

Translational Fidelity during Bacterial Stresses and Host Interactions



Abstract
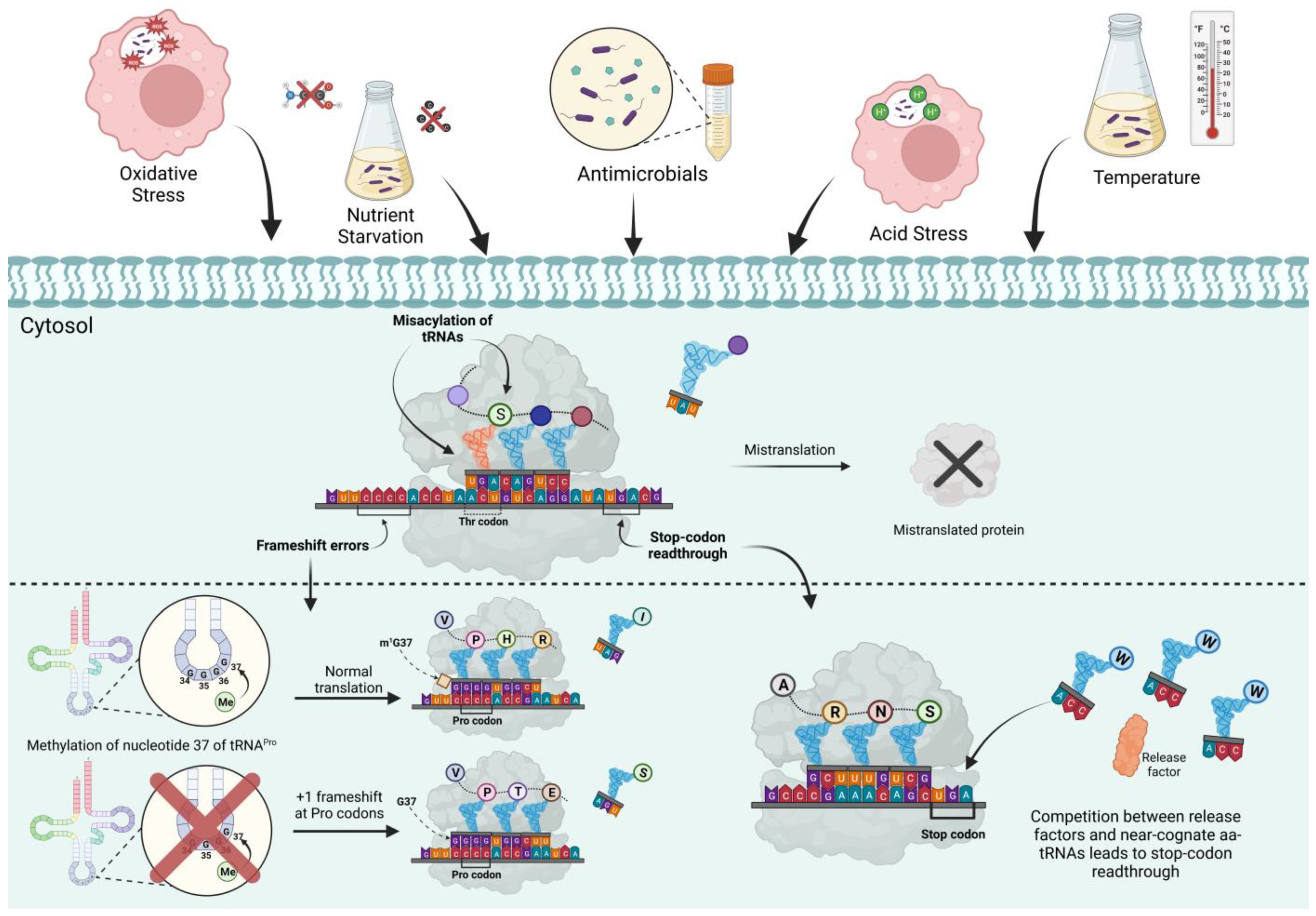
1. Introduction
2. Translational Fidelity during Bacterial Stresses
2.1. Effects of Oxidative Stress on Translational Fidelity
2.2. Effects of Metabolic Stresses on Translational Fidelity
2.3. Antibiotics Affecting Translational Fidelity
2.4. Translational Fidelity and Stress Resistance
3. Altered Translational Fidelity in Salmonella and Other Bacteria
3.1. Ribosomal Fidelity Mutations in Salmonella
3.2. Modification Defects of tRNAs in Bacterial Pathogens
3.3. Glu and Asp Misincorporation in Mycobacteria
3.4. Editing Defects in Host-Restricted Bacteria
3.5. Trans-Editing in Streptococci
3.6. Aminoglycoside-Induced Biofilm Formation in Pseudomonas aeruginosa
3.7. Mistranslating ProRS/tRNAPro in Streptomyces
4. Concluding Remarks and Future Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pontes, M.H.; Sevostyanova, A.; Groisman, E.A. When Too Much ATP Is Bad for Protein Synthesis. J. Mol. Biol. 2015, 427, 2586–2594. [Google Scholar] [CrossRef] [PubMed]
- Nissen, P.; Hansen, J.; Ban, N.; Moore, P.B.; Steitz, T.A. The structural basis of ribosome activity in peptide bond synthesis. Science 2000, 289, 920–930. [Google Scholar] [CrossRef] [PubMed]
- Ogle, J.M.; Ramakrishnan, V. Structural insights into translational fidelity. Annu. Rev. Biochem. 2005, 74, 129–177. [Google Scholar] [CrossRef] [PubMed]
- Pape, T.; Wintermeyer, W.; Rodnina, M. Induced fit in initial selection and proofreading of aminoacyl-tRNA on the ribosome. EMBO J. 1999, 18, 3800–3807. [Google Scholar] [CrossRef] [PubMed]
- Rodnina, M.V.; Wintermeyer, W. Ribosome fidelity: tRNA discrimination, proofreading and induced fit. Trends Biochem. Sci. 2001, 26, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Ibba, M.; Söll, D. Aminoacyl-tRNA synthesis. Annu. Rev. Biochem. 2000, 69, 617–650. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.; Reynolds, N.; Ibba, M. Aminoacyl-tRNA synthesis and translational quality control. Annu. Rev. Microbiol. 2009, 63, 61–78. [Google Scholar] [CrossRef]
- Mascarenhas, A.P.; An, S.; Rosen, A.E.; Martinis, S.A.; Musier-Forsyth, K. Fidelity mechanisms of the aminoacyl-tRNA synthetases. In Protein Engineering; RajBhandary, U.L., Köhrer, C., Eds.; Springer-Verlag: New York, NY, USA, 2008; pp. 153–200. [Google Scholar]
- Ahel, I.; Korencic, D.; Ibba, M.; Söll, D. Trans-editing of mischarged tRNAs. Proc. Natl. Acad. Sci. USA 2003, 100, 15422–15427. [Google Scholar] [CrossRef]
- Vargas-Rodriguez, O.; Musier-Forsyth, K. Exclusive use of trans-editing domains prevents proline mistranslation. J. Biol. Chem. 2013, 288, 14391–14399. [Google Scholar] [CrossRef]
- Mohler, K.; Ibba, M. Translational fidelity and mistranslation in the cellular response to stress. Nat. Microbiol. 2017, 2, 17117. [Google Scholar] [CrossRef]
- Evans, C.R.; Fan, Y.; Weiss, K.; Ling, J. Errors during gene expression: Single-cell heterogeneity, stress resistance, and microbe-host interactions. mBio 2018, 9, e01018-18. [Google Scholar] [CrossRef] [PubMed]
- Mohler, K.; Aerni, H.R.; Gassaway, B.; Ling, J.; Ibba, M.; Rinehart, J. MS-READ: Quantitative measurement of amino acid incorporation. Biochim. Biophys. Acta 2017, 1861, 3081–3088. [Google Scholar] [CrossRef] [PubMed]
- Wohlgemuth, I.; Garofalo, R.; Samatova, E.; Gunenc, A.N.; Lenz, C.; Urlaub, H.; Rodnina, M.V. Translation error clusters induced by aminoglycoside antibiotics. Nat. Commun. 2021, 12, 1830. [Google Scholar] [CrossRef]
- Zhang, H.; Lyu, Z.; Fan, Y.; Evans, C.R.; Barber, K.W.; Banerjee, K.; Igoshin, O.A.; Rinehart, J.; Ling, J. Metabolic stress promotes stop-codon readthrough and phenotypic heterogeneity. Proc. Natl. Acad. Sci. USA 2020, 117, 22167–22172. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Evans, C.R.; Barber, K.W.; Banerjee, K.; Weiss, K.J.; Margolin, W.; Igoshin, O.A.; Rinehart, J.; Ling, J. Heterogeneity of stop codon readthrough in single bacterial cells and implications for population fitness. Mol. Cell 2017, 67, 826–836. [Google Scholar] [CrossRef]
