

The Role of Immunity in the Pathogenesis of SARS-CoV-2 Infection and in the Protection Generated by COVID-19 Vaccines in Different Age Groups

 ,
,
Abstract
1. Introduction
2. Review of the Literature and Discussion
2.1. Development and Maturation of the Normal Immune System
2.1.1. The First Stage during Infancy
Innate Immunity
Adaptive Immunity
2.1.2. The Second Stage: From Childhood to Adolescence–Adulthood
3. Immune Response to the Different Types of Vaccines in Children
3.1. Immune Response to COVID-19 Vaccines
3.2. Safety of Anti-COVID-19 Vaccines from Infancy to Adolescence
3.2.1. Live-Attenuated Vaccines
3.2.2. Inactivated Vaccines
3.2.3. DNA Vaccines
3.2.4. RNA Vaccines
3.2.5. Subunit Vaccines
3.2.6. Vector Vaccines
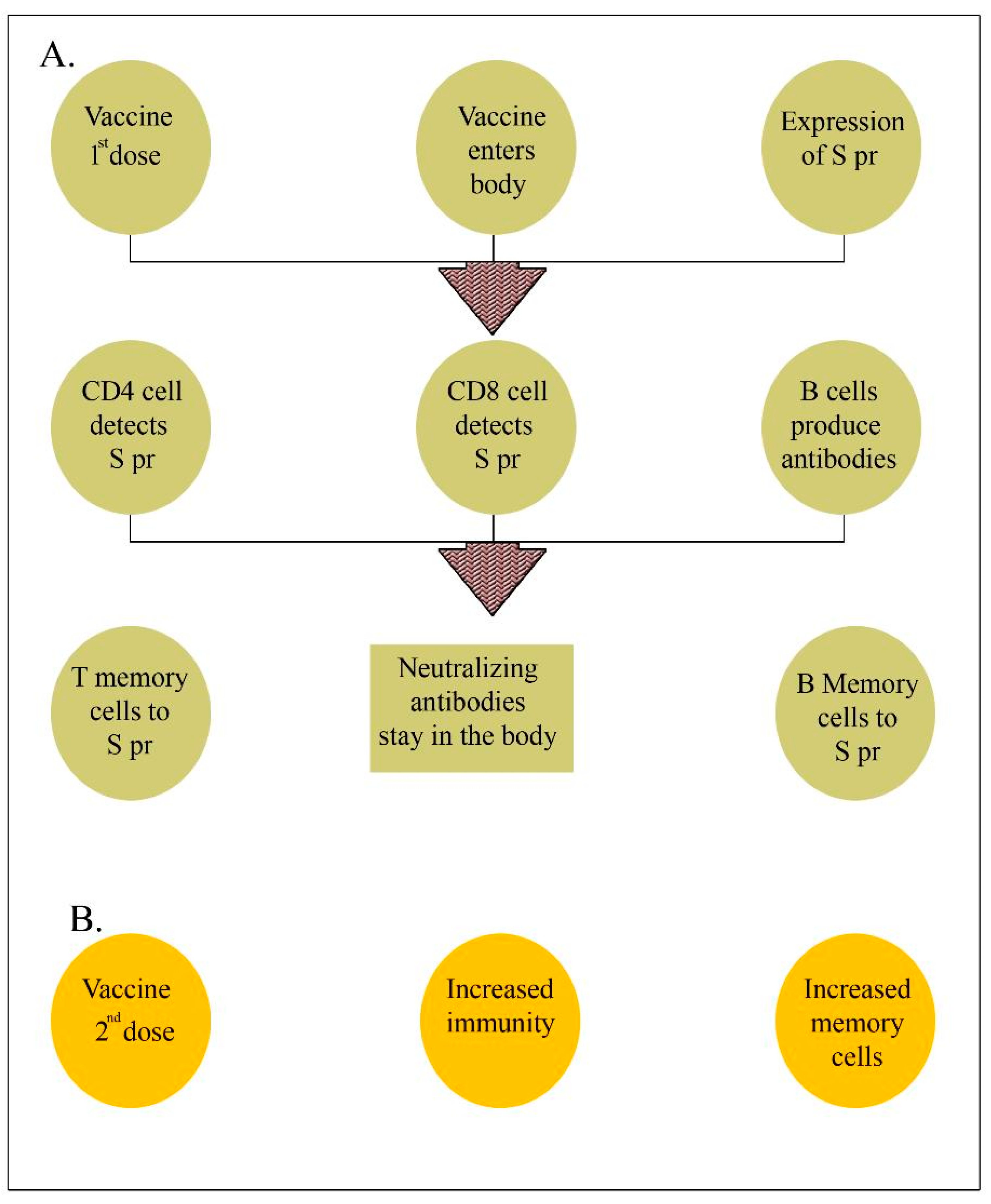
3.3. Different Immunological Mechanisms after COVID-19 Vaccination

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccines | Manufacturers | Immune Response |
|---|---|---|
| mRNA - BNT162b2 - mRNA-1273 | - Pfizer/BioNTech + Fosun Pharma - Moderna + National Institute of Allergic and Infectious diseases | Expression of the viral S protein encoded in the mRNA and antigen presentation; induction of IFN I release, stimulating Th1 response, antibodies, and memory T and B cell generation |
| Adenoviral Vectored - AZD122 (ChAdOx1-S) - Ad26.COV2 S - Sputnik V (rAd26-S + rAd5-S) | - AstraZeneca + University of Oxford - Janssen Pharmaceutical by Johnson & Johnson - Gamaleya Research Institute of Epidemiology and Microbiology | Emulation of viral infection; induces expression of IFN, antibodies, and memory T and B cells, along with T CD8 activity |
| Inactivated - CoronaVac - Sinopharm - Covaxin - Valneva | Chinese Sinovac Biotech. BIBP B Beijing BIBP and WIBP Indian Baharat Biotech French Biotechnology Com | Similar to adenoviral vectors; induction of IFN, memory T and B cells, antibody production, and T CD8 activity |
| Protein Subunit - PREVENT-19 (NVX CoV-2373) | Novavax | Enhanced humoral responses and B and T memory cells |
| Live-Attenuated - COVI-VAC | - Codagenix and Serum Institute of India | Induces antibody-based humoral and T lymphocyte-based cellular immune responses |
3.4. Vaccination of Children Aged 5–15 Years
3.5. Vaccination for Children Aged Less Than Five Years
3.6. Virology of SARS-CoV-2 and the Generated Immunity against Viral Components
3.6.1. Role of ACE Receptor Expression during COVID-19 Infection and Vaccination
3.6.2. Classes of Antibodies Produced in Response to COVID-19 Vaccines
3.6.3. Viral Mutations and Childhood Immunization
3.7. Safety of COVID-19 Vaccines for Infants and Children
3.7.1. Parental Hesitancy towards Vaccination of Their Children
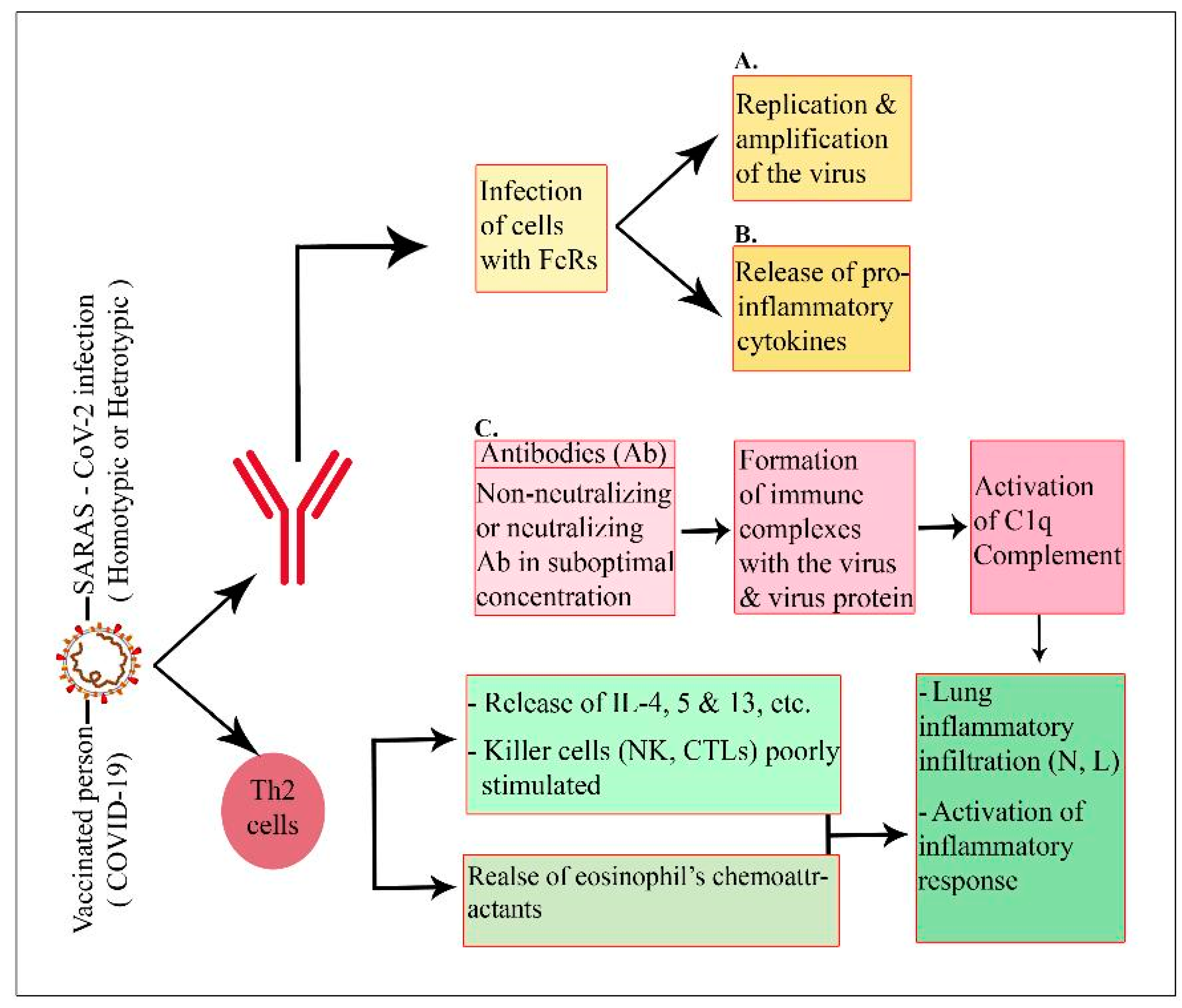
3.7.2. Mechanisms of the Adverse Effects of Vaccination in Adults and Children
4. Immune Response to SARS-CoV-2 Infection
4.1. COVID-19-Generated Immunity in Neonates to Adolescents
| Immunocompetent Cells | Immunological Elements | Functions |
|---|---|---|
| B cells | SARS-CoV-2 neutralizing antibodies | - Viral neutralization |
| - Antibody-dependent cellular cytotoxicity (ADCC) | ||
| - Antibody-dependent cellular phagocytosis (ADCP) | ||
| - Complement activation (ADCA) [71,150,151,152,153,154,155,156] | ||
| TFH cells | CD4-positive cells | - Interact with B cells to enhance survival and provide cellular help for antibody production [11,36,71,148,161] * |
| CD4 Th1 T cells | IL-2, IFN-γ, and TNF-α | - CD8 T cell help, inflammation, and killing of virally infected cells |
| CD4 Th2 T cells | IL-4, IL-5, and IL-13 | - B cell help and induction of antibody class-switching |
| CD4 Th17 cells | IL-17A, IL-17F, IL-21, and IL-22 | - Inflammation via neutrophil recruitment and activation and innate cell activation |
| CD4 Regulatory T cell (Treg) | IL-10 and TGF-β | - Suppression of inflammation and other T cells via cytokines or contact-dependent mechanisms |
| CD8 cytotoxic T cells | Granzymes, perforin, IFN-γ, TNF-α, and FASL expression | - Killing of virally infected cells [29,44,45,46,47,48,63,71,74] |

4.2. Clinical Information Regarding COVID-19 in Children
4.3. Severe COVID-19 Immunopathology in Kids and Adults
4.3.1. Innate Immunity’s Function
4.3.2. The Role of Adaptive Immunity
The Cellular Components
The Humoral Response
4.3.3. Anamnestic Responses to SARS-CoV-2
4.3.4. Cytokine Storm
| (A) [67,173,174,176,195] | ||||||
|---|---|---|---|---|---|---|
| Disease Severity | B Cells | Plasma Cells | IgG/IgA | B Cells | Plasma Cells | IgG/IgA |
| (Adults) | (Children) | |||||
| Severe/PIMS-Ts | ↓ | ↑ | ↑ ss | ↓ | ↓ | ↓ |
| Mild | uc | ↑ | ↑ tr | uc | ↑ | ↑ |
| Asymptomatic | uc | ↑ | ↑ tr | uc | ↑ | ↑ |
| (B) [174,176,182,195] | ||||||
| Disease Severity | CD3+ | CD8+ | CD4+ | CD8+ | CD8+ | CD4+ |
| (Adults) | (Children) | |||||
| Severe/PIMS-Ts | ↓ | ↓ | ↓ | ↓ | ↓ | ↓ |
| Mild | ↑ | ↑ | ↑ | ↑ | ↑ | ↑ |
| Asymptomatic | uc | uc | uc | uc | un | un |
| (C) [17,67,156,173,195] | ||||||
| Disease Severity | Monocytes | Neutrophils | NK | Monocytes | Neutrophils | NK |
| (Adults) | (Children) | |||||
| Severe/PIMS-Ts | ↑ | ↑ | ↓ | ↓ | ↓ | ↓ |
| Mild | ↑ | ↑ | ↓ | nr | nr | ↑ |
| Asymptomatic | nr | nr | ↑ | nr | nr | ↑ |
4.3.5. PIMS-TS in Severe Pediatric COVID19
4.4. Energy Allocation Perspective on COVID-19 in Children
5. Concluding Immunological Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anderson, E.J.; Campbell, J.D.; Creech, C.B.; Frenck, R.; Kamidani, S.; Munoz, F.M.; Nachman, S.; Spearmanet, P. Warp spread of coronavirus disease 2019 (COVID-19) vaccines: Why are children stuck in neutral? Clin. Infect. Dis. 2020, 73, 336–340. [Google Scholar] [CrossRef]
- McGown, R.J.; Cennimo, D.J. Coronavirus Disease 2019 (COVID-19) in Children. Available online: https://emedicine.medscape.com/article/2500132-overview (accessed on 10 February 2022).
- Mayo Clinic Staff. COVID-19 in Babies and Children. Available online: https://www.mayoclinic.org/diseases-conditions/coronavirus/in-depth/coronavirus-in-babies-and-children/art-20484405 (accessed on 7 July 2022).
- Peppers, B. Why Vaccine Doses Differ for Babies, Kids, Teens and Adults—An Immunologist Explains How Your Immune System Changes as You Mature. The Conversation. Available online: https://theconversation.com/why-vaccine-doses-differ-for-babies-kids-teens-and-adults-an-immunologist-explains-how-your-immune-system-changes-as-you-mature-168708 (accessed on 21 November 2021).
- Elahi, S.H. Neonatal and children’s immune system and COVID-19: Biased immune tolerance versus resistance strategy. J. Immunol. 2020, 205, 1990–1997. [Google Scholar] [CrossRef]
- Cao, Q.; Chen, Y.C.; Chen, C.L.; Chiu, C.H. SARS-CoV-2 infection in children: Transmission dynamics and clinical characteristics. J. Formos. Med. Assoc. 2020, 119, 670–673. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. China Medical Treatment Expert Group for Covid-19. Clinical characteristics of coronavirus disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Behnke, J.M.; Barnard, C.J.; Wakelin, D. Understanding chronic nematode infections: Evolutionary considerations, current hypotheses and the way forward. Int. J. Parasitol. 1992, 22, 861–907. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R.; Schneider, D.S.; Soares, M.P. Disease tolerance as a defense strategy. Science 2012, 335, 936–941. [Google Scholar] [CrossRef]
