


Taenia crassiceps Cysticercosis in a Wild Muskrat and a Domestic Dog in the Northeastern United States

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. The Muskrat Case
2.2. The Dog Case
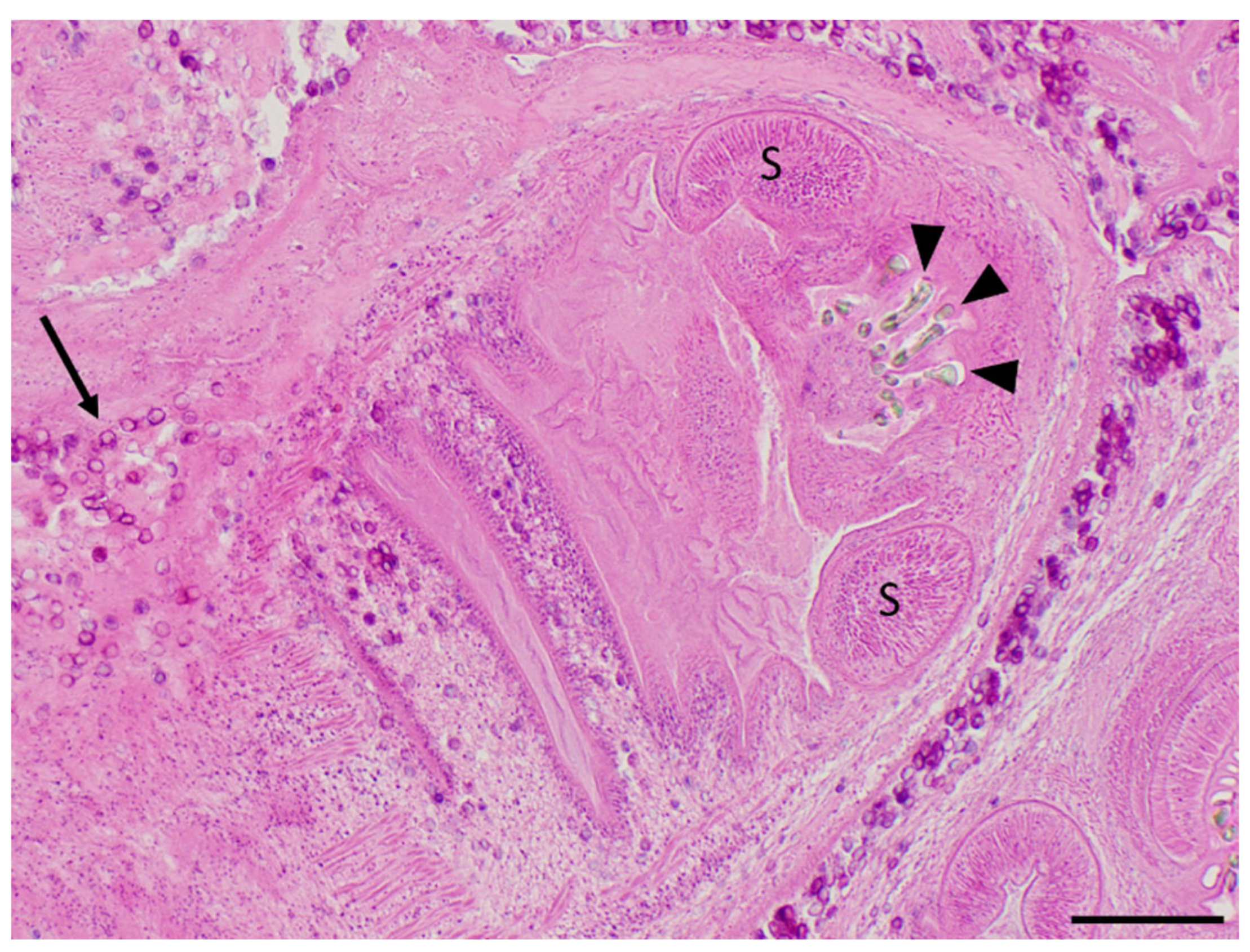
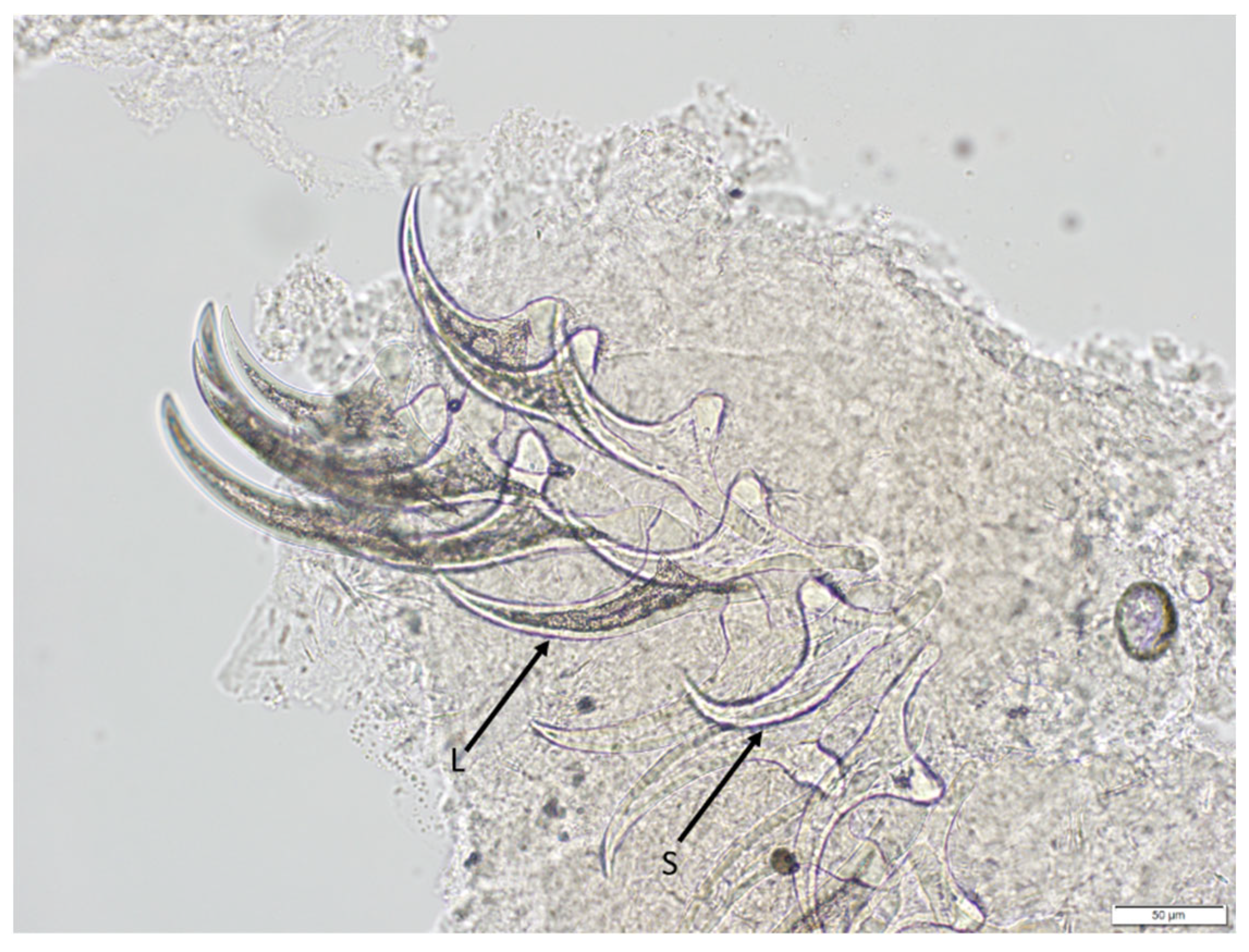
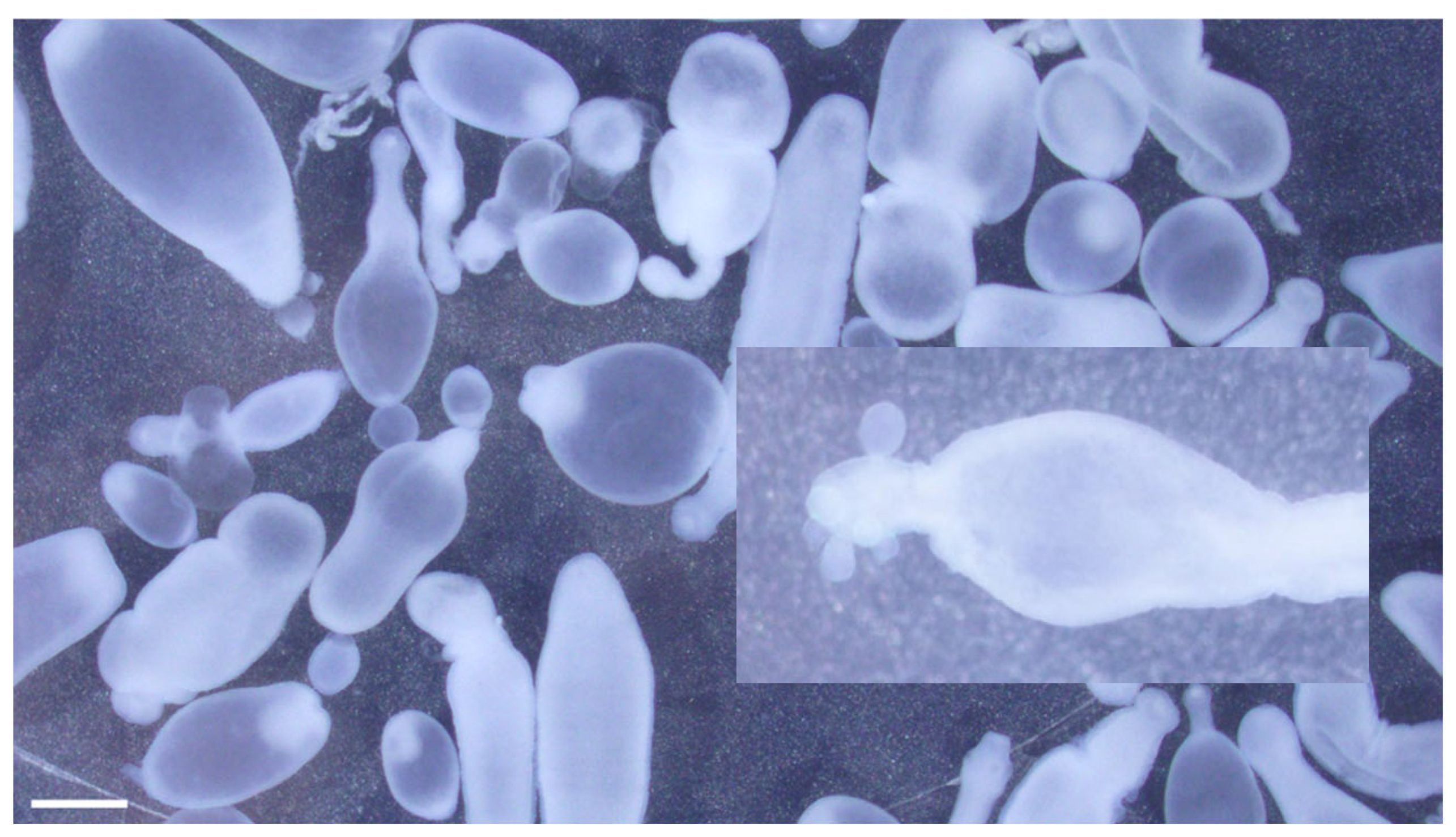
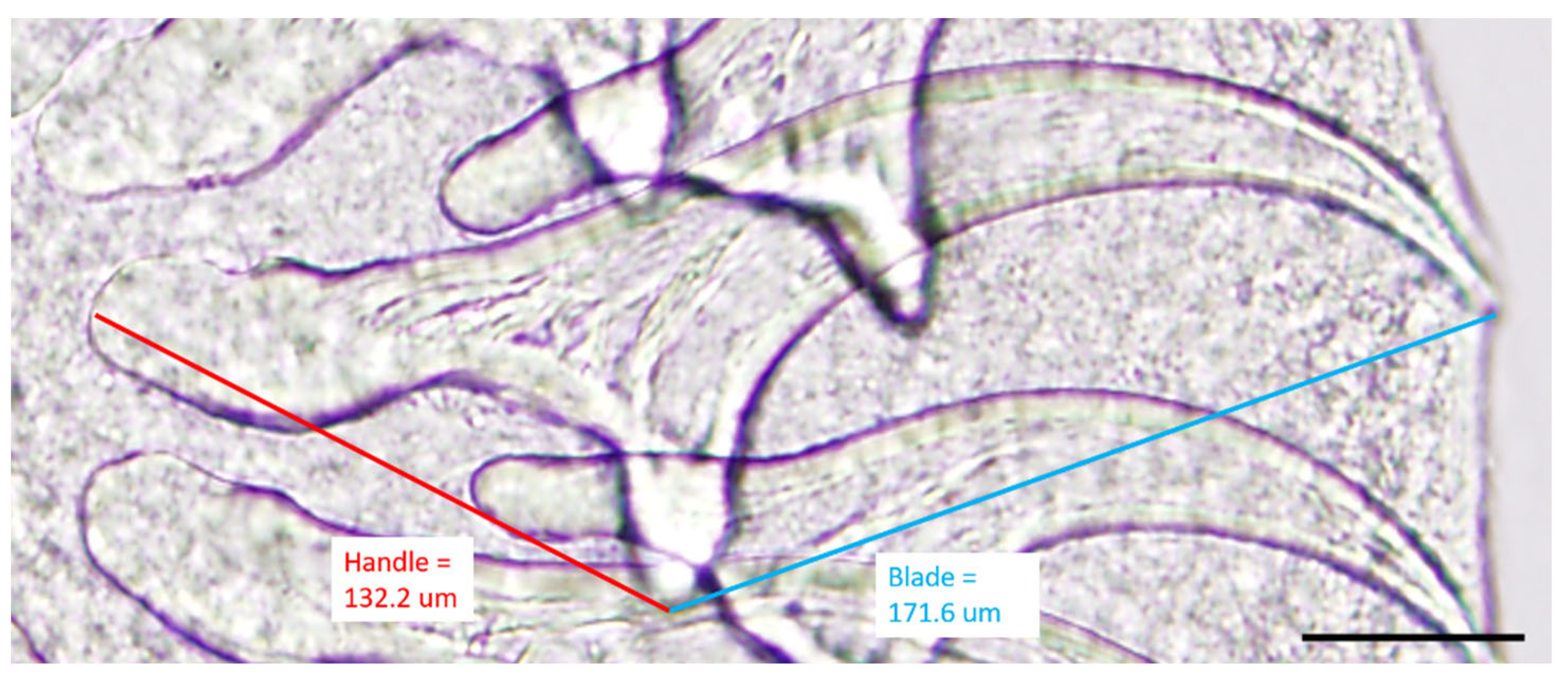
2.3. Morphological Identification: Microscopy
2.4. Molecular Identification: PCR
3. Results
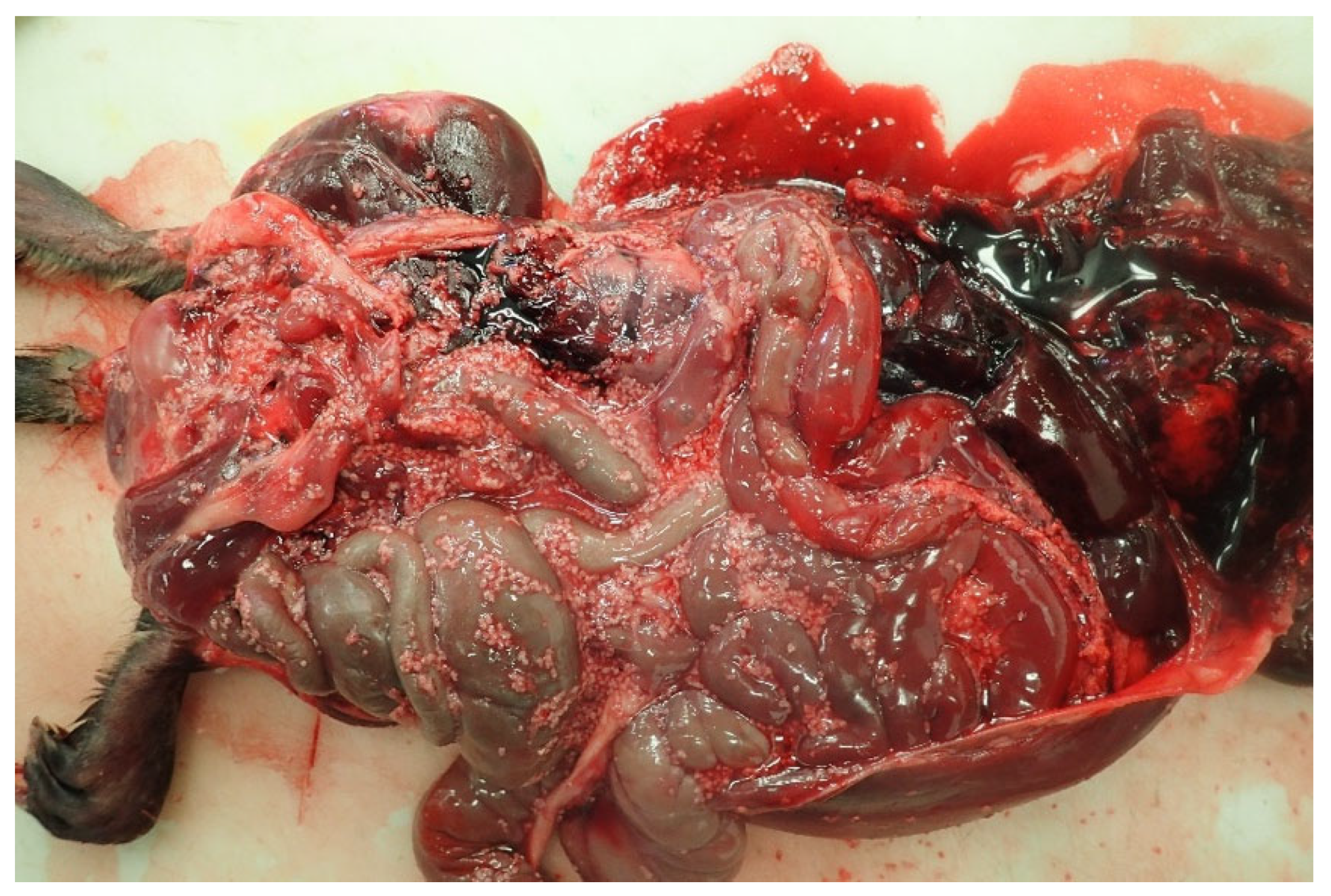
3.1. The Muskrat Case
3.2. The Dog Case
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Freeman, R.S. Studies on the biology of Taenia crassiceps (Zeder, 1800) Rudolph, 1810 (Cestoda). Can. J. Zool. 1962, 40, 969–990. [Google Scholar] [CrossRef]
- Hoberg, E.P.; Ebinger, W.; Render, J.A. Fatal Cysticercosis by Taenia crassiceps (Cyclophyllidea: Taeniidae) in a Presumed Immunocompromised Canine Host. J. Parasitol. 1999, 85, 1174–1178. [Google Scholar] [CrossRef]