- Dunn, J.G.; Foo, C.K.; Belletier, N.G.; Gavis, E.R.; Weissman, J.S. Ribosome profiling reveals pervasive and regulated stop codon readthrough in Drosophila melanogaster. eLife 2013, 2, e01179. [Google Scholar] [CrossRef]
- Wangen, J.R.; Green, R. Stop codon context influences genome-wide stimulation of termination codon readthrough by aminoglycosides. eLife 2020, 9, e52611. [Google Scholar] [CrossRef]
- Kramer, E.B.; Vallabhaneni, H.; Mayer, L.M.; Farabaugh, P.J. A comprehensive analysis of translational missense errors in the yeast Saccharomyces cerevisiae. RNA 2010, 16, 1797–1808. [Google Scholar] [CrossRef]
- Kramer, E.B.; Farabaugh, P.J. The frequency of translational misreading errors in E. coli is largely determined by tRNA competition. RNA 2007, 13, 87–96. [Google Scholar] [CrossRef]
- Battesti, A.; Majdalani, N.; Gottesman, S. The RpoS-mediated general stress response in Escherichia coli. Annu. Rev. Microbiol. 2011, 65, 189–213. [Google Scholar] [CrossRef]
- Storz, G.; Imlay, J.A. Oxidative stress. Curr. Opin. Microbiol. 1999, 2, 188–194. [Google Scholar] [CrossRef]
- Fang, F.C.; Frawley, E.R.; Tapscott, T.; Vazquez-Torres, A. Bacterial stress responses during host infection. Cell Host Microbe 2016, 20, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Hampton, M.B.; Kettle, A.J.; Winterbourn, C.C. Inside the neutrophil phagosome: Oxidants, myeloperoxidase, and bacterial killing. Blood 1998, 92, 3007–3017. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; Hampton, M.B.; Livesey, J.H.; Kettle, A.J. Modeling the reactions of superoxide and myeloperoxidase in the neutrophil phagosome: Implications for microbial killing. J. Biol. Chem. 2006, 281, 39860–39869. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Thompson, L.; Lyu, Z.; Cameron, T.A.; De Lay, N.R.; Krachler, A.M.; Ling, J. Optimal translational fidelity is critical for Salmonella virulence and host interactions. Nucleic Acids Res. 2019. Epub ahead of print. [Google Scholar] [CrossRef]
- Bjorkman, J.; Samuelsson, P.; Andersson, D.I.; Hughes, D. Novel ribosomal mutations affecting translational accuracy, antibiotic resistance and virulence of Salmonella Typhimurium. Mol. Microbiol. 1999, 31, 53–58. [Google Scholar] [CrossRef]
- Fan, Y.; Wu, J.; Ung, M.H.; De Lay, N.; Cheng, C.; Ling, J. Protein mistranslation protects bacteria against oxidative stress. Nucleic Acids Res. 2015, 43, 1740–1748. [Google Scholar] [CrossRef]
- Evans, C.R.; Fan, Y.; Ling, J. Increased mistranslation protects E. coli from protein misfolding stress due to activation of a RpoS-dependent heat shock response. FEBS Lett. 2019, 593, 3220–3227. [Google Scholar] [CrossRef]
- Ballesteros, M.; Fredriksson, A.; Henriksson, J.; Nystrom, T. Bacterial senescence: Protein oxidation in non-proliferating cells is dictated by the accuracy of the ribosomes. EMBO J. 2001, 20, 5280–5289. [Google Scholar] [CrossRef]
- Ling, J.; Söll, D. Severe oxidative stress induces protein mistranslation through impairment of an aminoacyl-tRNA synthetase editing site. Proc. Natl. Acad. Sci. USA 2010, 107, 4028–4033. [Google Scholar] [CrossRef]
- Wu, J.; Fan, Y.; Ling, J. Mechanism of oxidant-induced mistranslation by threonyl-tRNA synthetase. Nucleic Acids Res. 2014, 42, 6523–6531. [Google Scholar] [CrossRef] [PubMed]
- Bullwinkle, T.; Reynolds, N.M.; Raina, M.; Moghal, A.B.; Matsa, E.; Rajkovic, A.; Kayadibi, H.; Fazlollahi, F.; Ryan, C.; Howitz, N.; et al. Oxidation of cellular amino acid pools leads to cytotoxic mistranslation of the genetic code. eLife 2014, 3, e02501. [Google Scholar] [CrossRef] [PubMed]