- Yu, J.C.; Khodadadi, H.; Malik, A.; Davidson, B.; Salles, E.D.S.L.; Bahatia, J.; Hale, V.L.; Baban, B. Innate immunity of neonates and infants. Front. Immunol. 2018, 9, 1759–1778. [Google Scholar] [CrossRef]
- Fidel, P.L., Jr.; Noverr, M.C. Could an unrelated live attenuated vaccine serves as a preventive measure to dampen septic inflammation associated with COVID-19 infection? mBio 2020, 11, e00907-20. [Google Scholar] [CrossRef] [PubMed]
- Elahi, S.; Ertelt, J.M.; Kinder, M.; Jaing, T.T.; Zhang, X.; Xin, X.; Chaturvedi, V.; Stronh, B.S.; Qualls, J.E.; Steinbrecher, K.A.; et al. Immunosuppressive CD71+ erythroid cells compromise neonatal host defence against infection. Nature 2013, 504, 158–162. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Guo, J.; Huang, Z.; Lin, L.; Lv, J. Coronavirus disease 2019 (COVID-19) and cardiovascular disease: A viewpoint on the potential influence of angiotensin—Converting enzyme inhibitors/angiotensin receptor blockers on onset and severity of acute respiratory syndrome coronavirus 2 infection. J. Am. Heart Assoc. 2020, 9, e016219. [Google Scholar] [CrossRef] [PubMed]
- Bhat, B.V.; Ravikumar, S. Challenges in neonatal COVID-19 infection. Indian J. Pediatr. 2020, 87, 577–578. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Baker, A. Recombinant human ACE2: Acing out angiotensin II in ARDS therapy. Crit. Care 2017, 21, 305. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; McGoogan, J.M. Characteristics of and important lessons from the coronavirus disease 2019 (COVID-19) outbreak in China. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef] [PubMed]
- Livingston, E.; Bucher, K. Coronavirus Disease 2019 (COVID-19) in Italy. JAMA 2020, 323, 1335. [Google Scholar] [CrossRef]
- CDC COVID-19 Response Team. Coronavirus Disease 2019 in Children—United States, 12 February–2 April 2020. Morb. Mortal Wkly. Rep. 2020, 69, 422–426. [Google Scholar] [CrossRef]
- Walter, E.B.; Talaat, K.R.; Sabharwal, C.H.; Gurtman, A.; Lockhart, S.; Paulsen, G.C.; Barnett, E.D.; Muñoz, F.M.; Maldonado, Y.; Pahud, B.A.; et al. Evaluation of the BNT162b2 Covid-19 Vaccine in Children 5 to 11 Years of Age. N. Engl. J. Med. 2022, 386, 35–46. [Google Scholar] [CrossRef]
- Nussbaum, C.; Gloning, A.; Pruenster, M.; Frommhold, D.; Bierschenk, S.; Genzel-Boroviczeny, O.; von Andrian, U.H.; Quackenbush, E.; Sperandio, M. Neutrophil and endothelial adhesive function during human fetal ontogeny. J. Leukoc. Biol. 2013, 93, 175–184. [Google Scholar] [CrossRef]
- Filias, A.; Theodorou, G.L.; Mouzopoulou, S.; Varvarigou, A.A.; Mantagos, S.; Karakantza, M. Phagocytic ability of neutrophils and monocytes in neonates. BMC Pediatr. 2011, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.M.; Slifka, M.K. Longevity of T-cell memory following acute viral infection. Adv. Exp. Med. Biol. 2010, 684, 96–107. [Google Scholar] [CrossRef]
- Förster-Waldl, E.; Sadeghi, K.; Tamandl, D.; Gerhold, B.; Hallwirth, U.; Rohrmeister, K.; Hayde, M.; Prusa, A.R.; Herkner, K.; Boltz-Nitulescu, G.; et al. Monocyte toll-like receptor 4 expression and LPS-induced cytokine production increase during gestational aging. Pediatr. Res. 2005, 58, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.R.; Qing, G.; Byers, D.M.; Stadnyk, A.W.; Al-Hertani, W.; Bortolussi, R. Role of MyD88 in diminished tumor necrosis factor alpha production by newborn mononuclear cells in response to lipopolysaccharide. Infect. Immun. 2004, 72, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, K.; Berger, A.; Langgartner, M.; Prusa, A.R.; Hayde, M.; Herkner, K.; Pollak, A.; Spittler, A.; Forster-Waldl, E. Immaturity of infection control in preterm and term newborns is associated with impaired toll-like receptor signaling. J. Infect. Dis. 2007, 195, 296–302. [Google Scholar] [CrossRef]
- Al-Hertani, W.; Yan, S.R.; Byers, D.M.; Bortolussi, R. Human newborn polymorphonuclear neutrophils exhibit decreased levels of MyD88 and attenuated p38 phosphorylation in response to lipopolysaccharide. Clin. Investig. Med. 2007, 30, E44–E53. [Google Scholar] [CrossRef]
- Blahnik, M.J.; Ramanathan, R.; Riley, C.R.; Minoo, P. Lipopolysaccharide-induced tumor necrosis factor-alpha and IL-10 production by lung macrophages from preterm and term neonates. Pediatr. Res. 2001, 50, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Willems, F.; Vollstedt, S.; Suter, M. Phenotype and function of neonatal DC. Eur. J. Immunol. 2009, 39, 26–35. [Google Scholar] [CrossRef]
- Schüller, S.S.; Sadeghi, K.; Wisgrill, L.; Dangl, A.; Diesner, S.C.; Prusa, A.R.; Klebermasz-Schrehof, K.; Greber-Platzer, S.; Neumüller, J.; Helmer, H.; et al. Preterm neonates display altered plasmacytoid dendritic cell function and morphology. J. Leukoc. Biol. 2013, 93, 781–788. [Google Scholar] [CrossRef]
- De Wit, D.; Tonon, S.; Olislagers, V.; Goriely, S.; Boutriaux, M.; Goldman, M.; Willems, F. Impaired responses to toll-like receptor 4 and toll-like receptor 3 ligands in human cord blood. J. Autoimmun. 2003, 21, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Ivarsson, M.A.; Loh, L.; Marquardt, N.; Kekalainen, E.; Berglin, L.; Bjorkstrom, N.K.; Westgren, M.; Nixon, D.F.; Michaelsson, J. Differentiation and functional regulation of human fetal NK cells. J. Clin. Investig. 2013, 123, 3889–3901. [Google Scholar] [CrossRef]
- McGreal, E.P.; Hearne, K.; Spiller, O.B. Off to a slow start: Under-development of the complement system in term newborns is more substantial following premature birth. Immunobiology 2012, 217, 176–186. [Google Scholar] [CrossRef]
- Mold, J.E.; Venkatasubrahmanyam, S.; Burt, T.D.; Michaelsson, J.; Rivera, J.M.; Galkina, S.A.; Weinberg, K.; Stoddart, C.A.; McCune, J.M. Fetal and adult hematopoietic stem cells give rise to distinct T cell lineages in humans. Science 2010, 330, 1695–1699. [Google Scholar] [CrossRef] [PubMed]
- Takahata, Y.; Nomura, A.; Takada, H.; Ohga, S.; Furuno, K.; Hikino, S.; Nakayama, H.; Sakaguchi, S.; Hara, T. CD25+CD4+ T cells in human cord blood: An immunoregulatory subset with naive phenotype and specific expression of forkhead box p3 (Foxp3) gene. Exp. Hematol. 2004, 32, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Al Attas, R.A.; Liacini, A. Tolerance to noninherited maternal antigens allows successful second kidney transplantation: A case report. Transplant. Proc. 2019, 51, 526–529. [Google Scholar] [CrossRef]
- Mackroth, M.S.; Malhotra, I.; Mungai, P.; Koech, D.; Muchiri, E.; King, C.L. Human cord blood CD4+CD25hi regulatory T cells suppress prenatally acquired T cell responses to Plasmodium falciparum antigens. J. Immunol. 2011, 186, 2780–2791. [Google Scholar] [CrossRef]
