, e0004310. [Google Scholar] [CrossRef]
- Pancholi, V. Multifunctional α-enolase: Its role in diseases. Cell. Mol. Life Sci. 2001, 58, 902–920. [Google Scholar] [CrossRef] [PubMed]
- Jolodar, A.; Fischer, P.; Bergmann, S.; Buttner, D.W.; Hammerschmidt, S.; Brattig, N.W. Molecular cloning of an alpha-enolase from the human filarial parasite Onchocerca volvulus that binds human plasminogen. Biochim. Biophys. Acta 2003, 1627, 111–120. [Google Scholar] [CrossRef]
- Bernal, D.; de la Rubia, J.E.; Carrasco-Abad, A.M.; Toledo, R.; Mas-Coma, S.; Marcilla, A. Identification of enolase as a plasminogen-binding protein in excretory-secretory products of Fasciola hepatica. FEBS Lett. 2004, 563, 203–206. [Google Scholar] [CrossRef]
- Gan, W.; Zhao, G.; Xu, H.; Wu, W.; Du, W.; Huang, J.; Yu, X.; Hu, X. Reverse vaccinology approach identifies an Echinococcus granulosus tegumental membrane protein enolase as vaccine candidate. Parasitol. Res. 2010, 106, 873–882. [Google Scholar] [CrossRef]
- Wongkamchai, S.; Chiangjong, W.; Sinchaikul, S.; Chen, S.T.; Choochote, W.; Thongboonkerd, V. Identification of Brugia malayi immunogens by an immunoproteomics approach. J. Proteom. 2011, 74, 1607–1613. [Google Scholar] [CrossRef] [PubMed]
- Ayón-Núñez, D.A.; Fragoso, G.; Bobes, R.J.; Laclette, J.P. Plasminogen-binding proteins as an evasion mechanism of the host’s innate immunity in infectious diseases. Biosci. Rep. 2018, 38, BSR20180705. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; You, Y.; Luo, X.; Zheng, Y.; Cai, X. Molecular and biochemical characterization of Taenia solium α-enolase. Vet. Parasitol. 2018, 254, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.; Yeom, J.; Wang, H.; Kim, S.; Ahn, C.; Kim, J.; Yang, H.; Kong, Y. Taenia solium metacestode fasciclin-like protein is reactive with sera of chronic neurocysticercosis. Trop. Med. Int. Health 2014, 19, 719–725. [Google Scholar] [CrossRef]
- Ahn, C.; Kim, J.; Bae, Y.; Kim, S.; Shin, J.; Yang, Y.; Kang, I.; Kong, Y. Fasciclin calcareous corpuscle binary complex mediated protein-protein interactions in Taenia solium metacestode. Parasites Vectors 2017, 10, 438. [Google Scholar] [CrossRef]
- Ahn, C.; Kim, J.; Huh, S.; Kang, I.; Kong, Y. Advances in serological diagnosis of Taenia solium neurocysticercosis in Korea. Genom. Inf. 2019, 17, e7. [Google Scholar] [CrossRef]
- Barthel, D.; Schindler, S.; Zipfel, P.F. Plasminogen is a complement inhibitor. J. Biol. Chem. 2012, 287, 18831–18842. [Google Scholar] [CrossRef]
- Espíndola, N.; De Gaspari, E.; Nakamura, P.; Vaz, A. Cross-reactivity of anti-Taenia crassiceps cysticerci immune antibodies with Taenia solium antigens. Vet. Parasitol. 2000, 89, 321–326. [Google Scholar] [CrossRef]
- Rodríguez-Sosa, M.; Rosas, L.E.; David, J.R.; Bojalil, R.; Satoskar, A.R.; Terrazas, L.I. Macrophage migration inhibitory factor plays a critical role in mediating protection against the helminth parasite Taenia crassiceps. Infect. Immun. 2003, 71, 1247–1254. [Google Scholar] [CrossRef]
