
LipL41 and LigA/LigB Gene Silencing on a LipL32 Knockout Leptospira interrogans Reveals the Impact of Multiple Mutations on Virulence

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Plasmid Curing on lipL32 Knockout L. interrogans
2.3. Bacterial Conjugation
2.4. Electrophoresis and Immunoblotting
2.5. Animal Ethics Statement
2.6. Animal Infection
2.7. Quantification of Leptospires in Target Organs
3. Results
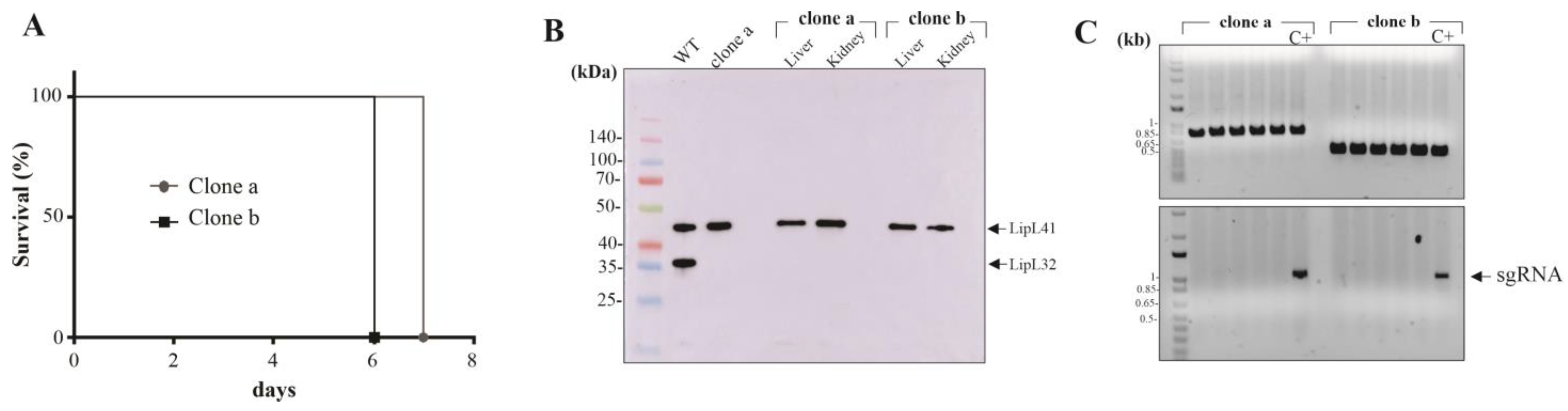
3.1. Plasmid Curing in the LipL32 Mutant L. interrogans
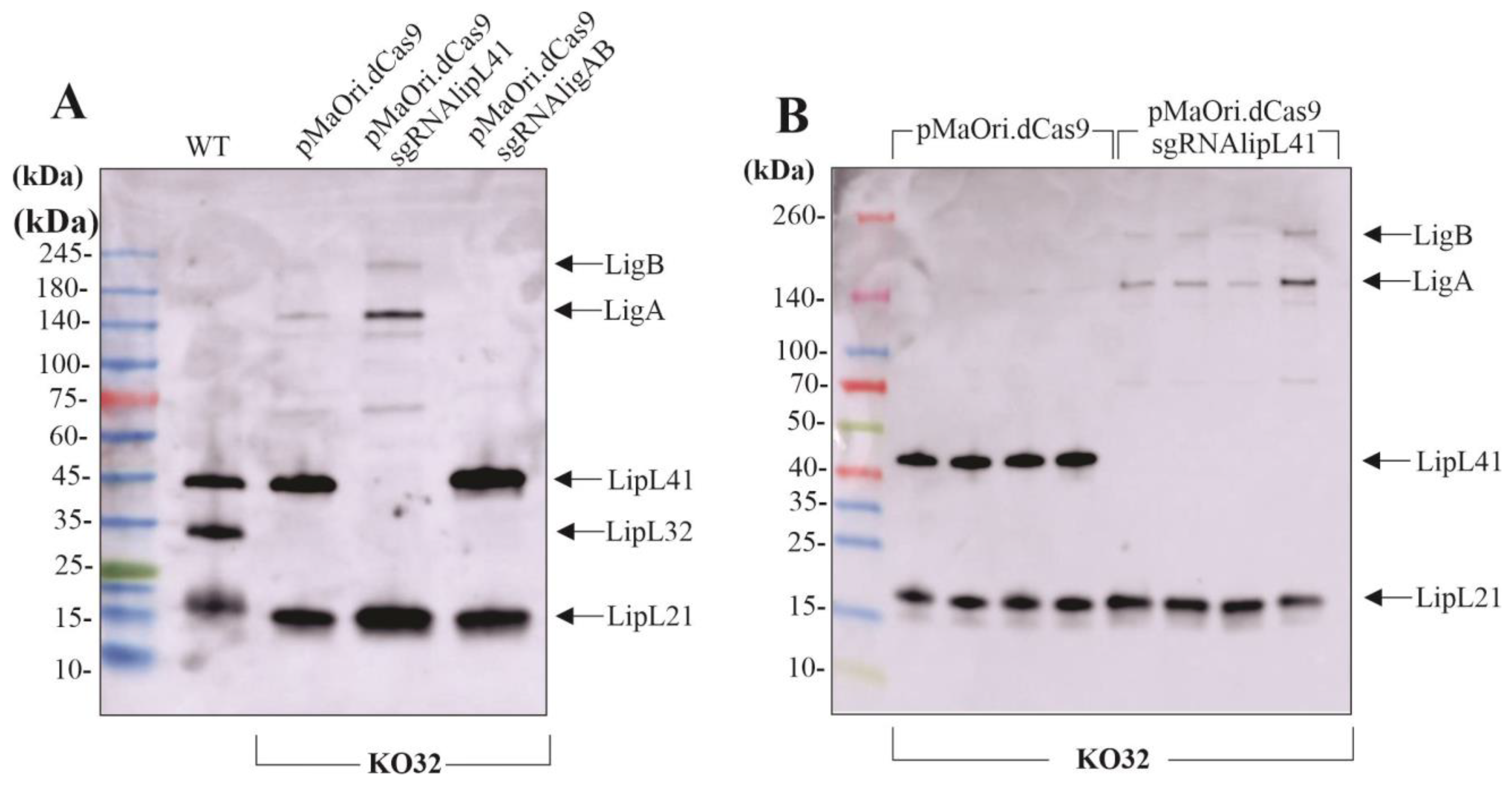
3.2. Gene Silencing by CRISPRi in KO32 L. interrogans
3.3. Effect of Multiple Protein Mutation upon Leptospira Virulence
3.4. Bacterial Burden in Target Organs and Mutant Recovery
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bharti, A.R.; Nally, J.E.; Ricaldi, J.N.; Matthias, M.A.; Diaz, M.M.; Lovett, M.A.; Levett, P.N.; Gilman, R.H.; Willig, M.R.; Gotuzzo, E.; et al. Leptospirosis: A zoonotic disease of global importance. Lancet Infect. Dis. 2003, 3, 757–771. [Google Scholar] [CrossRef]
- Haake, D.A.; Levett, P.N. Leptospirosis in humans. Curr. Top. Microbiol. Immunol. 2015, 387, 65–97. [Google Scholar]
- Torgerson, P.R.; Hagan, J.E.; Costa, F.; Calcagno, J.; Kane, M.; Martinez-Silveira, M.S.; Goris, M.G.A.; Stein, C.; Ko, A.I.; Abela-Ridder, B. Global Burden of Leptospirosis: Estimated in Terms of Disability Adjusted Life Years. PLoS Negl. Trop. Dis. 2015, 9, e0004122. [Google Scholar] [CrossRef]
- Costa, F.; Hagan, J.E.; Calcagno, J.; Kane, M.; Torgerson, P.; Martinez-Silveira, M.S.; Stein, C.; Abela-Ridder, B.; Ko, A.I. Global Morbidity and Mortality of Leptospirosis: A Systematic Review. PLoS Negl. Trop. Dis. 2015, 9, e0003898. [Google Scholar] [CrossRef]
- Ko, A.I.; Reis, M.G.; Dourado, C.M.R.; Johnson, W.D.; Riley, L.W.; Salvador Leptospirosis Study Group. Urban epidemic of severe leptospirosis in Brazil. Lancet 1999, 354, 820–825. [Google Scholar] [CrossRef]
- Reis, R.B.; Ribeiro, G.S.; Felzemburgh, R.D.M.; Santana, F.S.; Mohr, S.; Melendez, A.X.T.O.; Queiroz, A.; Santos, A.C.; Ravines, R.R.; Tassinari, W.S.; et al. Impact of environment and social gradient on Leptospira infection in urban slums. PLoS Negl. Trop. Dis. 2008, 2, e228. [Google Scholar] [CrossRef]
- Levett, P.N. Leptospirosis. Clin. Microbiol. Rev. 2001, 14, 296–326. [Google Scholar] [CrossRef]