- Holt, P.G. The role of genetic and environmental factors in the development of T-cell mediated allergic disease in early life. Paediatr. Respir. Rev. 2004, 5, S27–S30. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, D.; Fleming, P.; Virasami, A.; Michel, M.L.; Sebire, N.J.; Costeloe, K.; Carr, R.; Klein, N.; Hayday, A. Interleukin-8 (CXCL8) production is a signatory T cell effector function of human newborn infants. Nat. Med. 2014, 20, 1206–1210. [Google Scholar] [CrossRef]
- Gibbons, D.L.; Haque, S.F.; Silberzahn, T.; Hamilton, K.; Langford, C.; Ellis, P.; Carr, R.; Hayday, A.C. Neonates harbour highly active gamma-delta T cells with selective impairments in preterm infants. Eur. J. Immunol. 2009, 39, 1794–1806. [Google Scholar] [CrossRef]
- Sanz, E.; Munoz, A.N.; Monserrat, J.; Van-Den-Rym, A.; Escoll, P.; Ranz, I.; Alvarez-Mon, M.; de-la-Hera, A. Ordering human CD34+CD10−CD19+ pre/pro-B-cell and CD19- common lymphoid progenitor stages in two pro-B-cell development pathways. Proc. Natl Acad. Sci. USA 2010, 107, 5925–5930. [Google Scholar] [CrossRef]
- Griffin, D.O.; Rothstein, T.L. A small CD11b+ human B1 cell subpopulation stimulates T cells and is expanded in lupus. J. Exp. Med. 2011, 208, 2591–2598. [Google Scholar] [CrossRef]
- Pihlgren, M.; Friedli, M.; Tougne, C.; Rochat, A.F.; Lambert, P.H.; Siegrist, C.A. Reduced ability of neonatal and early-life bone marrow stromal cells to support plasmablast survival. J. Immunol. 2006, 176, 165–172. [Google Scholar] [CrossRef]
- Mohey Eldin, M.; El Shikh, M.M.; Pitzalis, C. Follicular dendritic cells in health and disease. Front. Immunol. 2012, 3, 292. [Google Scholar] [CrossRef]
- De Kleer, I.; Willems, F.; Lambrecht, B.; Goriely, S. Ontogeny of myeloid cells. Front. Immunol. 2014, 5, 423. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Torchinsky, M.B.; Gobert, M.; Xiong, H.; Xu, M.; Linehan, J.L.; Alonzo, F.; Ng, C.; Chen, A.; Lin, X.; et al. Focused specificity of intestinal TH17 cells towards commensal bacterial antigens. Nature 2014, 510, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Ivaylo, I.I.; Atarashi, K.; Manel, N.; Brodie, E.L.; Shima, T.; Karaoz, U.; Wei, D.; Goldfar, K.C.; Santee, C.A.; Lynch, S.V.; et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 2009, 139, 485–498. [Google Scholar] [CrossRef]
- Tosato, F.; Bucciol, G.; Pantano, G.; Putti, M.C.; Sanzari, M.C.; Basso, G.; Plebani, M. Lymphocytes subsets reference values in childhood. Cytometry 2015, 87A, 81–85. [Google Scholar] [CrossRef]
- Schramm, C.A.; Douek, D.C. Beyond hot spots: Biases in antibody somatic hypermutation and implications for vaccine design. Front. Immunol. 2018, 9, 1876. [Google Scholar] [CrossRef]
- Jung, J.H.; Rha, M.-S.; Sa, M.; Choi, H.K.; Jeon, J.H.; Seok, H.; Park, D.W.; Park, S.-H.; Jeong, H.W.; Choi, W.S.; et al. SARS-CoV-2-specific T cell memory is sustained in COVID-19 convalescent patients for 10 months with successful development of stem cell-like memory T cells. Nat. Commun. 2021, 12, 4043. [Google Scholar] [CrossRef]
- Zinkernagel, R.M. On immunological memory. Phil. Trans. R. Soc. Lond. 2000, 355, 369–371. [Google Scholar] [CrossRef]
- Hayward, A.C.; Fragaszy, E.B.; Bermingham, A.; Wang, L.; Copas, A.; Edmunds, W.J.; Ferguson, N.; Goonetilleke, N.; Harvey, G.; Kovar, J.; et al. Comparative community burden and severity of seasonal and pandemic influenza: Results of the Flu Watch cohort study. Lancet Respir. Med. 2014, 2, 445–454. [Google Scholar] [CrossRef]
- Campion, S.L.; Brodie, T.M.; Fischer, W.; Korber, B.T.; Rossetti, A.; Goonetilleke, N.; McMichael, A.J.; Sallusto, F. Proteome-wide analysis of HIV-specific naive and memory CD4+ T cells in unexposed blood donors. J. Exp. Med. 2014, 211, 1273–1280. [Google Scholar] [CrossRef]
- Round, J.L.; O’Connell, R.M.; Mazmanian, S.K. Coordination of tolerogenic immune responses by the commensal microbiota. J. Autoimmun. 2010, 34, J220–J225. [Google Scholar] [CrossRef]
- Su, L.F.; Kidd, B.A.; Han, A.; Kotzin, J.J.; Davis, M.M. Virus-specific CD4+ memory-phenotype T cells are abundant in unexposed adults. Immunity 2013, 38, 373–383. [Google Scholar] [CrossRef]
- Rowntree, L.C.; Nguyen, T.H.O.; Halim, H.; Purcell, A.W.; Rossjohn, J.; Gras, S.; Kotsimbos, T.C.; Mifsud, N.A. Inability to detect cross-reactive memory T cells challenges the frequency of heterologous immunity among common viruses. J. Immunol. 2018, 200, 3993–4003. [Google Scholar] [CrossRef]
- Su, Z.J.; Chen, H.B.; Zhang, J.K.; Xu, L. Effects of dendritic cells from cord blood CD34+ cells on human hepatocarcinoma cell line BEL-7402 in vitro and in SCID mice. World J. Gastroenterol. 2005, 11, 2502–2507. [Google Scholar] [CrossRef]
- Klein, U.; Dalla-Favera, R. Germinal centres: Role in B-cell physiology and malignancy. Nat. Rev. Immunol. 2008, 8, 22–33. [Google Scholar] [CrossRef]
- Kristóf, K.G.; Mácsik-Valent, B.; Matkó, J.; Bajtay, Z.; Erdei, A. Revisiting the coreceptor function of complement receptor type 2 (CR2, CD21); Co-engagement with the B-cell receptor inhibits the activation, proliferation, and antibody production of human B cells. Front. Immunol. 2021, 12, 620427. [Google Scholar] [CrossRef]
- Chumakov, K.; Avidan, M.S.; Benn, C.S.; Bertozzi, S.M.; Blatt, L.; Chang, A.Y.; Jamison, D.T.; Shabaana, A.; Khader, S.A.; Kottilil, S.; et al. Old vaccines for new infections: Exploiting innate immunity to control COVID-19 and prevent future pandemics. Proc. Natl. Acad. Sci. USA 2021, 118, e2101718118. [Google Scholar] [CrossRef]
- Lerkvaleekul, B.; Apiwattanakul, N.; Klinmalai, C.; Hongeng, S.; Vilaiyuk, S. Age-related changes in lymphocyte subpopulations in healthy Thai children. J. Clin. Lab. Anal. 2020, 34, e23156. [Google Scholar] [CrossRef]
- Trama, A.M.; Moody, A.; Alam, M.; Jaeger, F.H.; Lockwood, B.; Parks, R.; Lloyd, K.E.; Stolarchuk, C.; Scearce, R.; Foulger, A.; et al. HIV-1 envelope gp41 antibodies can originate from terminal ileum B cells that share cross-reactivity with commensal bacteria. Cell Host Microbe 2014, 16, 215–226. [Google Scholar] [CrossRef]
- Hinton, P.R.; Xiong, J.M.; Johlfs, M.G.; Tang, M.T.; Keller, S.; Tsurushita, N. An engineered human IgG1 antibody with longer serum half-life. J. Immunol. 2006, 176, 346–356. [Google Scholar] [CrossRef]
- Virgin, H.W.; Wherry, E.J.; Ahmed, R. Redefining chronic viral infection. Cell 2009, 138, 30–50. [Google Scholar] [CrossRef]
- Forrest, C.; Hislop, A.D.; Rickinson, A.B.; Zuo, B. Proteome-wide analysis of CD8+ T cell responses to EBV reveals differences between primary and persistent infection. PLoS Pathog. 2018, 14, e1007110. [Google Scholar] [CrossRef] [PubMed]
- Attaf, M.; Huseby, E.; Sewell, A.K. αβ T cell receptors as predictors of health and disease. Cell. Mol. Immunol. 2015, 12, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, M.; Abualrous, E.T.; Sticht, J.; Álvaro-Benito1, M.; Stolzenberg, S.; Noé, F.; Freund, C. Major histocompatibility complex (MHC) class I and MHC class II proteins: Conformational plasticity in antigen presentation. Front. Immunol. 2017, 8, 292. [Google Scholar] [CrossRef]
- Murdoch Children’s Research Institute. BCG Vaccination to Protect HealthcareWorkers against COVID-19 (BRACE), Clinical Trials.gov. 2021. Available online: https://clinicaltrials.gov/ct2/show/NCT04327206 (accessed on 18 April 2021).