- Steiner, R.E.; Kyle, A.M.; Ibba, M. Oxidation of phenylalanyl-tRNA synthetase positively regulates translational quality control. Proc. Natl. Acad. Sci. USA 2019, 116, 10058–10063. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Gorini, L.; Davis, B.D. Misreading of RNA codewords induced by aminoglycoside antibiotics. Mol. Pharmacol. 1965, 1, 93–106. [Google Scholar] [PubMed]
- Hoffman, L.R.; D’Argenio, D.A.; MacCoss, M.J.; Zhang, Z.; Jones, R.A.; Miller, S.I. Aminoglycoside antibiotics induce bacterial biofilm formation. Nature 2005, 436, 1171–1175. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Wierzbowski, J.; Cottarel, G.; Collins, J.J. Mistranslation of membrane proteins and two-component system activation trigger antibiotic-mediated cell death. Cell 2008, 135, 679–690. [Google Scholar] [CrossRef]
- Samhita, L.; Raval, P.K.; Agashe, D. Global mistranslation increases cell survival under stress in Escherichia coli. PLoS Genet. 2020, 16, e1008654. [Google Scholar] [CrossRef]
- Bregeon, D.; Colot, V.; Radman, M.; Taddei, F. Translational misreading: A tRNA modification counteracts a +2 ribosomal frameshift. Genes Dev. 2001, 15, 2295–2306. [Google Scholar] [CrossRef]
- Shippy, D.C.; Eakley, N.M.; Lauhon, C.T.; Bochsler, P.N.; Fadl, A.A. Virulence characteristics of Salmonella following deletion of genes encoding the tRNA modification enzymes GidA and MnmE. Microb. Pathog. 2013, 57, 1–9. [Google Scholar] [CrossRef]
- Schwartz, M.H.; Waldbauer, J.R.; Zhang, L.; Pan, T. Global tRNA misacylation induced by anaerobiosis and antibiotic exposure broadly increases stress resistance in Escherichia coli. Nucleic Acids Res. 2016, 44, 10292–10303. [Google Scholar] [CrossRef]
- Su, H.W.; Zhu, J.H.; Li, H.; Cai, R.J.; Ealand, C.; Wang, X.; Chen, Y.X.; Kayani, M.U.; Zhu, T.F.; Moradigaravand, D.; et al. The essential mycobacterial amidotransferase GatCAB is a modulator of specific translational fidelity. Nat. Microbiol. 2016, 1, 16147. [Google Scholar] [CrossRef] [PubMed]
- Javid, B.; Sorrentino, F.; Toosky, M.; Zheng, W.; Pinkham, J.T.; Jain, N.; Pan, M.; Deighan, P.; Rubin, E.J. Mycobacterial mistranslation is necessary and sufficient for rifampicin phenotypic resistance. Proc. Natl. Acad. Sci. USA 2014, 111, 1132–1137. [Google Scholar] [CrossRef] [PubMed]
- Durand, J.M.; Dagberg, B.; Uhlin, B.E.; Bjork, G.R. Transfer RNA modification, temperature and DNA superhelicity have a common target in the regulatory network of the virulence of Shigella flexneri: The expression of the virF gene. Mol. Microbiol. 2000, 35, 924–935. [Google Scholar] [CrossRef] [PubMed]
- Fleming, B.A.; Blango, M.G.; Rousek, A.A.; Kincannon, W.M.; Tran, A.; Lewis, A.J.; Russell, C.W.; Zhou, Q.; Baird, L.M.; Barber, A.E.; et al. A tRNA modifying enzyme as a tunable regulatory nexus for bacterial stress responses and virulence. Nucleic Acids Res. 2022, 50, 7570–7590. [Google Scholar] [CrossRef]
- Bacher, J.M.; Waas, W.F.; Metzgar, D.; Crécy-Lagard, V.; Schimmel, P. Genetic code ambiguity confers a selective advantage on Acinetobacter baylyi. J. Bacteriol. 2007, 189, 6494–6496. [Google Scholar] [CrossRef]
- Kermgard, E.; Yang, Z.; Michel, A.M.; Simari, R.; Wong, J.; Ibba, M.; Lazazzera, B.A. Quality control by isoleucyl-tRNA synthetase of Bacillus subtilis is required for efficient sporulation. Sci. Rep. 2017, 7, 41763. [Google Scholar] [CrossRef] [PubMed]
- Kelly, P.; Backes, N.; Mohler, K.; Buser, C.; Kavoor, A.; Rinehart, J.; Phillips, G.; Ibba, M. Alanyl-tRNA synthetase quality control prevents global dysregulation of the Escherichia coli proteome. mBio 2019, 10, e02921-19. [Google Scholar] [CrossRef]
- Aggarwal, S.D.; Lloyd, A.J.; Yerneni, S.S.; Narciso, A.R.; Shepherd, J.; Roper, D.I.; Dowson, C.G.; Filipe, S.R.; Hiller, N.L. A molecular link between cell wall biosynthesis, translation fidelity, and stringent response in Streptococcus pneumoniae. Proc. Natl. Acad. Sci. USA 2021, 118, e2018089118. [Google Scholar] [CrossRef]