- Arocker-Mettinger, E.; Huber-Spitzy, V.; Auer, H.; Grabner, G.; Stur, M. Taenia crassiceps in der Vorderkammer des menschlichen Auges—Ein Fallbericht. Klin. Mon. Augenheilkd. 1992, 201, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Ballweber, L.R. Taenia crassiceps subcutaneous cysticercosis in an adult dog. Vet. Rec. 2009, 165, 693–694. [Google Scholar] [PubMed]
- Bauer, C.; Thiel, W.; Bachmann, R. Taenia crassiceps metacestodes in the subcutis of a dog. Kleintierpraxis 1998, 43, 37–41. [Google Scholar]
- Suja, M.S.; Mahadevan, A.; Madhusudana, S.N.; Vijayasarathy, S.K.; Shankar, S.K. Cerebral cysticercosis mimicking rabies in a dog. Vet. Rec. 2003, 153, 304–305. [Google Scholar] [CrossRef]
- Wünschmann, A.; Garlie, V.; Averbeck, G.; Kurtz, H.; Hoberg, E.P. Cerebral cysticercosis by Taenia crassiceps in a domestic cat. J. Vet. Diagn. Investig. 2003, 15, 484–488. [Google Scholar] [CrossRef]
- Chermette, R.; Bussieras, J.; Marionneau, J.; Boyer, E.; Roubin, C.; Prophette, B.; Maillard, H.; Fabiani, B. Taenia crassiceps invasive cysticercosis in an AIDS patient. Bull. Acad. Nat. Med. Paris 1995, 179, 777–783. [Google Scholar]