- Sciutto, E.; Fragoso, G.; Baca, M.; De la Cruz, V.; Lemus, L.; Lamoyi, E. Depressed T-cell proliferation associated with susceptibility to experimental Taenia crassiceps infection. Infect. Immun. 1995, 63, 2277–2281. [Google Scholar] [CrossRef]
- Terrazas, L.I.; Bojalilt, R.; Govezensky, T.; Larralde, C. Shift from an Early Protective TH1-Type Immune Response to a Late Permissive TH2-Type Response in Murine Cysticercosis (Taenia crassiceps). J. Parasitol. 1998, 84, 74–81. [Google Scholar] [CrossRef]
- Toenjes, S.A.; Kuhn, R.E. The initial immune response during experimental cysticercosis is of the mixed Thl/Th2 type. Parasitol. Res. 2003, 89, 407–413. [Google Scholar] [CrossRef]
- Al-Sherbiny, M.; Osman, A.; Barakat, R.; El Morshedy, H.; Bergquist, R.; Olds, R. In vitro cellular and humoral responses to Schistosoma mansoni vaccine candidate antigens. Acta Trop. 2003, 88, 117–130. [Google Scholar] [CrossRef]
- Alvarez, J.I.; Londono, D.P.; Alvarez, A.L.; Trujillo, J.; Jaramillo, M.M.; Restrepo, B.I. Granuloma formation and parasite disintegration in porcine cysticercosis: Comparison with human neurocysticercosis. J. Comp. Pathol. 2002, 127, 186–193. [Google Scholar] [CrossRef]
- Mohammad, I.; Nousiainen, K.; Bhosale, S.D.; Starskaia, I.; Moulder, R.; Rokka, A.; Cheng, F.; Mohanasundaram, P.; Eriksson, J.E.; Goodlett, D.R.; et al. Quantitative proteomic characterization and comparison of T helper 17 and induced regulatory T cells. PLoS Biol. 2018, 16, e2004194. [Google Scholar] [CrossRef]
- Moura, V.B.; Lima, S.B.; Matos-Silva, H.; Vinaud, M.C.; Loyola, P.R.; Lino, R.S. Cellular immune response in intraventricular experimental neurocysticercosis. Parasitology 2016, 143, 334–342. [Google Scholar] [CrossRef]
- Vargas-Parada, L.; Laclette, J.P. Gene structure of Taenia solium paramyosin. Parasitol. Res. 2003, 89, 375–378. [Google Scholar] [CrossRef]
- Soleyman, N.M.; Darnhofer, B.; Birner-Gruenberger, R.; Abnous, K.; Borji, H. Proteomic analysis of soluble protein extract of adult Toxocara cati. Comparative Immunology. Microbiol. Infect. Dis. 2020, 73, 101528. [Google Scholar] [CrossRef]
- Dallacasagrande, V.; Hajjar, K.A. Annexin A2 in inflammation and host defense. Cells 2020, 9, 1499. [Google Scholar] [CrossRef]
- Larralde, C.; Morales, J.; Terrazas, L.I.; Govezensky, T.; Romano, M. Sex hormone changes induced by the parasite lead to feminization of the male host in murine Taenia crassiceps cysticercosis. J. Steroid Biochem. Mol. Biol. 1995, 52, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Escobedo, G.; Larralde, C.; Chavarria, A.; Cerbón, M.A.; Morales-Montor, J. Molecular mechanisms involved in the differential effects of sex steroids on the reproductions and infectivity of Taenia crassiceps. J. Parasitol. 2004, 90, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Terrazas, L.I.; Bojalil, R.; Rodríguez-Sosa, M.; Govesenky, T.; Larralde, C. Taenia crassiceps cysticercosis: A role for prostaglandin E2 in susceptibility. Parastol. Res. 1999, 85, 1025–1031. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Protein (Spot number) | MASCOT | Taenia solium PRJNA170813 | Theoretical data MW | Access to UniprotKB | |