- Li, L.; Boniecki, M.T.; Jaffe, J.D.; Imai, B.S.; Yau, P.M.; Luthey-Schulten, Z.A.; Martinis, S.A. Naturally occurring aminoacyl-tRNA synthetases editing-domain mutations that cause mistranslation in Mycoplasma parasites. Proc. Natl. Acad. Sci. USA 2011, 108, 9378–9383. [Google Scholar] [CrossRef]
- Melnikov, S.V.; van den Elzen, A.; Stevens, D.L.; Thoreen, C.C.; Soll, D. Loss of protein synthesis quality control in host-restricted organisms. Proc. Natl. Acad. Sci. USA 2018, 115, E11505–E11512. [Google Scholar] [CrossRef]
- Vargas-Rodriguez, O.; Badran, A.H.; Hoffman, K.S.; Chen, M.; Crnkovic, A.; Ding, Y.; Krieger, J.R.; Westhof, E.; Soll, D.; Melnikov, S. Bacterial translation machinery for deliberate mistranslation of the genetic code. Proc. Natl. Acad. Sci. USA 2021, 118, e2110797118. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Musier-Forsyth, K. Trans-editing of Cys-tRNAPro by Haemophilus influenzae YbaK protein. J. Biol. Chem. 2004, 279, 42359–42362. [Google Scholar] [CrossRef]
- Imlay, J.A. Cellular defenses against superoxide and hydrogen peroxide. Annu. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. The molecular mechanisms and physiological consequences of oxidative stress: Lessons from a model bacterium. Nat. Rev. Microbiol. 2013, 11, 443–454. [Google Scholar] [CrossRef]
- Dock-Bregeon, A.; Sankaranarayanan, R.; Romby, P.; Caillet, J.; Springer, M.; Rees, B.; Francklyn, C.S.; Ehresmann, C.; Moras, D. Transfer RNA-mediated editing in threonyl-tRNA synthetase. The class II solution to the double discrimination problem. Cell 2000, 103, 877–884. [Google Scholar] [CrossRef]
- Beebe, K.; Ribas de Pouplana, L.; Schimmel, P. Elucidation of tRNA-dependent editing by a class II tRNA synthetase and significance for cell viability. EMBO J. 2003, 22, 668–675. [Google Scholar] [CrossRef]
- Dock-Bregeon, A.C.; Rees, B.; Torres-Larios, A.; Bey, G.; Caillet, J.; Moras, D. Achieving error-free translation; the mechanism of proofreading of threonyl-tRNA synthetase at atomic resolution. Mol. Cell. 2004, 16, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wu, J.; Lyu, Z.; Ling, J. Impact of alanyl-tRNA synthetase editing deficiency in yeast. Nucleic Acids Res. 2021, 49, 9953–9964. [Google Scholar] [CrossRef] [PubMed]
- Kavoor, A.; Kelly, P.; Ibba, M. Escherichia coli alanyl-tRNA synthetase maintains proofreading activity and translational accuracy under oxidative stress. J. Biol. Chem. 2022, 298, 101601. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.; Roy, H.; Ibba, M. Mechanism of tRNA-dependent editing in translational quality control. Proc. Natl. Acad. Sci. USA 2007, 104, 72–77. [Google Scholar] [CrossRef]
- Roy, H.; Ling, J.; Irnov, M.; Ibba, M. Post-transfer editing in vitro and in vivo by the beta subunit of phenylalanyl-tRNA synthetase. EMBO J. 2004, 23, 4639–4648. [Google Scholar] [CrossRef]
- Ling, J.; Yadavalli, S.S.; Ibba, M. Phenylalanyl-tRNA synthetase editing defects result in efficient mistranslation of phenylalanine codons as tyrosine. RNA 2007, 13, 1881–1886. [Google Scholar] [CrossRef]
- Bullwinkle, T.; Lazazzera, B.; Ibba, M. Quality control and infiltration of translation by amino acids outside of the genetic code. Annu. Rev. Genet. 2014, 48, 149–166. [Google Scholar] [CrossRef]
- Srinivas, P.; Steiner, R.E.; Pavelich, I.J.; Guerrero-Ferreira, R.; Juneja, P.; Ibba, M.; Dunham, C.M. Oxidation alters the architecture of the phenylalanyl-tRNA synthetase editing domain to confer hyperaccuracy. Nucleic Acids Res. 2021, 49, 11800–11809. [Google Scholar] [CrossRef]
- Willi, J.; Kupfer, P.; Evequoz, D.; Fernandez, G.; Katz, A.; Leumann, C.; Polacek, N. Oxidative stress damages rRNA inside the ribosome and differentially affects the catalytic center. Nucleic Acids Res. 2018, 46, 1945–1957. [Google Scholar] [CrossRef] [PubMed]