- Aygün, D.; Önal, P.; Apaydın, G.; Çokuğraş, H. Coronavirus infections in childhood and vaccine studies. Turk. Arch. Pediatr. 2021, 56, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Abdulla, Z.A.; Al-Bashir, S.M.; Al-Salih, N.S.; Aldamen, A.A.; Abdulazeez, M.Z. A summary of the SARS-CoV-2 vaccines and technologies available or under development. Pathogens 2021, 10, 788. [Google Scholar] [CrossRef]
- Flanagan, K.L.; Best, E.; Crawford, N.W.; Giles, M.; Koirala, A.; Macartney, K.; Russell, F.; Teh, B.W.; Wen, S.C. Progress and pitfalls in the quest for effective SARS-CoV-2 (COVID-19) vaccines. Front Immunol. 2020, 11, 579250. [Google Scholar] [CrossRef]
- García-Montero, C.; Fraile-Martínez, O.; Bravo, C.; Torres-Carranza, D.; Sanchez-Trujillo, L.; Gómez-Lahoz, A.M.; Guijarro, L.G.; García-Honduvilla, N.; Asúnsolo, A.; Julia Bujan, J.; et al. An updated review of SARS-CoV-2 vaccines and the importance of effective vaccination programs in pandemic times. Vaccines 2021, 9, 433. [Google Scholar] [CrossRef] [PubMed]
- Del Barco, A.A.; Ortega, M. Epidemiology and public health in the COVID-19 epidemic. Medicine 2020, 13, 1297–1304. [Google Scholar]
- Shan Su, S.H.; Du, L.; Jiang, S.H. Learning from the past: Development of safe and effective COVID-19 vaccines. Nat. Rev. Microbiol. 2021, 19, 211–219. [Google Scholar] [CrossRef]
- Blakney, A.K.; Bekker, L. DNA vaccines join the fight against COVID-19. Lancet 2022, 399, 1281–1282. [Google Scholar] [CrossRef]
- Moore, D.L. COVID-19 Vaccine for Children and Adolescents. Canadian Paediatric Society. Available online: https://cps.ca/en/documents/position/covid-19-vaccine-for-children-and-adolescents (accessed on 25 April 2022).
- Fact Check-No Evidence COVID-19 Vaccines Have Caused INCREASE in Cancers, Contrary to Claims Made on Social Media. Available online: https://www.reuters.com/article/factcheck-cancer-covid-idUSL1N2UM24J (accessed on 20 April 2022).
- Ku, M.W.; Bourgine, M.; Authié, P.; Lopez, J.; Nemirov, K.; Moncoq, F.; Noirat, A.; Vesin, B.; Nevo, F.; Blanc, C.; et al. Intranasal vaccination with a lentiviral vector protects against SARS-CoV-2 in preclinical animal models. Cell Host Microbe 2021, 29, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Lee, H.K. Delivery Routes for COVID-19 Vaccines. Vaccines 2021, 9, 524. [Google Scholar] [CrossRef] [PubMed]
- US Food and Drugs Administration. FDA Authorizes Pfizer-BioNTech COVID-10 Vaccine for Emergency Use in Children 5 through 11 Years of Age. Available online: https://www.fda.gov/news-events/press-announcements/fda-authorizes-pfizer-biontech-covid-19-vaccine-emergency-use-children-5-through-11-years-age (accessed on 20 November 2021).
- World Health Organization. Interim Statement on COVID-19 Vaccination for Children and Adolescents. Available online: https://www.who.int/news/item/24-11-2021-interim-statement-on-covid-19-vaccination-for-children-and-adolescents (accessed on 15 December 2021).
- FDA News Release. Coronavirus (COVID-19) Update: FDA Authorizes Moderna and Pfizer-BioNTech COVID-19 Vaccines for Children Down to 6 Months of Age. Available online: https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-authorizes-moderna-and-pfizer-biontech-covid-19-vaccines-children#:~:text=For%20the%20Moderna%20COVID%2D19,years%20of%20age%20and%20older (accessed on 3 July 2022).
- Therapeutic Goods Administration of Australia. TGA Commences Evaluation of Pfizer’s COVID-19 Vaccine (COMIRNATY) for Children Aged 6 Months to Less than 5 Years. Available online: https://www.tga.gov.au/media-release/tga-commences-evaluation-pfizers-covid-19-vaccine-comirnaty-children-aged-6-months-less-5-years#:~:text=The%20Therapeutic%20Goods%20Administration%20(TGA,less%20than%205%20years%20old (accessed on 5 August 2022).
- Centers for Disease Control and Prevention (CDC). COVID-19 Vaccines for Children and Teens. Available online: https://www.cdc.gov/coronavirus/2019-ncov/vaccines/recommendations/childrenteens.html#:~:text=About%20Vaccination%20for%20Children%20and,with%20their%20COVID%2D19%20vaccines (accessed on 30 July 2022).
- Bailey, L. COVID Vaccine for Kids under 5: What Happens Next? Available online: https://news.umich.edu/covid-vaccine-for-kids-under-5-what-happens-next/ (accessed on 25 February 2022).
- Chen, C. What’s Holding Up the COVID Vaccines for Children Under 5? Available online: https://www.medscape.com/viewarticle/969603 (accessed on 7 April 2022).
- Kung, Y.; Lee, K.; Chiang, H.; Huang, S.; Wu, C.; Shih, S. Molecular virology of SARS-CoV-2 and related coronaviruses. Microbiol. Mol. Biol. Rev. 2022, 86, e0002621. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Geng, X.; Tan, Y.; Li, Q.; Xu, C.; Xu, J.; Hao, L.; Zeng, Z.; Luo, X.; Liu, F.; et al. New understanding of the damage of SARS-CoV-2 infection outside the respiratory system. Biomed. Pharmacother. 2020, 127, 110195. [Google Scholar] [CrossRef]
- Weiss, S.R.; Leibowitz, J.L. Coronavirus pathogenesis. Adv. Virus Res. 2011, 81, 85–164. [Google Scholar] [CrossRef]
- Gralinski, L.E.; Menachery, V.D. Return of the coronavirus: 2019-nCoV. Viruses 2020, 12, 135. [Google Scholar] [CrossRef]
- Wiersinga, W.J.; Rhodes, A.; Cheng, A.C.; Peacock, S.J.; Prescott, H.C. Pathophysiology, transmission, diagnosis and treatment of coronavirus disease 2019 (COVID-19). A Review. JAMA 2020, 324, 782–793. [Google Scholar] [CrossRef]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, genetic recombination, and pathogenesis of coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef]
- Moss, P. The T cell immune response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef]
- Kyriakidis, N.C.; Lopez-Cortes, A.; Gonzalez, E.V.; Grimaldos, A.B.; Prado, E.O. SARS-CoV-2 vaccines strategies: A comprehensive review of phase 3 candidates. NPJ Vaccines 2021, 6, 28. [Google Scholar] [CrossRef]
- Chao, Y.X.; Rötzschke, O.; Tan, E.K. The Role of IgA in COVID-19. Brain Behav. Immun. 2020, 87, 182–183. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Stoesser, N.; Matthews, P.C.; Ayoubkhani, D.; Studley, R.; Bell, I.; Bell, J.I.; Newton, J.N.; Jeremy Farrar, J.; Diamond, I. Antibody responses to SARS-CoV-2 vaccines in 45,965 adults from the general population of the United Kingdom. Nat. Microbiol. 2021, 6, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Islam, T.; Islam, R. New coronavirus variants are creating more challenges to global healthcare system: A brief report on the current knowledge. Clin. Pathol. 2022, 15, 2632010X221075584. [Google Scholar] [CrossRef] [PubMed]
- Planas, D.; Veyer, D.; Baidaliuk, A.; Staropoli, I.; Guivel-Benhassine, F.; Rajah, M.M.; Planchais, C.; Porrot, F.; Robillard, N.; Julien Puech, J.; et al. Reduced sensitivity of SARS-CoV-2 variant delta to antibody neutralization. Nature 2021, 596, 276–280. [Google Scholar] [CrossRef]
- Kannan, S.R.; Spratt, A.N.; Cohen, A.R.; Naqvi, S.H.; Chand, H.S.; Thomas, P.; Quinnd, T.P.; Lorsone, C.L.; Byrareddy, S.N.; Singh, K.; et al. Evolutionary analysis of the Delta and Delta plus variants of the SARS-CoV-2 viruses. J. Autoimmun. 2021, 124, 102715. [Google Scholar] [CrossRef]
- Padilla-Rojas, C.; Jimenez-Vasquez, V.; Hurtado, V.; Mestanza, O.; Molina, I.S.; Barcena, L.; Ruiz, S.M.; Wendy Lizarraga, S.A.; Henri Bailon, H.; Cáceres, O.; et al. Genomic analysis reveals a rapid spread and predominance of lambda (C.37) SARS-COV-2 lineage in Peru despite circulation of variants of concern. J. Med. Virol. 2021, 93, 6845–6849. [Google Scholar] [CrossRef]
- Kimball, S. Pfizer Covid Vaccine Was Just 12% Effective against Omicron in Kids 5 to 11, Study Finds. Available online: https://www.cnbc.com/2022/02/28/pfizer-covid-vaccine-was-just-12percent-effective-against-omicron-in-kids-5-to-11-study-finds.html (accessed on 15 March 2022).
- Collier, D.A.; Ferreira, I.A.T.M.; Kotagiri, P.; Datir, R.P.; Lim, E.Y.; Touizer, E.; Meng, B.; Abdullahi, A.; Elmer, A.; Kingston, N.; et al. Age-related immune response heterogeneity to SARS-CoV-2 vaccine BNT162b2. Nature 2021, 596, 417–422. [Google Scholar] [CrossRef]
- Mayo Clinic Staff. COVID-19 Vaccines for Kids: What You Need to Know. Available online: https://www.mayoclinic.org/diseases-conditions/coronavirus/in-depth/covid-19-vaccines-for-kids/art-20513332 (accessed on 24 July 2022).
- Kostoff, R.N.; Calina, D.; Kanduc, D.; Briggs, M.B.; Vlachoyiannopoulose, P.; Svistunovf, A.A.; Tsatsakisg, A. Why are we vaccinating children against COVID-19? Toxicol. Rep. 2021, 8, 1665–1684. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. COVID-19 Vaccines for Children and Teens. Available online: https://www.cdc.gov/coronavirus/2019-ncov/vaccines/recommendations/children-teens.html (accessed on 4 August 2022).