- Segura, E.R.; Ganoza, C.A.; Campos, K.; Ricaldi, J.N.; Torres, S.; Silva, H.; Céspedes, M.J.; Matthias, M.A.; Swancutt, M.A.; Liñán, R.L.; et al. Clinical spectrum of pulmonary involvement in leptospirosis in a region of endemicity, with quantification of leptospiral burden. Clin. Infect. Dis. 2005, 40, 343–351. [Google Scholar] [CrossRef]
- McBride, A.J.; Athanazio, D.A.; Reis, M.G.; Ko, A.I. Leptospirosis. Curr. Opin. Infect. Dis. 2005, 18, 376–386. [Google Scholar] [CrossRef]
- Putz, E.J.; Nally, J.E. Investigating the Immunological and Biological Equilibrium of Reservoir Hosts and Pathogenic. Front. Microbiol. 2020, 11, 2005. [Google Scholar] [CrossRef]
- Campos, Â.P.; Miranda, D.F.H.; Rodrigues, H.W.S.; da Silva Carneiro Lustosa, M.; Martins, G.H.C.; Mineiro, A.L.B.B.; Castro, V.; Azevedo, S.S.; de Sousa Silva, S.M.M. Seroprevalence and risk factors for leptospirosis in cattle, sheep, and goats at consorted rearing from the State of Piauí, northeastern Brazil. Trop. Anim. Health Prod. 2017, 49, 899–907. [Google Scholar] [CrossRef]
- Pinto, P.S.; Libonati, H.; Lilenbaum, W. A systematic review of leptospirosis on dogs, pigs, and horses in Latin America. Trop. Anim. Health Prod. 2017, 49, 231–238. [Google Scholar] [CrossRef]
- Guitian, J.; Thurmond, M.C.; Hietala, S.K. Infertility and abortion among first-lactation dairy cows seropositive or seronegative for Leptospira interrogans serovar hardjo. J. Am. Vet. Med. Assoc. 1999, 215, 515–518. [Google Scholar]
- Martins, G.; Lilenbaum, W. Control of bovine leptospirosis: Aspects for consideration in a tropical environment. Res. Vet. Sci. 2017, 112, 156–160. [Google Scholar] [CrossRef]
- Fernandes, L.G.V.; Guaman, L.P.; Vasconcellos, S.A.; Heinemann, M.B.; Picardeau, M.; Nascimento, A.L.T.O. Gene silencing based on RNA-guided catalytically inactive Cas9 (dCas9): A new tool for genetic engineering in Leptospira. Sci. Rep. 2019, 9, 1839. [Google Scholar] [CrossRef]
- Fernandes, L.G.V.; Hornsby, R.L.; Nascimento, A.L.T.O.; Nally, J.E. Genetic manipulation of pathogenic Leptospira: CRISPR interference (CRISPRi)-mediated gene silencing and rapid mutant recovery at 37 °C. Sci. Rep. 2021, 11, 1768. [Google Scholar] [CrossRef]
- Fernandes, L.G.V.; Putz, E.J.; Stasko, J.; Lippolis, J.D.; Nascimento, A.L.T.O.; Nally, J.E. Evaluation of LipL32 and LigA/LigB Knockdown Mutants in Leptospira interrogans Serovar Copenhageni: Impacts to Proteome and Virulence. Front. Microbiol. 2022, 12, 799012. [Google Scholar] [CrossRef]