- Shcherbik, N.; Pestov, D.G. The Impact of Oxidative Stress on Ribosomes: From Injury to Regulation. Cells 2019, 8, 1379. [Google Scholar] [CrossRef] [PubMed]
- Kyuma, T.; Kizaki, H.; Ryuno, H.; Sekimizu, K.; Kaito, C. 16S rRNA methyltransferase KsgA contributes to oxidative stress resistance and virulence in Staphylococcus aureus. Biochimie 2015, 119, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Youngman, E.M.; McDonald, M.E.; Green, R. Peptide release on the ribosome: Mechanism and implications for translational control. Annu. Rev. Microbiol. 2008, 62, 353–373. [Google Scholar] [CrossRef] [PubMed]
- Kuhlenkoetter, S.; Wintermeyer, W.; Rodnina, M.V. Different substrate-dependent transition states in the active site of the ribosome. Nature 2011, 476, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.J.; Trobro, S.; He, S.L.; Aqvist, J.; Green, R. A Role for the 2’ OH of peptidyl-tRNA substrate in peptide release on the ribosome revealed through RF-mediated rescue. Chem. Biol. 2012, 19, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Indrisiunaite, G.; Pavlov, M.Y.; Heurgue-Hamard, V.; Ehrenberg, M. On the pH dependence of class-1 RF-dependent termination of mRNA translation. J. Mol. Biol. 2015, 427, 1848–1860. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.; Villanueva, P.; O’Malley, L.; Murphy, P.; Ling, J. Genome-wide screening reveals metabolic regulation of translational fidelity. BioRxiv 2022. [Google Scholar]
- Zheng, D.; Constantinidou, C.; Hobman, J.L.; Minchin, S.D. Identification of the CRP regulon using in vitro and in vivo transcriptional profiling. Nucleic Acids Res. 2004, 32, 5874–5893. [Google Scholar] [CrossRef]
- Becker, B.; Cooper, M.A. Aminoglycoside antibiotics in the 21st century. ACS Chem. Biol. 2013, 8, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.P.; Clemons, W.M.; Brodersen, D.E.; Morgan-Warren, R.J.; Wimberly, B.T.; Ramakrishnan, V. Functional insights from the structure of the 30S ribosomal subunit and its interactions with antibiotics. Nature 2000, 407, 340–348. [Google Scholar] [CrossRef]
- Demirci, H.; Murphy, F.t.; Murphy, E.; Gregory, S.T.; Dahlberg, A.E.; Jogl, G. A structural basis for streptomycin-induced misreading of the genetic code. Nat. Commun. 2013, 4, 1355. [Google Scholar] [CrossRef]
- Davis, B.D.; Chen, L.L.; Tai, P.C. Misread protein creates membrane channels: An essential step in the bactericidal action of aminoglycosides. Proc. Natl. Acad. Sci. USA 1986, 83, 6164–6168. [Google Scholar] [CrossRef]
- Fredriksson, A.; Ballesteros, M.; Peterson, C.N.; Persson, O.; Silhavy, T.J.; Nystrom, T. Decline in ribosomal fidelity contributes to the accumulation and stabilization of the master stress response regulator sigmaS upon carbon starvation. Genes Dev. 2007, 21, 862–874. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Gilbert, W.; Gorini, L. Streptomycin, suppression, and the code. Proc. Natl. Acad. Sci. USA 1964, 51, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, D.; Kamath, D.; Gregory, S.T.; O’Connor, M. Modulation of decoding fidelity by ribosomal proteins S4 and S5. J. Bacteriol. 2015, 197, 1017–1025. [Google Scholar] [CrossRef]
- Lyu, Z.; Ling, J. Increase in ribosomal fidelity benefits Salmonella upon bile salt exposure. Genes 2022, 13, 184. [Google Scholar] [CrossRef] [PubMed]
- El Yacoubi, B.; Bailly, M.; de Crecy-Lagard, V. Biosynthesis and function of posttranscriptional modifications of transfer RNAs. Annu. Rev. Genet. 2012, 46, 69–95. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T. The expanding world of tRNA modifications and their disease relevance. Nat. Rev. Mol. Cell Biol. 2021, 22, 375–392. [Google Scholar] [CrossRef] [PubMed]
- Petrullo, L.A.; Gallagher, P.J.; Elseviers, D. The role of 2-methylthio-N6-isopentenyladenosine in readthrough and suppression of nonsense codons in Escherichia coli. Mol. Gen. Genet. 1983, 190, 289–294. [Google Scholar] [CrossRef]
