
Epidemiology and Transmission of Theileria orientalis in Australasia


Abstract
:1. Introduction
2. Classification of T. orientalis
3. Epidemiology of Theileriosis Caused by T. orientalis
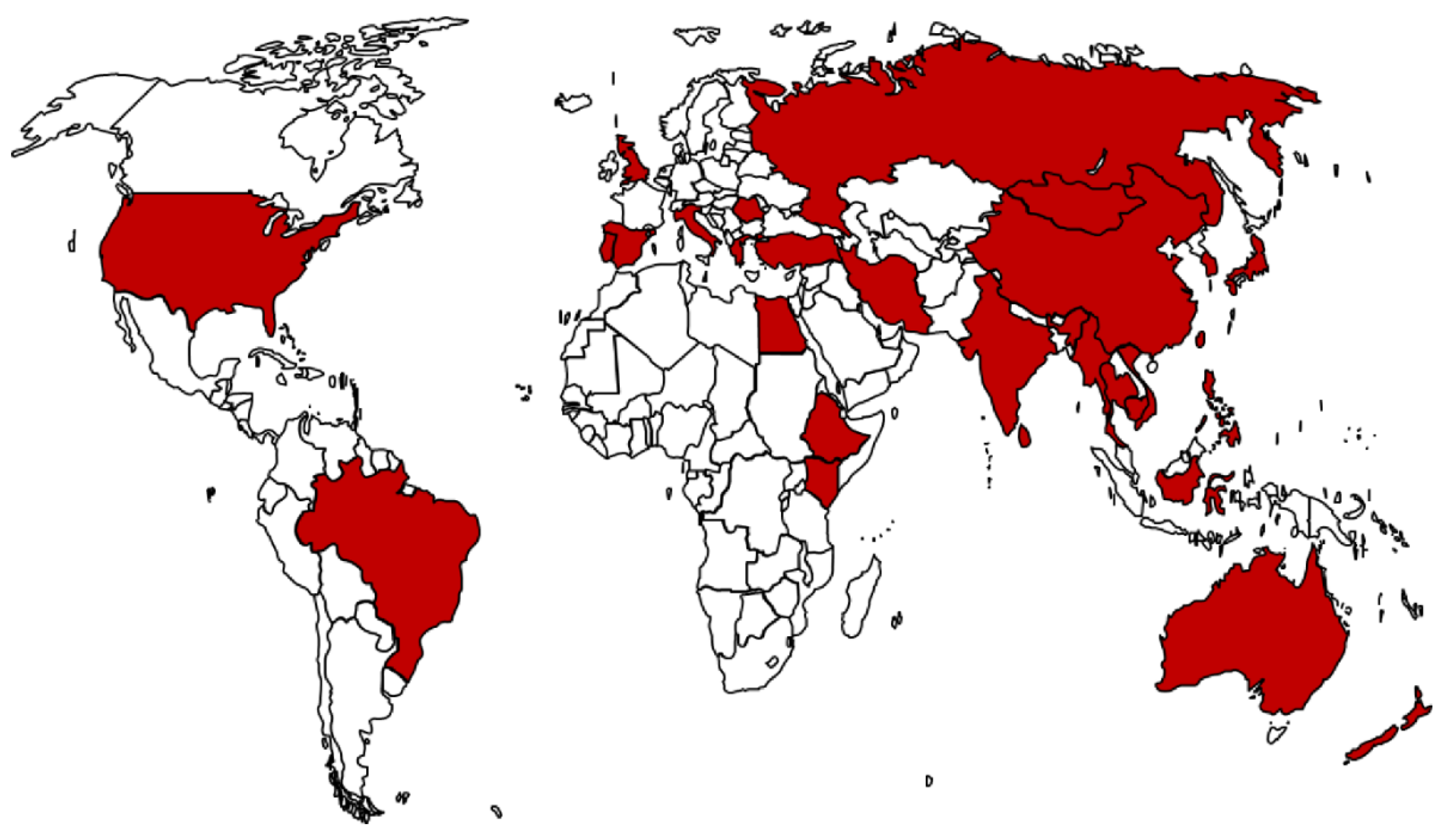
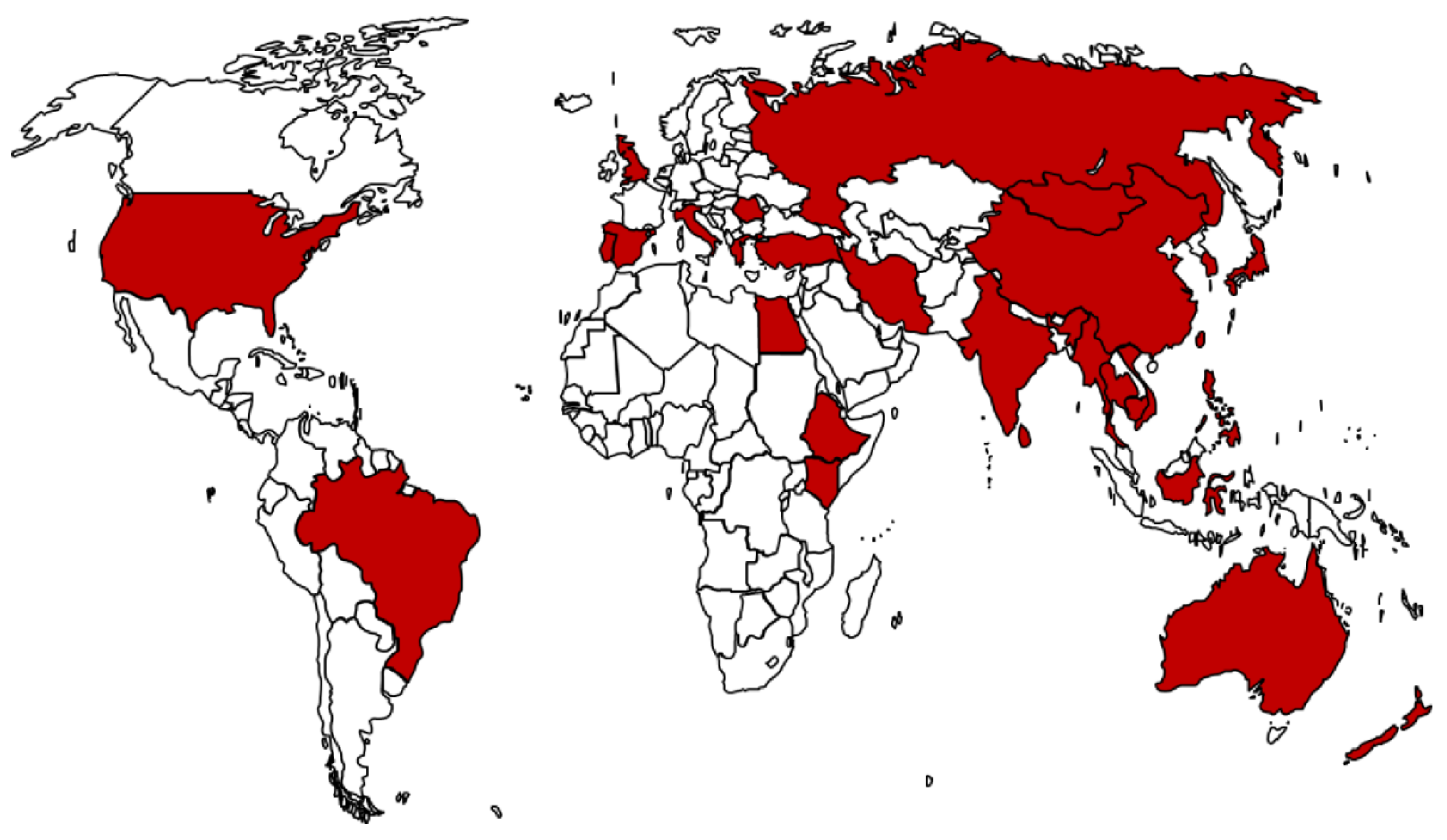
3.1. Worldwide Distribution
3.2. Theileriosis in Australasia
3.2.1. Introduction and Historical Aspects
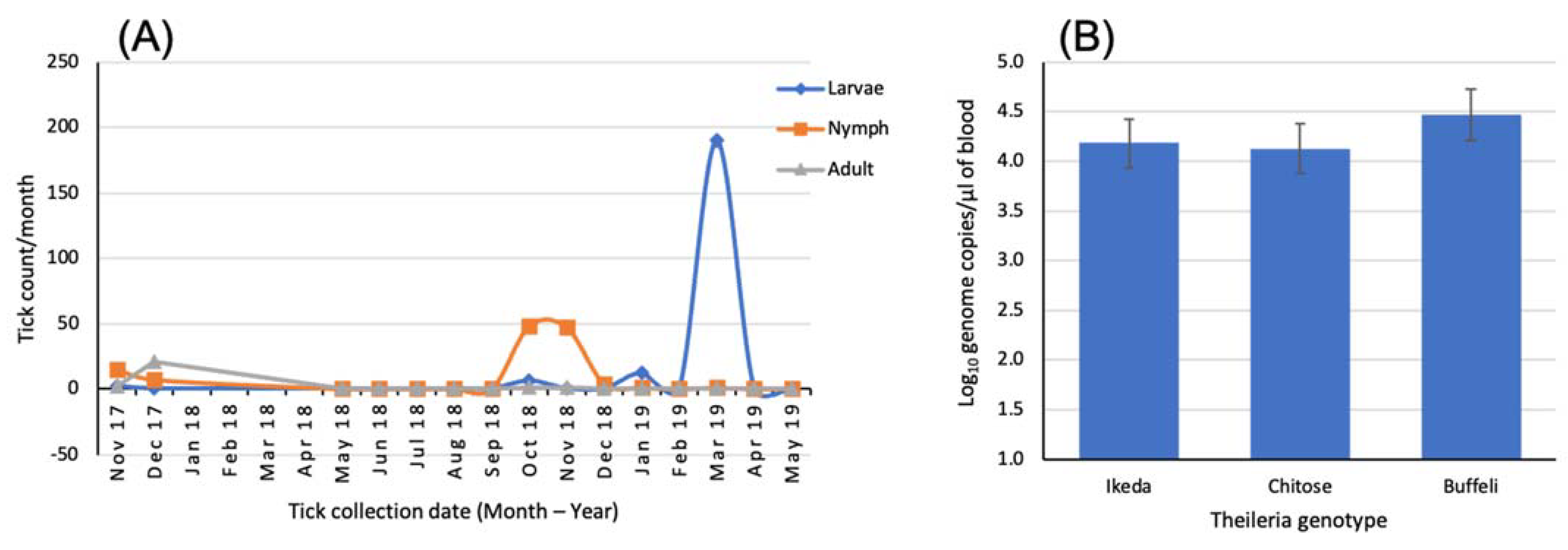
3.2.2. Pathogen Factors
3.2.3. Host Factors
3.2.4. Environmental Factors
4. Lifecycle, Vectors, and Modes of Transmission
4.1. Biological Vectors
4.2. Mechanical Vectors
4.3. Other Means of Transmission
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sivakumar, T.; Hayashida, K.; Sugimoto, C.; Yokoyama, N. Evolution and genetic diversity of Theileria. Infect. Genet. Evol. 2014, 27, 250–263. [Google Scholar] [CrossRef] [PubMed]
- Yam, J.; Bogema, D.; Jenkins, C. Oriental Theileriosis. In Ticks and Tick-Borne Pathogens; IntechOpen: London, UK, 2018. [Google Scholar]
- Gebrekidan, H.; Perera, P.K.; Ghafar, A.; Abbas, T.; Gasser, R.B.; Jabbar, A. An appraisal of oriental theileriosis and the Theileria orientalis complex, with an emphasis on diagnosis and genetic characterisation. Parasitol. Res. 2019, 119, 11–22. [Google Scholar] [CrossRef]