- Heldwein, K.; Biedermann, H.; Hamperl, W.; Bretzel, G.; Löscher, T.; Laregina, D.; Frosch, M.; Büttner, D.; Tappe, D. Subcutaneous Taenia crassiceps infection in a patient with non-Hodgkin’s lymphoma. Am. J. Trop. Med. Hyg. 2006, 75, 108–111. [Google Scholar] [CrossRef]
- Maillard, H.; Marionneau, J.; Prophette, B.; Boyer, E.; Celerier, P. Taenia crassiceps cysticercosis and AIDS. AIDS 1998, 12, 1551–1552. [Google Scholar] [CrossRef]
- Ntoukas, V.; Tappe, D.; Pfütze, D.; Simon, M.; Holzmann, T. Cerebellar cysticercosis caused by larval Taenia crassiceps tapeworm in immunocompetent woman, Germany. Emerg. Infect. Dis. 2013, 19, 2008–2011. [Google Scholar] [CrossRef] [PubMed]
- Shea, M.; Maberley, A.L.; Walters, J.; Freeman, R.S.; Fallis, A.M. Intraocular Taenia crassiceps (Cestoda). Trans. Am. Acad. Ophthalmol. Otolaryngol. 1973, 77, 778–783. [Google Scholar]
- Bowles, J.; Blair, D.; McManus, D.P. Genetic variants within the genus Echinococcus identified by mitochondrial DNA sequencing. Mol. Biochem. Parasitol. 1992, 54, 165–173. [Google Scholar] [CrossRef]