- Fernandes, L.G.V.; Nascimento, A.L.T.O. A Novel Breakthrough in Leptospira spp. Mutagenesis: Knockout by Combination of CRISPR/Cas9 and Non-homologous End-Joining Systems. Front. Microbiol. 2022, 13, 915382. [Google Scholar] [CrossRef]
- Qi, L.S.; Larson, M.H.; Gilbert, L.A.; Doudna, J.A.; Weissman, J.S.; Arkin, A.P.; Lim, W.A. Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef]
- Fernandes, L.G.V.; Teixeira, A.F.; Nascimento, A.L.T.O. Evaluation of Leptospira interrogans knockdown mutants for LipL32, LipL41, LipL21, and OmpL1 proteins. Front. Microbiol. 2023, 14, 1199660. [Google Scholar] [CrossRef]
- Turner, L.H. Leptospirosis. 3. Maintenance, isolation and demonstration of leptospires. Trans. R. Soc. Trop. Med. Hyg. 1970, 64, 623–646. [Google Scholar] [CrossRef] [PubMed]
- Demarre, G.; Guérout, A.-M.; Matsumoto-Mashimo, C.; Rowe-Magnus, D.A.; Marlière, P.; Mazel, D. A new family of mobilizable suicide plasmids based on broad host range R388 plasmid (IncW) and RP4 plasmid (IncPalpha) conjugative machineries and their cognate Escherichia coli host strains. Res. Microbiol. 2005, 156, 245–255. [Google Scholar] [CrossRef]
- Picardeau, M. Conjugative transfer between Escherichia coli and Leptospira spp. as a new genetic tool. Appl. Environ. Microbiol. 2008, 74, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Pappas, C.J.; Benaroudj, N.; Picardeau, M. A replicative plasmid vector allows efficient complementation of pathogenic Leptospira strains. Appl. Environ. Microbiol. 2015, 81, 3176–3181. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, A.L.; Ko, A.I.; Martins, E.A.L.; Monteiro-Vitorello, C.B.; Ho, P.L.; Haake, D.A.; Verjovski-Almeida, S. Comparative genomics of two Leptospira interrogans serovars reveals novel insights into physiology and pathogenesis. J. Bacteriol. 2004, 186, 2164–2172. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.-X.; Fu, G.; Jiang, X.-G.; Zeng, R.; Miao, Y.-G.; Xu, H.; Zhang, Y.-X.; Xiong, H.; Lu, G.; Lu, L.-F.; et al. Unique physiological and pathogenic features of Leptospira interrogans revealed by whole-genome sequencing. Nature 2003, 422, 888–893. [Google Scholar] [CrossRef] [PubMed]
- Bulach, D.M.; Zuerner, R.L.; Wilson, P.; Seemann, T.; McGrath, A.; Cullen, P.A.; Davis, J.; Johnson, M.; Kuczek, E.; Alt, D.P.; et al. Genome reduction in Leptospira borgpetersenii reflects limited transmission potential. Proc. Natl. Acad. Sci. USA 2006, 103, 14560–14565. [Google Scholar] [CrossRef]