- Jhaveri, R.; Adler-Shohet, F.C.; Blyth, C.C.; Chiotos, K.; Gerber, J.S.; Green, M.; Kociolek, L.; Martin-Blais, R.; Palazzi, D.; Shane, A.L.; et al. Weighing the risks of perimyocarditis with the benefits of SARS-CoV-2 MRNA vaccination in adolescents. J. Pediatr. Infect. Dis. Soc. 2021, 10, piab061. [Google Scholar] [CrossRef]
- Gargano, J.W.; Wallace, M.; Hadler, S.C.; Langley, G.; Su, J.R.; Oster, M.E.; Broder, K.R.; Gee, J.; Weintraub, E.; Shimabukuro, T.; et al. Use of MRNA COVID-19 vaccine after reports of myocarditis among vaccine recipients: Update from the Advisory Committee on Immunization Practices—United States, June 2021. Mmwr. Morb. Mortal. Wkly. Rep. 2021, 70, 977–982. [Google Scholar] [CrossRef] [PubMed]
- Schauer, J.; Buddhe, S.; Colyer, J.; Sagiv, E.; Law, Y.; Chikkabyrappa, S.M.; Portman, M.A. Myopericarditis after the Pfizer MRNA COVID-19 vaccine in adolescents. J. Pediatrics 2021, 238, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Frenck, R.W.; Klein, N.P.; Kitchin, N.; Gurtman, A.; Absalon, J.; Lockhart, S.; Perez, J.L.; Walter, E.B.; Senders, S.; Bailey, R.; et al. Safety, Immunogenicity, and efficacy of the BNT162b2 COVID-19 vaccine in adolescents. N. Engl. J. Med. 2021, 385, 239–250. [Google Scholar] [CrossRef]
- Han, B.; Song, Y.; Li, C.; Yang, W.; Ma, Q.; Jiang, Z.; Li, M.; Lian, X.; Jiao, W.; Wang, L.; et al. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 Vaccine (CoronaVac) in healthy children and adolescents: A double-blind, randomised, controlled, phase 1/2 clinical trial. Lancet Infect. Dis. 2021, 21, 1645–1653. [Google Scholar] [CrossRef]
- Garrido, C.; Curtis, A.D.; Dennis, M.; Pathak, S.H.; Gao, H.; Montefiori, D.; Tomai, M.; Fox, C.B.; Kozlowski, P.A.; Scobey, T.; et al. SARS-CoV-2 vaccines elicit durable immune responses in infant rhesus macaques. Sci. Immunol. 2021, 6, eabj3684. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Finn, A.; Pollard, A.J. Should we be vaccinating children against COVID-19 in high-income countries? Expert Rev. Vaccines 2021, 20, 1043–1046. [Google Scholar] [CrossRef]
- Frenck, R.W.; Klein, N.P.; Kitchin, N.; Gurtman, A.; Absalon, J.; Lockhart, S.; Perez, J.L.; Walter, E.B.; Senders, S.; Bailey, R.; et al. Education and mental health: Good reasons to vaccinate children. Lancet 2021, 385, 239–250. [Google Scholar] [CrossRef]
- Callaway, E. COVID Vaccines and Kids: Five questions as trials begin. Nature 2021, 592, 670–671. [Google Scholar] [CrossRef]
- Ladhani, S.N. Crossing the Rubicon: A Fine line between waiting and vaccinating adolescents against COVID-19. J. Infect. 2021, 83, 294–297. [Google Scholar] [CrossRef]
- Martin, B.; DeWitt, P.E.; Russell, S.; Anand, A.; Bradwell, K.R.; Bremer, C.; Gabriel, D.; Girvin, A.T.; Hajagos, J.G.; McMurry, J.A.; et al. Children with SARS-CoV-2 in the National COVID Cohort Collaborative (N3C). medRxiv 2021. [Google Scholar] [CrossRef]
- Plotkin, S.A.; Levy, O. Considering mandatory vaccination of children for COVID-19. Pediatrics 2021, 147, e2021050531. [Google Scholar] [CrossRef]
- Archard, D.; Brierley, J.; Cave, E. Compulsory childhood vaccination: Human rights, solidarity, and best interests. Med. Law Rev. 2021, 6, 716–727. [Google Scholar] [CrossRef] [PubMed]
- Gostin, L.O.; Shaw, J.; Salmon, D.A. Mandatory SARS-CoV-2 vaccinations in K-12 schools, colleges/universities, and businesses. JAMA 2021, 326, 25–26. [Google Scholar] [CrossRef]
- Seneff, S.; Nigh, G. Worse than the disease? Reviewing some possible unintended consequences of the mRNA vaccines against COVID-19. Int. J. Vacc. Theory Pract. Res. 2021, 2, 38–79. [Google Scholar] [CrossRef]
- Suzuki, Y.J.; Gychka, S.G. SARS-CoV-2 spike protein elicits cell signaling in human host cells: Implications for possible consequences of COVID-19 vaccines. Vaccines 2021, 9, 36. [Google Scholar] [CrossRef]
- Avolio, E.; Carrabba, M.; Milligan, R.; Williamson, M.K.; Beltrami, A.P.; Gupta, K.; Elvers, K.T.; Gamez, M.; Foster, R.R.; Gillespie, K.; et al. SARS-CoV-2 Spike protein disrupts human cardiac pericytes function through CD147 receptor-mediated signalling: A potential non-infective mechanism of COVID-19 microvascular disease. Clin. Sci. 2021, 135, 2667–2689. [Google Scholar] [CrossRef] [PubMed]
- Rhea, E.M.; Logsdon, A.F.; Hansen, K.M.; Williams, L.M.; Reed, M.J.; Baumann, K.K.; Holden, S.J.; Raber, J.; Banks, W.A.; Erickson, M.A. The S1 protein of SARS-CoV-2 crosses the blood–brain barrier in mice. Nat. Neurosci. 2021, 24, 368–378. [Google Scholar] [CrossRef]
- Vojdani, A.; Vojdani, E.; Kharrazian, D. Reaction of human monoclonal antibodies to SARS-CoV-2 proteins with tissue antigens: Implications for autoimmune diseases. Front. Immunol. 2020, 11, 617089. [Google Scholar] [CrossRef]
- Ndeupen, S.; Qin, Z.; Jacobsen, S.; Estanbouli, H.; Bouteau, A.; Igyártó, B.Z. The mRNA-LNP platform’s lipid nanoparticle component used in preclinical vaccine studies is highly inflammatory. bioRxiv 2021, 24, 103479. [Google Scholar] [CrossRef]
- Sellaturay, P.; Nasser, S.; Islam, S.; Gurugama, P.; Ewan, P.W. Polyethylene glycol (PEG) is a cause of anaphylaxis to the Pfizer/BioNTech mRNA COVID-19 vaccine. Clin. Exp. Allergy 2021, 51, 861–863. [Google Scholar] [CrossRef]
- Igyártó, B.Z.; Jacobsen, S.; Ndeupen, S. Future considerations for the mRNA-lipid nanoparticle vaccine platform. Curr. Opin. Virol. 2021, 48, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Vera-Lastra, O.; Ordinola Navarro, A.; Cruz Domiguez, M.P.; Medina, G.; Sánchez Valadez, T.I.; Jara, L.J. Two cases of Graves’ disease following SARS-CoV-2 vaccination: An autoimmune/inflammatory syndrome induced by adjuvants. Thyroid 2021, 31, 1436–1439. [Google Scholar] [CrossRef] [PubMed]
- İremli, B.G.; Şendur, S.N.; Ünlütürk, U. Three cases of subacute thyroiditis following SARS-CoV-2 vaccine: Postvaccination ASIA syndrome. J. Clin. Endocrinol. Metab. 2021, 106, 2600–2605. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P.R. Thrombosis after covid-19 vaccination. BMJ 2021, 373, n958. [Google Scholar] [CrossRef]
- Merchant, H.A. CoViD vaccines and thrombotic events: EMA issued warning to patients and healthcare professionals. J. Pharm. Policy Pract. 2021, 14, 32. [Google Scholar] [CrossRef]
- Schultze, J.L.; Aschenbrenner, A.C. COVID-19 and the human innate immune System. Cell 2021, 184, 1671–1692. [Google Scholar] [CrossRef]
- Zhang, Q.; Bastard, P.; Liu, Z.; Pen, J.L.; Moncada-Velez, M.; Chen, J.; Ogishi, M.; Sabli, I.K.D.; Hodeib, S.; Korol, C.; et al. Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science 2020, 370, eabd4570. [Google Scholar] [CrossRef] [PubMed]
- Decker, M.-L.; Gotta, V.; Wellmann, S.; Ritz, N. Cytokine profiling in healthy children shows association of age with cytokine concentrations. Sci. Rep. 2017, 7, 17842. [Google Scholar] [CrossRef] [PubMed]
- Vella, L.A.; Giles, J.R.; Baxter, A.E.; Oldridge, D.A.; Diorio, C.; Kuri-Cervantes, L.; Alanio, C.; Pampena, M.B.; Wu, J.E.; Chen, Z.; et al. Deep immune profiling of MIS-C demonstrates marked but transient immune activation compared to adult and pediatric COVID-19. Sci. Immunol. 2021, 6, eabf7570. [Google Scholar] [CrossRef]
- Lu, W.; Yang, L.; Li, X.; Sun, M.; Zhang, A.; Qi, S.; Chen, Z.; Zhang, L.; Li, J.; Xiong, H. Early immune responses and prognostic factors in children with COVID-19: A Single-center retrospective analysis. BMC Pediatr. 2021, 21, 181. [Google Scholar] [CrossRef]
- Wu, H.; Zhu, H.; Yuan, C.; Yao, C.; Luo, W.; Shen, X.; Wang, J.; Shao, J.; Xiang, Y. Clinical and immune features of hospitalized pediatric patients with coronavirus disease 2019 (COVID-19) in Wuhan, China. JAMA Netw. Open 2020, 3, e2010895. [Google Scholar] [CrossRef] [PubMed]
- Vono, M.; Huttner, A.; Lemeille, S.; Martinez-Murillo, P.; Meyer, B.; Baggio, S.; Sharma, S.; Thiriard, A.; Marchant, A.; Godeke, G.-J.; et al. Robust innate responses to SARS-CoV-2 in children resolve faster than in adults without compromising adaptive immunity. Cell Rep. 2021, 37, 109773. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Rinaldi, E.; Zusi, C.; Beatrice, G.; Saccomani, M.D.; Dalbeni, A. Coronavirus disease 2019 (COVID-19) in children and/or adolescents: A meta-analysis. Pediatr. Res. 2021, 89, 733–737. [Google Scholar] [CrossRef] [PubMed]
- Neeland, M.R.; Bannister, S.; Clifford, V.; Dohle, K.; Mulholland, K.; Sutton, P.; Curtis, N.; Steer, A.C.; Burgner, D.P.; Crawford, N.W.; et al. Innate cell profiles during the acute and convalescent phase of SARS-CoV-2 infection in children. Nat. Commun. 2021, 12, 1084. [Google Scholar] [CrossRef] [PubMed]
- Seery, V.; Raiden, S.C.; Algieri, S.C.; Grisolía, N.A.; Filippo, D.; Carli, N.D.; Lalla, S.D.; Cairoli, H.; Chiolo, M.J.; Meregalli, C.N.; et al. Blood neutrophils from children with COVID-19 exhibit both inflammatory and anti-inflammatory markers. Ebiomedicine 2021, 67, 103357. [Google Scholar] [CrossRef]