- Loos-Frank, B. An up-date of Verster’s (1969) ‘Taxonomic revision of the genus Taenia Linnaeus’ (Cestoda) in table format. Syst. Parasitol. 2000, 45, 155–183. [Google Scholar] [CrossRef] [PubMed]
- Bilqees, F.M.; Freeman, R.S. Histogenesis of the rostellum of Taenia crassiceps (Zeder, 1800) (Cestoda), with special reference to hook development. Can. J. Zool. 1969, 47, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.K.; Gerald, W.E.; Kuhn, R.E. Growth and development of larval Taenia crassiceps (Cestoda)—I.: Aneuploidy in the anomalous Orf strain. Int. J. Parasitol. 1972, 2, 261–262. [Google Scholar] [CrossRef]
- Bowman, D.D. Georgis’ Parasitology for Veterinarians, 11th ed.; Saunders Co.: Philadelphia, PA, USA, 2020; pp. 414–415. [Google Scholar]
- Lescano, A.G.; Zunt, J. Other cestodes: Sparganosis, coenurosis and Taenia crassiceps cysticercosis. Handb. Clin. Neurol. 2013, 114, 335–345. [Google Scholar] [CrossRef]
- Jones, A.; Pybus, M.J. Chapter 7: Taeniasis and Echinococcosis. In Parasitic Diseases of Wild Mammals, 2nd ed.; Samuel, M.W., Pybus, M.J., Kocan, A.A., Eds.; Ohio State Universtiy Press: Columbus, OH, USA, 2001; pp. 150–192. [Google Scholar] [CrossRef]