|---|---|---|---|---|---|
| Score | Sequence coverage (%) | Contig/Gene ID | (kDa/pI (pH) | ||
| GROUP 1 | |||||
| 14-3-3 protein zeta/delta (3201) | 78 | 36 | 00616/TsM_000719200 | 28,069.73/4.63 | U6JEE0_ECHGR |
| Myosin regulatory light chain y (3202) | 94 | 49 | 00860/TsM_001153700 | 21,929.57/4.53 | A0A068YDE7_ ECHMU |
| Annexin B7 (3306) | 97 | 27 | 00183/TsM_000793200 | 111.55/4.39 | Q52MU2 _TAESO |
| Tubulin beta 2C chain (3401) | 112 | 19 | 01071/TsM_001072900 | 112.04/4.49 | A0A068WIF6_ ECHGR |
| Heat shock protein 60 kDa protein (3402) | 105 | 41 | 00260/TsM_000546800 | 60,568.63/5.09 | A3F4T7 _TAEAS |
| GROUP 2 | |||||
| 14-3-3 protein epsilon (3102) | 110 | 58 | 00306/TsM_000569000 | 29,217.94/4.70 | U6JEE0 _ECHGR |
| ATP synthase subunit beta (3404) | 104 | 34 | 00029/TsM_000805000 | 55,799.04/5.39 | A0A0R3VYB8 _TAEAS |
| Protein disulfide isomerase (4401) | 100 | 39 | 04289/TsM_000671300 | 55,214.79/4.61 | A0A068WY47_ECHGR |
| Paramyosin (4501) | 100 | 36 | 00569/TsM_001115200 | 107,772.95/5.00 | Q68J63_ TAESO |
| Enolase 3 (5302) | 121 | 42 | 02256/TsM_000447100 | 46,557.32/6.90 | A0A2P1AM74 _TAESO |
| Fasciclin-1 (5506) | 106 | 29 | 00312/TsM_000655200 | 95,014.71/6.66 | X2D553_TAESO |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez, L.; Díaz-Zaragoza, M.; Hernández, M.; Navarro, L.; Hernández-Ávila, R.; Encarnación-Guevara, S.; Ostoa-Saloma, P.; Landa, A. Differential Protein Expression of Taenia crassiceps ORF Strain in the Murine Cysticercosis Model Using Resistant (C57BL/6) Mice. Pathogens 2023, 12, 678. https://doi.org/10.3390/pathogens12050678
Jiménez L, Díaz-Zaragoza M, Hernández M, Navarro L, Hernández-Ávila R, Encarnación-Guevara S, Ostoa-Saloma P, Landa A. Differential Protein Expression of Taenia crassiceps ORF Strain in the Murine Cysticercosis Model Using Resistant (C57BL/6) Mice. Pathogens. 2023; 12(5):678. https://doi.org/10.3390/pathogens12050678
Chicago/Turabian StyleJiménez, Lucía, Mariana Díaz-Zaragoza, Magdalena Hernández, Luz Navarro, Ricardo Hernández-Ávila, Sergio Encarnación-Guevara, Pedro Ostoa-Saloma, and Abraham Landa. 2023. "Differential Protein Expression of Taenia crassiceps ORF Strain in the Murine Cysticercosis Model Using Resistant (C57BL/6) Mice" Pathogens 12, no. 5: 678. https://doi.org/10.3390/pathogens12050678
APA StyleJiménez, L., Díaz-Zaragoza, M., Hernández, M., Navarro, L., Hernández-Ávila, R., Encarnación-Guevara, S., Ostoa-Saloma, P., & Landa, A. (2023). Differential Protein Expression of Taenia crassiceps ORF Strain in the Murine Cysticercosis Model Using Resistant (C57BL/6) Mice. Pathogens, 12(5), 678. https://doi.org/10.3390/pathogens12050678