- Valiathan, R.; Ashman, M.; Asthana, D. Effects of ageing on the immune system: Infants to elderly. Scand. J. Immunol. 2016, 83, 255–266. [Google Scholar] [CrossRef]
- Schouten, L.R.; van Kaam, A.H.; Kohse, F.; Veltkamp, F.; Bos, L.D.; de Beer, F.M.; van Hooijdonk, R.T.; Horn, J.; Straat, M.; Witteveen, E.; et al. Age-dependent differences in pulmonary host responses in ARDS: A prospective observational cohort study. Ann. Intensive Care 2019, 9, 55. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, S.; Hedrich, C.M. SARS-CoV-2 infections in children and young People. Clin. Immunol. 2020, 220, 108588. [Google Scholar] [CrossRef] [PubMed]
- Henry, B.M.; Benoit, S.W.; de Oliveira, M.H.S.; Hsieh, W.C.; Benoit, J.; Ballout, R.A.; Plebani, M.; Lippi, G. Laboratory abnormalities in children with mild and severe coronavirus disease 2019 (COVID-19): A Pooled analysis and review. Clin. Biochem. 2020, 81, 1–8. [Google Scholar] [CrossRef]
- Moratto, D.; Giacomelli, M.; Chiarini, M.; Savarè, L.; Saccani, B.; Motta, M.; Timpano, S.; Poli, P.; Paghera, S.; Imberti, L.; et al. Immune response in children with COVID-19 is characterized by lower levels of T-cell activation than infected adults. Eur. J. Immunol. 2020, 50, 1412–1414. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, L.; Du, H.; Zhang, J.; Li, Y.Y.; Qu, J.; Zhang, W.; Wang, Y.; Bao, S.; Li, Y.; et al. SARS-CoV-2 infection in children. N. Engl. J. Med. 2020, 382, 1663–1665. [Google Scholar] [CrossRef]
- Pierce, C.A.; Sy, S.; Galen, B.; Goldstein, D.Y.; Orner, E.P.; Keller, M.J.; Herold, K.C.; Herold, B.C. Natural Mucosal Barriers and COVID-19 in Children. JCI Insight 2021, 6, e148694. [Google Scholar] [CrossRef] [PubMed]
- Lucas, C.; Wong, P.; Klein, J.; Castro, T.B.R.; Silva, J.; Sundaram, M.; Ellingson, M.K.; Mao, T.; Oh, J.E.; Israelow, B.; et al. Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature 2020, 584, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Yonker, L.M.; Neilan, A.M.; Bartsch, Y.; Patel, A.B.; Regan, J.; Arya, P.; Gootkind, E.; Park, G.; Hardcastle, M.; John, A.S.; et al. Pediatric Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): Clinical Presentation, Infectivity, and Immune Responses. J. Pediatr. 2020, 227, 45–52.e5. [Google Scholar] [CrossRef]
- Chakraborty, S.; Gonzalez, J.; Edwards, K.; Mallajosyula, V.; Buzzanco, A.S.; Sherwood, R.; Buffone, C.; Kathale, N.; Providenza, S.; Xie, M.M.; et al. Proinflammatory IgG Fc structures in patients with severe COVID-19. Nat. Immunol. 2021, 22, 67–73. [Google Scholar] [CrossRef]
- Liu, P.; Cai, J.; Jia, R.; Xia, S.; Wang, X.; Cao, L.; Zeng, M.; Xu, J. Dynamic surveillance of SARS-CoV-2 shedding and neutralizing antibody in children with COVID-19. Emerg. Microbes Infect. 2020, 9, 1254–1258. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, J.; Jia, R.; Yi, C.; Gu, W.; Liu, P.; Dong, X.; Zhou, H.; Shang, B.; Cheng, S.; et al. Protective humoral immunity in SARS-CoV-2 infected pediatric patients. Cell Mol. Immunol. 2020, 17, 768–770. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; Connors, T.J.; Zhu, Y.; Baldwin, M.R.; Lin, W.-H.; Wontakal, S.; Szabo, P.A.; Wells, S.B.; Dogra, P.; Gray, J.; et al. Distinct antibody responses to SARS-CoV-2 in children and adults across the COVID-19 clinical spectrum. Nat. Immunol. 2021, 22, 25–31. [Google Scholar] [CrossRef]
- Hachim, A.; Gu, H.; Kavian, O.; Kwan, M.Y.; Chan, W.; Yau, Y.S.; Chiu, S.S.; Tsang, O.T.; Hui, D.S.; Ma, F.; et al. The SARS-CoV-2 antibody landscape is lower in magnitude for structural proteins, diversified for accessory proteins and stable long-term in children. medRxiv 2021. [Google Scholar] [CrossRef]
- Bartsch, Y.C.; Wang, C.; Zohar, T.; Fischinger, S.; Atyeo, C.; Burke, J.S.; Kang, J.; Edlow, A.G.; Fasano, A.; Baden, L.R.; et al. Humoral signatures of protective and pathological SARS-CoV-2 infection in children. Nat. Med. 2021, 27, 454–462. [Google Scholar] [CrossRef]
- Cotugno, N.; Ruggiero, A.; Bonfante, F.; Petrara, M.R.; Zicari, S.; Pascucci, G.R.; Zangari, P.; Ioris, M.A.D.; Santilli, V.; Manno, E.C.; et al. Virological and immunological features of SARS-CoV-2-infected children who develop neutralizing antibodies. Cell Rep. 2021, 34, 108852. [Google Scholar] [CrossRef]
- Chen, Y.; Zuiani, A.; Fischinger, S.; Mullur, J.; Atyeo, C.; Travers, M.; Lelis, F.J.N.; Pullen, K.M.; Martin, H.; Tong, P.; et al. Quick COVID-19 healers sustain anti-SARS-CoV-2 antibody production. Cell 2020, 183, 1496–1507.e16. [Google Scholar] [CrossRef]
- Bloise, S.; Marcellino, A.; Testa, A.; Dilillo, A.; Mallardo, S.; Isoldi, S.; Martucci, V.; Sanseviero, M.T.; Giudice, E.D.; Iorfida, D.; et al. Serum IgG levels in children 6 months after SARS-CoV-2 infection and comparison with adults. Eur. J. Pediatr. 2021, 180, 3335–3342. [Google Scholar] [CrossRef]
- Garrido, C.; Hurst, J.H.; Lorang, C.G.; Aquino, J.N.; Rodriguez, J.; Pfeiffer, T.S.; Singh, T.; Semmes, E.C.; Lugo, D.J.; Rotta, A.T.; et al. Asymptomatic or mild symptomatic SARS-CoV-2 infection elicits durable neutralizing antibody responses in children and adolescents. JCI Insight 2021, 6, e148694. [Google Scholar] [CrossRef]
- Cohen, C.A.; Li, A.P.; Hachim, A.; Hui, D.S.; Kwan, M.Y.; Tsang, O.T.; Chiu, S.S.; Chan, W.H.; Yau, Y.S.; Kavian, N.; et al. SARS-CoV-2 specific T cell responses are lower in children and increase with age and time after infection. medRxiv 2021. [Google Scholar] [CrossRef]
- Erdeve, Ö.; Çetinkaya, M.; Baş, A.Y.; Narli, N.; Duman, N.; Vural, M.; Koç, E. The Turkish Neonatal Society proposal for the management of COVID-19 in the neonatal intensive care unit. Turk. Arch. Pediatr. 2020, 55, 86–92. [Google Scholar] [CrossRef]
- Çokuğraş, H.; Önal, P. SARS-CoV-2 infection in children. Turk. Arch. Pediatr. 2020, 55, 95–102. [Google Scholar] [CrossRef]
- Ludvigsson, J.F. Systematic review of COVID-19 in children shows milder cases and a better prognosis than adults. Acta Paediatr. 2020, 109, 1088–1095. [Google Scholar] [CrossRef]
- Davies, N.G.; Klepac, P.; Liu, Y.; Prem, K.; Jit, M.; CMMID COVID-19 Working Group; Eggo, R.M. Age-dependent effects in the transmission and control of COVID-19 epidemics. Nat. Med. 2020, 26, 1205–1211. [Google Scholar] [CrossRef]
- Bhumbra, S.; Malin, S.; Kirkpatrick, L.; Khaitan, A.; John, C.C.; Rowan, C.M.; Enane, L.A. Clinical features of critical coronavirus disease 2019 in children. Pediatr. Crit. Care Med. 2020, 21, 948–953. [Google Scholar] [CrossRef]
- Dong, Y.; Xi, M.; Hu, Y.; Qi, X.; Jiang, F.; Jiang, Z.; Tong, S. Epidemiological characteristics of 2143 pediatric patients with 2019 coronavirus disease in China. Pediatrics 2020, 145, e20200702. [Google Scholar] [CrossRef]
- Zimmermann, P.; Curtis, N. Coronavirus infections in children including COVID-19: An overview of the epidemiology, clinical features, diagnosis, treatment and prevention options in children. Pediatr. Infect. Dis. J. 2020, 39, 355–368. [Google Scholar] [CrossRef]
- Sinha, S.; Kunducorresponding, C.N. Cancer and COVID-19: Why are cancer patients more susceptible to COVID-19? Med. Oncol. 2021, 38, 101. [Google Scholar] [CrossRef]
- Gadotti, A.C.; de Castro Deus, M.; Telles, J.P.; Wind, R.; Goes, M.; Garcia Charello Ossoski, R.; de Padua, A.M.; de Noronha, L.; Moreno-Amaral, A.; Baena, C.P.; et al. IFN-γ is an independent risk factor associated with mortality in patients with moderate and severe COVID-19 infection. Virus Res. 2020, 289, 198171. [Google Scholar] [CrossRef]
- Sun, X.; Wang, T.; Cai, D.; Hu, Z.; Chen, J.; Liao, H.; Zhi, L.; Wei, H.; Zhang, Z.; Qiu, Y.; et al. Cytokine storm intervention in the early stages of COVID-19 pneumonia. Cytokine Growth Factor Rev. 2020, 53, 38–42. [Google Scholar] [CrossRef]
- Hu, B.; Huang, S.; Yin, L. The cytokine storm and COVID-19. J. Med. Virol. 2020, 93, 250–256. [Google Scholar] [CrossRef]
- Carsetti, R.; Zaffina, S.; Mortari, P.E.; Terreri, S.; Corrente, F.; Capponi, C.; Palomba, P.; Mirabella, M.; Cascioli, S.; Palange, P.; et al. Different innate and adaptive immune responses to SARS-CoV-2 infection of asymptomatic, mild, and severe cases. Front. Immunol. 2020, 11, 610300. [Google Scholar] [CrossRef]
- Bordallo, B.; Bellas, M.; Cortez, A.F.; Vieira, M.; Pinheiro, M. Severe COVID-19: What have we learned with the immunopathogenesis? Adv. Rheumatol. 2020, 60, 50. [Google Scholar] [CrossRef]
- Gkoutzourelas, A.; Bogdanos, D.P.; Sakkas, L.I. Kawasaki Disease and COVID-19. Mediterr. J. Rheumatol. 2020, 31, 268–274. [Google Scholar] [CrossRef]
- Bordoni, V.; Sacchi, A.; Cimini, E.; Notari, S.; Grassi, G.; Tartaglia, E.; Casetti, R.; Giancola, M.L.; Bevilacqua, N.; Maeurer, M.; et al. An Inflammatory Profile Correlates With Decreased Frequency of Cytotoxic Cells in Coronavirus Disease 2019. Clin. Infect. Dis. 2020, 71, 2272–2275. [Google Scholar] [CrossRef]