- Vacher, J.; Grosjean, H.; Houssier, C.; Buckingham, R.H. The effect of point mutations affecting Escherichia coli tryptophan tRNA on anticodon-anticodon interactions and on UGA suppression. J. Mol. Biol. 1984, 177, 329–342. [Google Scholar] [CrossRef]
- Ericson, J.U.; Bjork, G.R. Pleiotropic effects induced by modification deficiency next to the anticodon of tRNA from Salmonella Typhimurium LT2. J. Bacteriol. 1986, 166, 1013–1021. [Google Scholar] [CrossRef]
- Blum, P.H. Reduced leu operon expression in a miaA mutant of Salmonella typhimurium. J. Bacteriol. 1988, 170, 5125–5133. [Google Scholar] [CrossRef]
- Gamper, H.B.; Masuda, I.; Frenkel-Morgenstern, M.; Hou, Y.M. Maintenance of protein synthesis reading frame by EF-P and m(1)G37-tRNA. Nat. Commun. 2015, 6, 7226. [Google Scholar] [CrossRef]
- Bystrom, A.S.; Bjork, G.R. The structural gene (trmD) for the tRNA(m1G)methyltransferase is part of a four polypeptide operon in Escherichia coli K-12. Mol. Gen. Genet. 1982, 188, 447–454. [Google Scholar] [CrossRef]
- Bjork, G.R.; Wikstrom, P.M.; Bystrom, A.S. Prevention of translational frameshifting by the modified nucleoside 1-methylguanosine. Science 1989, 244, 986–989. [Google Scholar] [CrossRef]
- Gamper, H.; Li, H.; Masuda, I.; Miklos Robkis, D.; Christian, T.; Conn, A.B.; Blaha, G.; Petersson, E.J.; Gonzalez, R.L., Jr.; Hou, Y.M. Insights into genome recoding from the mechanism of a classic +1-frameshifting tRNA. Nat. Commun. 2021, 12, 328. [Google Scholar] [CrossRef] [PubMed]
- Masuda, I.; Hwang, J.Y.; Christian, T.; Maharjan, S.; Mohammad, F.; Gamper, H.; Buskirk, A.R.; Hou, Y.M. Loss of N(1)-methylation of G37 in tRNA induces ribosome stalling and reprograms gene expression. eLife 2021, 10, e70619. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.M.; Matsubara, R.; Takase, R.; Masuda, I.; Sulkowska, J.I. TrmD: A Methyl Transferase for tRNA Methylation With m(1)G37. Enzymes 2017, 41, 89–115. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Ehrlich, S.D.; Albertini, A.; Amati, G.; Andersen, K.K.; Arnaud, M.; Asai, K.; Ashikaga, S.; Aymerich, S.; Bessieres, P.; et al. Essential Bacillus subtilis genes. Proc. Natl. Acad. Sci. USA 2003, 100, 4678–4683. [Google Scholar] [CrossRef] [PubMed]
- O’Dwyer, K.; Watts, J.M.; Biswas, S.; Ambrad, J.; Barber, M.; Brule, H.; Petit, C.; Holmes, D.J.; Zalacain, M.; Holmes, W.M. Characterization of Streptococcus pneumoniae TrmD, a tRNA methyltransferase essential for growth. J. Bacteriol. 2004, 186, 2346–2354. [Google Scholar] [CrossRef] [PubMed]
- Bjork, G.R.; Jacobsson, K.; Nilsson, K.; Johansson, M.J.; Bystrom, A.S.; Persson, O.P. A primordial tRNA modification required for the evolution of life? EMBO J. 2001, 20, 231–239. [Google Scholar] [CrossRef]
- Masuda, I.; Matsubara, R.; Christian, T.; Rojas, E.R.; Yadavalli, S.S.; Zhang, L.; Goulian, M.; Foster, L.J.; Huang, K.C.; Hou, Y.M. tRNA methylation is a global determinant of bacterial multi-drug resistance. Cell Syst. 2019, 8, 302–314.e308. [Google Scholar] [CrossRef]
- Groisman, E.A.; Hollands, K.; Kriner, M.A.; Lee, E.J.; Park, S.Y.; Pontes, M.H. Bacterial Mg2+ homeostasis, transport, and virulence. Annu. Rev. Genet. 2013, 47, 625–646. [Google Scholar] [CrossRef]
- Gall, A.R.; Datsenko, K.A.; Figueroa-Bossi, N.; Bossi, L.; Masuda, I.; Hou, Y.M.; Csonka, L.N. Mg2+ regulates transcription of mgtA in Salmonella Typhimurium via translation of proline codons during synthesis of the MgtL peptide. Proc. Natl. Acad. Sci. USA 2016, 113, 15096–15101. [Google Scholar] [CrossRef]
- Yamada, Y.; Murao, K.; Ishikura, H. 5-(carboxymethylaminomethyl)-2-thiouridine, a new modified nucleoside found at the first letter position of the anticodon. Nucleic Acids Res. 1981, 9, 1933–1939. [Google Scholar] [CrossRef]