- Emery, D.L. Approaches to Integrated Parasite Management (IPM) for Theileria orientalis with an Emphasis on Immunity. Pathogens 2021, 10, 1153. [Google Scholar] [CrossRef]
- Taylor, M.A.; Coop, R.L.; Wall, R.L. Veterinary Parasitology, 4th ed.; Wiley Blackwell: New Delhi, India, 2016. [Google Scholar]
- Jabbar, A.; Abbas, T.; Sandhu, Z.-U.; A Saddiqi, H.; Qamar, M.F.; Gasser, R.B. Tick-borne diseases of bovines in Pakistan: Major scope for future research and improved control. Parasites Vectors 2015, 8, 1–13. [Google Scholar] [CrossRef]
- Robertson, W. African coast fever. J. Comp. Pathol. Ther. 1904, 17, 214-IN6. [Google Scholar] [CrossRef]
- Deken, R.D.; Martin, V.; Saido, A.; Madder, M.; Brandt, J.; Geysen, D. An outbreak of East Coast Fever on the Comoros: A consequence of the import of immunised cattle from Tanzania? Vet. Parasitol. 2007, 143, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Gul, N.; Ayaz, S.; Gul, I.; Adnan, M.; Shams, S.; Akbar, N. Tropical Theileriosis and East Coast Fever in Cattle: Present, Past and Future Perspective. Int. J. Curr. Microbiol. App. Sci. 2015, 4, 1000–1018. [Google Scholar]
- Dodd, S. Piroplasmosis of cattle in Queensland. J. Comp. Pathol. Ther. 1910, 23, 141–160. [Google Scholar] [CrossRef]