- Ko, A.I.; Goarant, C.; Picardeau, M. Leptospira: The dawn of the molecular genetics era for an emerging zoonotic pathogen. Nat. Rev. Microbiol. 2009, 7, 736–747. [Google Scholar] [CrossRef]
- Daroz, B.B.; Fernandes, L.G.V.; Cavenague, M.F.; Kochi, L.T.; Passalia, F.J.; Takahashi, M.B.; Filho, E.G.N.; Teixeira, A.F.; Nascimento, A.L.T.O. A Review on Host- Leptospira Interactions: What We Know and Future Expectations. Front. Cell Infect. Microbiol. 2021, 11, 777709. [Google Scholar] [CrossRef]
- Pereira, P.R.M.; Fernandes, L.G.V.; de Souza, G.O.; Vasconcellos, S.A.; Heinemann, M.B.; Romero, E.C.; Nascimento, A.L.T.O. Multifunctional and Redundant Roles of Leptospira interrogans Proteins in Bacterial-Adhesion and fibrin clotting inhibition. Int. J. Med. Microbiol. 2017, 307, 297–310. [Google Scholar] [CrossRef]
- Haake, D.A.; Chao, G.; Zuerner, R.L.; Barnett, J.K.; Barnett, D.; Mazel, M.; Matsunaga, J.; Levett, P.N.; Bolin, C.A. The leptospiral major outer membrane protein LipL32 is a lipoprotein expressed during mammalian infection. Infect. Immun. 2000, 68, 2276–2285. [Google Scholar] [CrossRef] [PubMed]
- Hauk, P.; Barbosa, A.S.; Ho, P.L.; Farah, C.S. Calcium binding to leptospira outer membrane antigen LipL32 is not necessary for its interaction with plasma fibronectin, collagen type IV, and plasminogen. J. Biol. Chem. 2012, 287, 4826–4834. [Google Scholar] [CrossRef] [PubMed]
- Murray, G.L. The lipoprotein LipL32, an enigma of leptospiral biology. Vet. Microbiol. 2013, 162, 305–314. [Google Scholar] [CrossRef]
- Malmström, J.; Beck, M.; Schmidt, A.; Lange, V.; Deutsch, E.W.; Aebersold, R. Proteome-wide cellular protein concentrations of the human pathogen Leptospira interrogans. Nature 2009, 460, 762–765. [Google Scholar] [CrossRef] [PubMed]
- Cullen, P.A.; Cordwell, S.J.; Bulach, D.M.; Haake, D.A.; Adler, B. Global analysis of outer membrane proteins from Leptospira interrogans serovar Lai. Infect. Immun. 2002, 70, 2311–2318. [Google Scholar] [CrossRef]