- Gruber, C.N.; Patel, R.S.; Trachtman, R.; Lepow, L.; Amanat, F.; Krammer, F.; Wilson, K.M.; Onel, K.; Geanon, D.; Tuballes, K.; et al. Mapping Systemic Inflammation and Antibody Responses in Multisystem Inflammatory Syndrome in Children (MIS-C). Cell 2020, 183, 982–995.e14. [Google Scholar] [CrossRef]
- Miri, S.M.; Noorbakhsh, F.; Mohebbi, S.R.; Ghaemi, A. Higher prevalence of asymptomatic or mild COVID-19 in children, claims and clues. J. Med. Virol. 2020, 92, 2257–2259. [Google Scholar] [CrossRef]
- Dhochak, N.; Singhal, T.; Kabra, S.K.; Lodha, R. Pathophysiology of COVID-19: Why Children Fare Better than Adults? Indian J. Pediatr. 2020, 87, 537–546. [Google Scholar] [CrossRef]
- Hedrich, C.M. COVID 19-consederation for the paediatric rheumatologist. Clin. Immunol. 2020, 214, 108420. [Google Scholar] [CrossRef]
- Li, H.; Chen, K.; Liu, M.; Xu, H.; Xu, Q. The profile of peripheral blood lymphocyte subsets and serum cytokines in children with 2019 novel coronavirus pneumonia. J. Infect. 2020, 81, 115–120. [Google Scholar] [CrossRef]
- Jacques, F.H.; Apedaile, E. Immunopathogenesis of COVID-19: Summary and Possible Interventions. Front. Immunol. 2020, 11, 564925. [Google Scholar] [CrossRef]
- Liu, J.; Li, S.; Liu, J.; Liang, B.; Wang, X.; Wang, H.; Li, W.; Tong, Q.; Yi, J.; Zhao, L.; et al. Longitudinal characteristics of lymphocyte responses and cytokine profiles in the peripheral blood of SARS-CoV-2 infected patients. EBioMedicine 2020, 55, 102763. [Google Scholar] [CrossRef]
- Ravichandran, S.; Tang, J.; Grubbs, G.; Lee, Y.; Pourhashemi, S.; Hussaini, L.; Lapp, S.A.; Jerris, R.C.; Singh, V.; Chahroudi, A.; et al. SARS-CoV-2 immune repertoire in MIS-C and pediatric COVID-19. Nat. Immunol. 2021, 22, 1452–1464. [Google Scholar] [CrossRef]
- Perez-Toledo, M.; Faustini, S.E.; Jossi, S.E.; Shields, A.M.; Kanthimathinathan, H.K.; Allen, J.D.; Watanabe, Y.; Goodall, M.; Wraith, D.C.; Tonny, V.; et al. Serology confirms SARS-CoV-2 infection in PCR-negative children presenting with paediatric inflammatory multi-system syndrome. medRxiv 2020, 32, 1125–1129. [Google Scholar] [CrossRef]
- Pierce, C.A.; Preston-Hurlburt, P.; Dai, Y.; Aschner, C.B.; Cheshenko, N.; Galen, B.; Garforth, S.J.; Herrera, N.G.; Jangra, R.K.; Morano, N.C.; et al. Immune responses to SARS-CoV-2 infection in hospitalized pediatric and adult patients. Sci. Transl. Med. 2020, 12, eabd5487. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Gaebler, C.; Wang, Z.; Lorenzi, J.C.C.; Muecksch, F.; Finkin, S.; Tokuyama, M.; Cho, A.; Jankovic, M.; Schaefer-Babajew, D.; Oliveira, T.Y.; et al. Evolution of antibody immunity to SARS-CoV-2. Nature 2021, 591, 639–644. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e1415. [Google Scholar] [CrossRef]
- Zhu, J.; Ji, P.; Pang, J.; Zhong, Z.; Li, H.; He, C.; Zhang, J.; Zhao, C. Clinical characteristics of 3062 COVID-19 patients: A meta-analysis. J. Med. Virol. 2020, 92, 1902–1914. [Google Scholar] [CrossRef]
- Nanda, A.; Vura, N.V.R.K.; Gravenstein, S. COVID-19 in older adults. Aging Clin. Exp. Res. 2020, 32, 1199–1202. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Rao, N.M.; Goenka, A.; Roderick, M.; Ramanan, A.V. Coronavirus Disease 2019 (COVID-19) in Children—What We Know So Far and What We Do Not. Indian Pediatr. 2020, 57, 435–442. [Google Scholar] [CrossRef]
- Han, H.; Ma, Q.; Li, C.; Liu, R.; Zhao, L.; Wang, W.; Zhang, P.; Liu, X.; Gao, G.; Liu, F.; et al. Profiling serum cytokines in COVID-19 patients reveals IL-6 and IL-10 are disease severity predictors. Emerg. Microbes Infect. 2020, 9, 1123–1130. [Google Scholar] [CrossRef]
- Warner, S.; Richter, A.; Stamataki, Z.; Kelly, D. Understanding COVID-19: Are children the key? BMJ Paediatr. Open 2021, 5, e001063. [Google Scholar] [CrossRef]
- Riphagen, S.; Gomez, X.; Gonzalez-Martinez, C.; Wilkinson, N.; Theocharis, P. Hyperinflammatory shock in children during COVID-19 pandemic. Lancet 2020, 395, 1607–1608. [Google Scholar] [CrossRef]
- Toubiana, J.; Poirault, C.; Corsia, A.; Bajolle, F.; Fourgeaud, J.; Angoulvant, F.; Debray, A.; Basmaci, R.; Salvador, E.; Biscardi, S.; et al. Outbreak of Kawasaki disease in children during COVID-19 pandemic: A prospective observational study in Paris, France. medRxiv 2020, 369, m2094. [Google Scholar] [CrossRef]
- Verdoni, L.; Mazza, A.; Gervasoni, A.; Martelli, L.; Ruggeri, M.; Ciuffreda, M.; Bonanomi, E.; D’Antiga, L. An outbreak of severe Kawasaki-like disease at the Italian epicentre of the SARS-CoV-2 epidemic: An observational cohort study. Lancet 2020, 395, 1771–1778. [Google Scholar] [CrossRef]
- Whittaker, E.; Bamford, A.; Kenny, J.; Kaforou, M.; Jones, C.; Shah, P.; Ramnarayan, P.; Fraisse, A.; Miller, O.; Davies, P.; et al. Clinical Characteristics of 58 Children With a Pediatric Inflammatory Multisystem Syndrome Temporally Associated With SARS-CoV-2. JAMA 2020, 324, 259–269. [Google Scholar] [CrossRef]
- Consiglio, C.R.; Cotugno, N.; Sardh, F.; Pou, C.; Amodio, D.; Rodriguez, L.; Tan, Z.; Zicari, S.; Ruggiero, A.; Pascucci, G.R.; et al. The Immunology of Multisystem Inflammatory Syndrome in Children with COVID-19. Cell 2020, 183, 968–981.e7. [Google Scholar] [CrossRef]
- Rodriguez, L.; Pekkarinen, P.T.; Lakshmikanth, T.; Tan, Z.; Consiglio, C.R.; Pou, C.; Chen, Y.; Mugabo, C.H.; Nguyen, N.A.; Nowlan, K.; et al. Systems-Level Immunomonitoring from Acute to Recovery Phase of Severe COVID-19. Cell Rep. Med. 2020, 1, 100078. [Google Scholar] [CrossRef]
- McArdle, A.J.; Vito, O.; Patel, H.; Seaby, E.G.; Shah, P.; Wilson, C.; Broderick, C.; Nijman, R.; Tremoulet, A.H.; Munblit, D.; et al. Treatment of Multisystem Inflammatory Syndrome in Children. N. Engl. J. Med. 2021, 385, 11–22. [Google Scholar] [CrossRef]
- Cheung, E.W.; Zachariah, P.; Gorelik, M.; Boneparth, A.; Kernie, S.G.; Orange, J.S.; Milner, J.D. Multisystem Inflammatory Syndrome Related to COVID-19 in Previously Healthy Children and Adolescents in New York City. JAMA 2020, 324, 294–296. [Google Scholar] [CrossRef]
- Ramaswamy, A.; Brodsky, N.N.; Sumida, T.S.; Comi, M.; Asashima, H.; Hoehn, K.B.; Li, N.; Liu, Y.; Shah, A.; Ravindra, N.G.; et al. Immune dysregulation and autoreactivity correlate with disease severity in SARS-CoV-2-associated multisystem inflammatory syndrome in children. Immunity 2021, 54, 1083–1095.e7. [Google Scholar] [CrossRef]
- Wang, E.Y.; Mao, T.; Klein, J.; Dai, Y.; Huck, J.D.; Jaycox, J.R.; Liu, F.; Zhou, T.; Israelow, B.; Wong, P.; et al. Diverse Functional Autoantibodies in Patients with COVID-19. Nature 2021, 595, 283–288. [Google Scholar] [CrossRef]
- Cheng, M.H.; Zhang, S.; Porritt, R.A.; Rivas, M.N.; Paschold, L.; Willscher, E.; Binder, M.; Arditi, M.; Bahar, I. Superantigenic character of an insert unique to SARS-CoV-2 spike supported by skewed TCR repertoire in patients with hyperinflammation. Proc. Natl. Acad. Sci. USA 2020, 117, 25254–25262. [Google Scholar] [CrossRef]
- Hoste, L.; Roels, L.; Naesens, L.; Bosteels, V.; Vanhee, S.; Dupont, S.; Bosteels, C.; Browaeys, R.; Vandamme, N.; Verstaen, K.; et al. TIM3+TRBV11-2 T cells and IFNγ signature in patrolling monocytes and CD16+ NK cells delineate MIS-C. J. Exp. Med. 2021, 219, e20211381. [Google Scholar] [CrossRef]
- Moreews, M.; Le Gouge, K.; Khaldi-Plassart, S.; Pescarmona, R.; Mathieu, A.-L.; Malcus, C.; Djebali, S.; Bellomo, A.; Dauwalder, O.; Perret, M.; et al. Polyclonal expansion of TCR Vb 21.3 + CD4 + and CD8 + T cells is a hallmark of multisystem inflammatory syndrome in children. Sci. Immunol. 2021, 6, eabh1516. [Google Scholar] [CrossRef]
- Porritt, R.A.; Paschold, L.; Rivas, M.N.; Cheng, M.H.; Yonker, L.M.; Chandnani, H.; Lopez, M.; Simnica, D.; Schultheiß, C.; Santiskulvong, C.; et al. HLA class I-associated expansion of TRBV11-2 T cells in Multisystem Inflammatory Syndrome in Children. J. Clin. Investig. 2021, 131, e146614. [Google Scholar] [CrossRef]
- Sancho-Shimizu, V.; Brodin, P.; Cobat, A.; Biggs, C.M.; Toubiana, J.; Lucas, C.L.; Henrickson, S.E.; Belot, A.; Haddad, E.; Beland, K.; et al. SARS-CoV-2–related MIS-C: A key to the viral and genetic causes of Kawasaki disease? J. Exp. Med. 2021, 218, e20210446. [Google Scholar] [CrossRef]
- Yonker, L.M.; Gilboa, T.; Ogata, A.F.; Senussi, Y.; Lazarovits, R.; Boribong, B.P.; Bartsch, Y.C.; Loiselle, M.; Rivas, M.N.; Porritt, R.A.; et al. Multisystem inflammatory syndrome in children is driven by zonulin-dependent loss of gut mucosal barrier. J. Clin. Investig. 2021, 131, e149633. [Google Scholar] [CrossRef]
- Morparia, K.; Park, M.J.; Kalyanaraman, M.; McQueen, D.; Bergel, M.; Phatak, T. Abdominal Imaging Findings in Critically Ill Children With Multisystem Inflammatory Syndrome Associated With COVID-19. Pediatr. Infect. Dis. J. 2020, 40, e82–e83. [Google Scholar] [CrossRef]