- Friedland, T.; Steiner, B.; Böckeler, W. Prävalenz der cysticercose bei bisams (Ondatra zibethica L.) in Schleswig-Holstein. Z. Jagdwiss. 1985, 31, 134–139. [Google Scholar] [CrossRef]
- Anderson, W.I.; Scott, D.W.; Hornbuckle, W.E.; King, J.M.; Tennant, B.C. Taenia crassiceps infection in the woodchuck: A retrospective study of 13 cases. Vet. Dermatol. 1990, 1, 85–92. [Google Scholar] [CrossRef]
- Escobedo, G.; Larralde, C.; Chavarria, A.; Cerbón, M.A.; Morales-Montor, J. Molecular mechanisms involved in the differential effects of sex steroids on the reproduction and infectivity of Taenia crassiceps. J. Parasitol. 2004, 90, 1235–1244. [Google Scholar] [CrossRef]
- Larralde, C.; Morales, J.; Terrazas, I.; Govezensky, T.; Romano, M.C. Sex hormone changes induced by the parasite lead to feminization of the male host in murine Taenia crassiceps cysticercosis. J. Steroid Biochem. Mol. Biol. 1995, 52, 575–580. [Google Scholar] [CrossRef]
- Adalid-Peralta, L.; Lopez-Roblero, A.; Camacho-Vázquez, C.; Nájera-Ocampo, M.; Guevara-Salinas, A.; Ruiz-Monroy, N.; Melo-Salas, M.; Morales-Ruiz, V.; López-Recinos, D.; Ortiz-Hernández, E.; et al. Regulatory T-cells as an-escape mechanism to the immune response in Taenia crassiceps infection. Front. Cell. Infect. Microbiol. 2021, 11, 630583. Available online: https://www.frontiersin.org/article/10.3389/fcimb.2021.630583 (accessed on 21 April 2022). [CrossRef]
- Fragoso, G.; Lamoyi, E.; Mellor, A.; Lomelí, C.; Hernández, M.; Sciutto, M. Increased resistance to Taenia crassiceps murine cysticercosis in Qa-2 transgenic mice. Infect. Immun. 1998, 66, 760–764. [Google Scholar] [CrossRef] [PubMed]
- François, A.; Favennec, L.; Cambon-Michot, C.; Gueit, I.; Biga, N.; Tron, F.; Brasseur, P.; Hemet, J. Taenia crassiceps invasive cysticercosis: A new human pathogen in acquired immunodeficiency syndrome? Am. J. Surg. Pathol. 1998, 22, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.; Kursh, L.; Nolan, T.; Perry, J. Subcutaneous Taenia crassiceps cysticercosis mass excision from an 11-year-old mixed-breed dog. J. Am. Anim. Hosp. Assoc. 