- Meyer, S.; Wittinghofer, A.; Versees, W. G-domain dimerization orchestrates the tRNA wobble modification reaction in the MnmE/GidA complex. J. Mol. Biol. 2009, 392, 910–922. [Google Scholar] [CrossRef] [PubMed]
- Yim, L.; Moukadiri, I.; Bjork, G.R.; Armengod, M.E. Further insights into the tRNA modification process controlled by proteins MnmE and GidA of Escherichia coli. Nucleic Acids Res. 2006, 34, 5892–5905. [Google Scholar] [CrossRef] [PubMed]
- Urbonavicius, J.; Qian, Q.; Durand, J.M.; Hagervall, T.G.; Bjork, G.R. Improvement of reading frame maintenance is a common function for several tRNA modifications. EMBO J. 2001, 20, 4863–4873. [Google Scholar] [CrossRef] [PubMed]
- Urbonavicius, J.; Stahl, G.; Durand, J.M.; Ben Salem, S.N.; Qian, Q.; Farabaugh, P.J.; Bjork, G.R. Transfer RNA modifications that alter +1 frameshifting in general fail to affect -1 frameshifting. RNA 2003, 9, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Jager, G.; Nilsson, K.; Bjork, G.R. The phenotype of many independently isolated +1 frameshift suppressor mutants supports a pivotal role of the P-site in reading frame maintenance. PLoS ONE 2013, 8, e60246. [Google Scholar] [CrossRef] [PubMed]
- Shippy, D.C.; Heintz, J.A.; Albrecht, R.M.; Eakley, N.M.; Chopra, A.K.; Fadl, A.A. Deletion of glucose-inhibited division (gidA) gene alters the morphological and replication characteristics of Salmonella enterica Serovar typhimurium. Arch. Microbiol. 2012, 194, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, S.; Tamaru, A.; Nakajima, C.; Nishimura, K.; Tanaka, Y.; Tokuyama, S.; Suzuki, Y.; Ochi, K. Loss of a conserved 7-methylguanosine modification in 16S rRNA confers low-level streptomycin resistance in bacteria. Mol. Microbiol. 2007, 63, 1096–1106. [Google Scholar] [CrossRef]
- Mikheil, D.M.; Shippy, D.C.; Eakley, N.M.; Okwumabua, O.E.; Fadl, A.A. Deletion of gene encoding methyltransferase (gidB) confers high-level antimicrobial resistance in Salmonella. J. Antibiot. 2012, 65, 185–192. [Google Scholar] [CrossRef]
- Sheppard, K.; Akochy, P.M.; Salazar, J.C.; Söll, D. The Helicobacter pylori amidotransferase GatCAB is equally efficient in glutamine-dependent transamidation of Asp-tRNAAsn and Glu-tRNAGln. J. Biol. Chem. 2007, 282, 11866–11873. [Google Scholar] [CrossRef]
- Nakamura, A.; Yao, M.; Chimnaronk, S.; Sakai, N.; Tanaka, I. Ammonia channel couples glutaminase with transamidase reactions in GatCAB. Science 2006, 312, 1954–1958. [Google Scholar] [CrossRef]
- Li, Y.Y.; Cai, R.J.; Yang, J.Y.; Hendrickson, T.L.; Xiang, Y.; Javid, B. Clinically relevant mutations of mycobacterial GatCAB inform regulation of translational fidelity. mBio 2021, 12, e0110021. [Google Scholar] [CrossRef] [PubMed]
- Rottem, S. Interaction of mycoplasmas with host cells. Physiol. Rev. 2003, 83, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Han, N.C.; Kavoor, A.; Ibba, M. Characterizing the amino acid activation center of the naturally editing-deficient aminoacyl-tRNA synthetase PheRS in Mycoplasma mobile. FEBS Lett. 2022, 596, 947–957. [Google Scholar] [CrossRef]
- Shepherd, J.; Ibba, M. Lipid II-independent trans editing of mischarged tRNAs by the penicillin resistance factor MurM. J. Biol. Chem. 2013, 288, 25915–25923. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Kim, I.S. Identification of macrophage genes responsive to extracellular acidification. Inflamm. Res 2013, 62, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Aguirre Rivera, J.; Larsson, J.; Volkov, I.L.; Seefeldt, A.C.; Sanyal, S.; Johansson, M. Real-time measurements of aminoglycoside effects on protein synthesis in live cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2013315118. [Google Scholar] [CrossRef] [PubMed]
- Eggertsson, G.; Söll, D. Transfer ribonucleic acid-mediated suppression of termination codons in Escherichia coli. Microbiol. Rev. 1988, 52, 354–374. [Google Scholar] [CrossRef]