- Ramcharan, T.; Nolan, O.; Lai, C.Y.; Prabhu, N.; Krishnamurthy, R.; Richter, A.G.; Jyothish, D.; Kanthimathinathan, H.K.; Welch, S.B.; Hackett, S.; et al. Paediatric Inflammatory Multisystem Syndrome: Temporally Associated with SARS-CoV-2 (PIMS-TS): Cardiac Features, Management and Short-Term Outcomes at a UK Tertiary Paediatric Hospital. Pediatr. Cardiol. 2020, 41, 1391–1401. [Google Scholar] [CrossRef]
- Yasuhara, J.; Kuno, T.; Takagi, H.; Sumitomo, N. Clinical characteristics of COVID-19 in children: A systematic review. Pediatr. Pulmonol. 2020, 55, 2565–2575. [Google Scholar] [CrossRef]
- Ciaglia, E.; Vecchione, C.; Puca, A.A. COVID-19 infection and circulating ACE2 levels: Protective role in women and children. Front. Pediatr. 2020, 8, 11–13. [Google Scholar] [CrossRef]
- Brodin, P. SARS-CoV-2 infections in children: Understanding diverse outcomes. Immunity 2022, 55, 201–209. [Google Scholar] [CrossRef]
- Leidman, E.; Duca, L.M.; Omura, J.D.; Proia, K.; Stephens, J.W.; Sauber-Schatz, E.K. COVID-19 trends among persons aged 0–24 Years. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 88–94. [Google Scholar] [CrossRef]
- Cotugno, N.; Ruggiero, A.; Pascucci, G.R.; Bonfante, F.; Petrara, M.R.; Pighi, C.; Cifaldi, L.; Zangari, P.; Bernardi, S.; Cursi, L.; et al. Virological and immunological features of SARS-COV-2 infected children with distinct symptomatology. Pediatr. Allergy Immunol. 2021, 32, 1833–1842. [Google Scholar] [CrossRef]
- Shen, K.; China National Clinical Research Center for Respiratory Diseases; Yang, Y.; Wang, T.; Zhao, D.; Jiang, Y.; Jin, R.; Zheng, Y.; Xu, B.; Xie, Z.; et al. Diagnosis, treatment, and prevention of 2019 novel coronavirus infection in children: Experts’ consensus statement. World J. Pediatr. 2020, 16, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Aranburu, A.; Mortari, E.P.; Baban, A.; Giorda, E.; Cascioli, S.; Marcellini, V.; Scarsella, M.; Ceccarelli, S.; Corbelli, S.; Cantarutti, N.; et al. Human B-cell memory is shaped by age- and tissue-specific T-independent and GC-dependent events. Eur. J. Immunol. 2016, 47, 327–344. [Google Scholar] [CrossRef] [PubMed]
- Victora, G.D.; Nussenzweig, M.C. Germinal Centers. Annu. Rev. Immunol. 2022, 40, 413–442. [Google Scholar] [CrossRef] [PubMed]
- Holodick, N.E.; Rodríguez-Zhurbenko, N.; Hernández, A.M. Defining Natural Antibodies. Front. Immunol. 2017, 8, 872. [Google Scholar] [CrossRef]
- Reyneveld, I.; Savelkoul, H.F.J.; Parmentier, H.K. Current Understanding of Natural Antibodies and Exploring the Possibilities of Modulation Using Veterinary Models. A Review. Front. Immunol. 2020, 11, 02139. [Google Scholar] [CrossRef]
- Grimsholm, O.; Mortari, E.P.; Davydov, A.N.; Shugay, M.; Obraztsova, A.S.; Bocci, C.; Marasco, E.; Marcellini, V.; Aranburu, A.; Farroni, C.; et al. The Interplay between CD27dull and CD27bright B Cells Ensures the Flexibility, Stability, and Resilience of Human B Cell Memory. Cell Rep. 2020, 30, 2963–2977.e6. [Google Scholar] [CrossRef]
- Mauri, C.; Menon, M. Human regulatory B cells in health and disease: Therapeutic potential. J. Clin. Investig. 2017, 127, 772–779. [Google Scholar] [CrossRef]



| Neonates and Infants | Children to Young Adults | Information |
|---|---|---|
| Innate Immunity | Innate Immunity | References |
| Poor neutrophil functions and muted innate immunity at birth | More mature immunity (fewer infections and effective vaccination) | [21,22,23] |
| Reduced M/M functions | Waned maternal Ig | [24,25,26,27] |
| Reduced bioactive molecules and tissue repair | Normal bioactive molecule production and tissue repair | [28] |
| Fewer mDCs | Normal mDcs | [29] |
| Reduced PcDcs (limited α/β interferon) | Normal PcDcs | [30] |
| Low concentration of IL-12 | Normal IL-12 | [31] |
| Normal TLR4, -7, and -9 | Normal LTR receptors | [30] |
| Reduced NK activity (50% of adult level) | Increased NK activity | [32] |
| Diminished concentration of alternative/lectin-binding complement pathways | Normal alternative/lectin-binding complementary pathways | [33] |
| Adaptive Immunity | Adaptive Immunity | |
| Maintained fetal Treg cells (self-tolerance) | Decreased Treg activity | [34,35,36,37] |
| Predominant Th2 response | Strong CD4 and CD8 activity | [38] |
| Enhanced γδ T cells (increased γ interferon production) | Increased CD45R naïve T cells | [39,40] |
| B1 cell stimulation (increased interferon production) | B cell hypermutation and nonexposure cross-reactions | [41,42] |
| B2 cell stimulation (IgM production) and short-lived plasma cells and Ig | Increased Ig production, increased plasma cells in the BM and retained antigens at the follicular DC | [41,43,44] |
| Diminished Th1 and CD8 activity, enhanced T17 activity | Increased Th1, Th2, and Th17 numbers and activity | [45,46,47,48] |
| Limited generation of B and T memory cells (limited hypermutation) | Generation of B and T memory cells, effective vaccination | [49,50,51,52,53,54,55,56,57,58] |
| Limited involvement of the classical complement pathway (low CD21) | Increased classical complementary pathway activity | [59] |
| Immunocompetent Cells | Immunological Elements | Functions |
|---|---|---|
| Dendritic cells (DCs) | Plasmacytoid (pDC) and myeloid (mDC) | - Antigen-presenting cells |
| - mDC links innate immunity with adaptive immunity | ||
| - pDC produces INF-α and INF-β upon viral exposure and expresses LTR7/9 [138,140] | ||
| Neutrophils | Phagocytic cells | - Neutrophil activation is lower in pediatric COVID-19 [141] |
| Monocyte–macrophages | Phagocytic cells | - Antigen-presenting cells to T cells |
| - Recruitment of lymphocytes (elements of | ||
| adaptive immunity) | ||
| - Production of varieties of bioactive molecules | ||
| - Activation of monocyte–macrophages enhanced via trained immunity [138,139,140] | ||
| Natural killer (NK) cell | Granzymes, perforin, IFN-γ, TNF-α, and FASL expression | - Cytolytic granule-mediated cell apoptosis |
| - Antibody-dependent cell-mediated cytotoxicity (ADCC) | ||
| - Cytokine-induced NK and cytotoxic T lymphocyte (CTL) activation | ||
| - Killing of virally infected cells (missing “self” hypothesis) | ||
| - Brief decrease in NK cells after the onset of COVID-19 symptoms [138,140] | ||
| Natural antibodies | Antibodies produced before viral infection or immunization | - Activation of the classical complement pathway |
| - Antimicrobial activities [95,96] | ||
| Alternative (properdin) complement pathway | C3 to C9 and other factors | - Part of the innate immunity |
| - Does not require antibodies for activation [95,96] | ||
| Epithelial and mucous membrane barriers | Mechanical barrier with protective armory | - Prevents microbial entrance |
| - Virus must attach to it before entrance [95,138] |
| Adults | Children | |||||
|---|---|---|---|---|---|---|
| Severe | Mild | Asymptomatic | PIMS-TS | Severe | Mild | Asymptomatic |
| ↑ IL-6 | ↑ IL-6 | ↔ IL-6 | ↑ IL-6 | ↔ IL-6 | ↔ IL-6 | ↔ IL-6 |
| ↑ IL-10 | ↑ IL-10 | ↑ IL-10 | ↑ IL-10 | ↑ IL-10 | ↑ IL-10 | ↑ IL-10 |
| ↑ IFN | ↑ IFN | ↔ IFN | ↑ IFN | ↑/↔ IFN | ↑/↔ IFN | ↔ IFN |
| ↑ TNF-α | ↑ TNF-α | ↔ TNF-α | ↑ TNF-α | ↔ TNF-α | ↔ TNF-α | ↔ TNF-α |
| ↑/↔ IL-1β | ↑/↔ IL-1β | ↔ IL-1β | n/a | ↔ IL-1β | ↔ IL-1β | ↔ IL-1β |
| ↑/↔ IL-8 | ↑/↔ IL-8 | ↔ IL-8 | ↑ IL-8 | ↑ IL-8 | ↔ IL-8 | ↔ IL-8 |
| ↑/↔ IL-17 | ↑/↔ IL-17 | ↔ IL-17 | ↑ IL-17 | ↑/↔ IL-17 | ↑/↔ IL-17 | ↔ IL-17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdulla, Z.A.; Al-Bashir, S.M.; Alzoubi, H.; Al-Salih, N.S.; Aldamen, A.A.; Abdulazeez, A.Z. The Role of Immunity in the Pathogenesis of SARS-CoV-2 Infection and in the Protection Generated by COVID-19 Vaccines in Different Age Groups. Pathogens 2023, 12, 329. https://doi.org/10.3390/pathogens12020329
Abdulla ZA, Al-Bashir SM, Alzoubi H, Al-Salih NS, Aldamen AA, Abdulazeez AZ. The Role of Immunity in the Pathogenesis of SARS-CoV-2 Infection and in the Protection Generated by COVID-19 Vaccines in Different Age Groups. Pathogens. 2023; 12(2):329. https://doi.org/10.3390/pathogens12020329
Chicago/Turabian StyleAbdulla, Zainalabideen A., Sharaf M. Al-Bashir, Hiba Alzoubi, Noor S. Al-Salih, Ala A. Aldamen, and Ahmed Z. Abdulazeez. 2023. "The Role of Immunity in the Pathogenesis of SARS-CoV-2 Infection and in the Protection Generated by COVID-19 Vaccines in Different Age Groups" Pathogens 12, no. 2: 329. https://doi.org/10.3390/pathogens12020329
APA StyleAbdulla, Z. A., Al-Bashir, S. M., Alzoubi, H., Al-Salih, N. S., Aldamen, A. A., & Abdulazeez, A. Z. (2023). The Role of Immunity in the Pathogenesis of SARS-CoV-2 Infection and in the Protection Generated by COVID-19 Vaccines in Different Age Groups. Pathogens, 12(2), 329. https://doi.org/10.3390/pathogens12020329