2021, 57, 232–236. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Abdu, A.; Wu, T.; Forzán, M.J.; Hammer, K.; Lejeune, M. Taenia crassiceps Cysticercosis in a Wild Muskrat and a Domestic Dog in the Northeastern United States. Pathogens 2023, 12, 204. https://doi.org/10.3390/pathogens12020204
Zhang Y, Abdu A, Wu T, Forzán MJ, Hammer K, Lejeune M. Taenia crassiceps Cysticercosis in a Wild Muskrat and a Domestic Dog in the Northeastern United States. Pathogens. 2023; 12(2):204. https://doi.org/10.3390/pathogens12020204
Chicago/Turabian StyleZhang, Yufei, Amira Abdu, Timothy Wu, María J. Forzán, Kimberly Hammer, and Manigandan Lejeune. 2023. "Taenia crassiceps Cysticercosis in a Wild Muskrat and a Domestic Dog in the Northeastern United States" Pathogens 12, no. 2: 204. https://doi.org/10.3390/pathogens12020204
APA StyleZhang, Y., Abdu, A., Wu, T., Forzán, M. J., Hammer, K., & Lejeune, M. (2023). Taenia crassiceps Cysticercosis in a Wild Muskrat and a Domestic Dog in the Northeastern United States. Pathogens, 12(2), 204. https://doi.org/10.3390/pathogens12020204