- Murray, G.L.; Srikram, A.; Hoke, D.E.; Wunder, E.A., Jr.; Henry, R.; Lo, M.; Zhang, K.; Sermswan, R.W.; Ko, A.I.; Adler, B. Major surface protein LipL32 is not required for either acute or chronic infection with Leptospira interrogans. Infect. Immun. 2009, 77, 952–958. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.; Bartpho, T.; Sermswan, R.W.; Bulach, D.M.; Eshghi, A.; Picardeau, M.; Adler, B.; Murray, G.L. Leptospiral outer membrane protein LipL41 is not essential for acute leptospirosis but requires a small chaperone protein, lep, for stable expression. Infect. Immun. 2013, 81, 2768–2776. [Google Scholar] [CrossRef]
- Pappas, C.J.; Picardeau, M. Control of Gene Expression in Leptospira spp. by Transcription Activator-Like Effectors Demonstrates a Potential Role for LigA and LigB in Leptospira interrogans Virulence. Appl. Environ. Microbiol. 2015, 81, 7888–7892. [Google Scholar] [CrossRef]
- Haake, D.A.; Matsunaga, J. Leptospiral Immunoglobulin-Like Domain Proteins: Roles in Virulence and Immunity. Front. Immunol. 2020, 11, 579907. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LipL32 Knockout Strain Containing CRISPRi Plasmids | |||||
|---|---|---|---|---|---|
| Endpoints/Total | Endpoint Day | Average Weight (%) | |||
| 4 | 5 | 6 | |||
| pMaOri.dCas9 | 5/5 | 6, 6, 6, 6, 7 | 108.7 | 101.2 | 98.8 |
| pMaOri.dCas9sgRNAlipL41 | 5/5 | 6, 6, 6, 7, 7 | 105.6 | 98.99 | 95.2 |
| pMaOri.dCas9sgRNAligAB | 0/4 | - | 107.8 | 104.8 | 105.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, L.G.V.; Foltran, B.B.; Teixeira, A.F.; Nascimento, A.L.T.O. LipL41 and LigA/LigB Gene Silencing on a LipL32 Knockout Leptospira interrogans Reveals the Impact of Multiple Mutations on Virulence. Pathogens 2023, 12, 1191. https://doi.org/10.3390/pathogens12101191
Fernandes LGV, Foltran BB, Teixeira AF, Nascimento ALTO. LipL41 and LigA/LigB Gene Silencing on a LipL32 Knockout Leptospira interrogans Reveals the Impact of Multiple Mutations on Virulence. Pathogens. 2023; 12(10):1191. https://doi.org/10.3390/pathogens12101191
Chicago/Turabian StyleFernandes, Luis Guilherme V., Bruno B. Foltran, Aline F. Teixeira, and Ana Lucia Tabet Oller Nascimento. 2023. "LipL41 and LigA/LigB Gene Silencing on a LipL32 Knockout Leptospira interrogans Reveals the Impact of Multiple Mutations on Virulence" Pathogens 12, no. 10: 1191. https://doi.org/10.3390/pathogens12101191
APA StyleFernandes, L. G. V., Foltran, B. B., Teixeira, A. F., & Nascimento, A. L. T. O. (2023). LipL41 and LigA/LigB Gene Silencing on a LipL32 Knockout Leptospira interrogans Reveals the Impact of Multiple Mutations on Virulence. Pathogens, 12(10), 1191. https://doi.org/10.3390/pathogens12101191