- Santos, M.A.; Tuite, M.F. The CUG codon is decoded in vivo as serine and not leucine in Candida albicans. Nucleic Acids Res. 1995, 23, 1481–1486. [Google Scholar] [CrossRef]
- Lant, J.T.; Berg, M.D.; Heinemann, I.U.; Brandl, C.J.; O’Donoghue, P. Pathways to disease from natural variations in human cytoplasmic tRNAs. J. Biol. Chem. 2019, 294, 5294–5308. [Google Scholar] [CrossRef]


{kind=link}
| Error Types | Bacteria | Sources of Error | Phenotypes | Ref. |
|---|---|---|---|---|
| Global | S. typhimurium, E. coli | Mutations in rpsD | Decreased cell invasion and animal colonization; increased resistance against oxidative stress and heat; decreased motility | [26,27,28,29] |
| High-fidelity | S. typhimurium, E. coli | Mutations in rpsL | Decreased cell invasion and animal colonization; decreased resistance against oxidative stress; decreased motility | [26,27,28] |
| Readthrough | E. coli | Carbon starvation | Increased protein oxidation during aging | [30] |
| Thr → Ser | E. coli | Oxidative stress damages the editing site of ThrRS | Mild growth defect with excess Ser | [13,31,32] |
| Phe → m-Tyr | E. coli, S. typhimurium, | Oxidation of Tyr to m-Tyr | PheRS editing defect decreases growth under oxidative stress | [33,34] |
| Global | E. coli, P. aeruginosa | Aminoglycosides | Bactericidal; increased biofilm formation at sublethal doses | [35,36,37] |
| Initiation errors | E. coli | Deleting initiator tRNAs | Increased tolerance to fluoroquinolones and heat stress | [38] |
| Frameshift | S. typhimurium | Deleting gidA or mnmE | Mutations in gidA and mnmE decreases Salmonella invasion and host colonization | [39,40] |
| Readthrough | E. coli, S. typhimurium | Acid stress, excess sugar | May promote tolerance to acid stress | [15] |
| Readthrough | E. coli | Chloramphenicol, etc. | Unclear | [16] |
| Multiple AA → Met | E. coli | Anaerobic growth and antibiotic stress | Decreased MetRS succinylation increases Met misacylation | [41] |
| Gln → Glu, Asn → Asp | M. smegmatis M. tuberculosis | Mutations in tRNAs or gatCAB | Increased phenotypic resistance to rifampicin | [42,43] |
| Readthrough | S. flexneri | Deleting miaA | Decreased expression of virulence genes | [44] |
| Frameshift | ExPEC | Deleting or overexpressing miaA | Deleting miaA attenuates virulence | [45] |
| Ile → Val | A. baylyi | Editing-defective IleRS | Improved growth with excess Val | [46] |
| Ile → Val | B. subtilis | Editing-defective IleRS | Sporulation defect | [47] |
| Ala → Ser | E. coli | C666A mutation in AlaRS | Decreased motility | [48] |
| Ala → Ser | S. pneumonia | Deleting murMN | Decreased macrophage phagocytosis | [49] |
| Leu → Val, Phe → Tyr etc. | M. mobile and other host-restricted bacteria | Natural editing-defective aaRSs | May be adaptive to parasitic cycle | [50,51] |
| Ala → Pro | Streptomyces spp. | ProRS/tRNAProA pair | Unclear | [52] |
| Pro → Ala | C. sticklandii, P. aeruginosa etc. | Deleting proX | Unclear | [9,10] |
| Pro → Cys | H. influenza, C. crescentus etc. | Deleting ybaK | Unclear | [10,53] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, Z.; Wilson, C.; Ling, J. Translational Fidelity during Bacterial Stresses and Host Interactions. Pathogens 2023, 12, 383. https://doi.org/10.3390/pathogens12030383
Lyu Z, Wilson C, Ling J. Translational Fidelity during Bacterial Stresses and Host Interactions. Pathogens. 2023; 12(3):383. https://doi.org/10.3390/pathogens12030383
Chicago/Turabian StyleLyu, Zhihui, Cierra Wilson, and Jiqiang Ling. 2023. "Translational Fidelity during Bacterial Stresses and Host Interactions" Pathogens 12, no. 3: 383. https://doi.org/10.3390/pathogens12030383
APA StyleLyu, Z., Wilson, C., & Ling, J. (2023). Translational Fidelity during Bacterial Stresses and Host Interactions. Pathogens, 12(3), 383. https://doi.org/10.3390/pathogens12030383