- Seddon, H. Diseases of Domestic Animals in Australia: Pt. 4; Department of Health. Service Publication (Veterinary Hygiene): Canberra, Australia, 1952. [Google Scholar]
- Stewart, N.; Standfast, N.; Baldock, F.; Reid, D.; Vos, A. The distribution and prevalence of Theileria buffeli in cattle in Queensland. Aust. Vet. J. 1992, 69, 59–61. [Google Scholar] [CrossRef] [PubMed]
- Uilenberg, G. Babesia—A historical overview. Vet. Parasitol. 2006, 138, 3–10. [Google Scholar] [CrossRef]
- Khukhuu, A.; Lan, D.T.B.; Long, P.T.; Ueno, A.; Li, Y.; Luo, Y.; de Macedo, A.C.C.; Matsumoto, K.; Inokuma, H.; Kawazu, S.-I.; et al. Molecular Epidemiological Survey of Theileria orientalis in Thua Thien Hue Province, Vietnam. J. Vet. Med. Sci. 2011, 73, 701–705. [Google Scholar] [CrossRef]
- Izzo, M.; Poe, I.; Horadagoda, N.; De Vos, A.; House, J. Haemolytic anaemia in cattle in NSW associated with Theileria infections. Aust. Vet. J. 2010, 88, 45–51. [Google Scholar] [CrossRef] [PubMed]
- McFadden, A.; Rawdon, T.G.; Meyer, J.; Makin, J.; Morley, C.M.; Clough, R.R.; Tham, K.; Müllner, P.; Geysen, D. An outbreak of haemolytic anaemia associated with infection of Theileria orientalis in naive cattle. N. Z. Vet. J. 2011, 59, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Bogema, D.R.; Micallef, M.L.; Liu, M.; Padula, M.P.; Djordjevic, S.P.; Darling, A.E.; Jenkins, C. Analysis of Theileria orientalis draft genome sequences reveals potential species-level divergence of the Ikeda, Chitose and Buffeli genotypes. BMC Genom. 2018, 19, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yam, J.; Bogema, D.R.; Micallef, M.L.; Djordjevic, S.P.; Jenkins, C. Complete Genomes of Theileria orientalis Chitose and Buffeli Genotypes Reveal within Species Translocations and Differences in ABC Transporter Content. Pathogens 2022, 11, 801. [Google Scholar] [CrossRef]
- Rogers, R.J.; Callow, L.L. Three fatal cases of theileria mutans infection. Aust. Vet. J. 1966, 42, 42–46. [Google Scholar] [CrossRef]
- Callow, L.L. Animal Health in Australia. Volume 5. Protozoal and Rickettsial Diseases; Australian Government Publishing Service: Canberra, Australia, 1984; pp. 169–173. [Google Scholar]
- Stewart, N.P.; de Vos, A.J.; McGregor, W.; Shiels, I. Haemaphysalis humerosa not H. longicornis is the likely vector of Theileria buffeli in Australia. Aust. Vet. J. 1987, 64, 280–282. [Google Scholar] [CrossRef]
- Kubota, S.; Sugimoto, C.; Kakuda, T.; Onuma, M. Analysis of immunodominant piroplasm surface antigen alleles in mixed populations of Theileria sergenti and T. buffeli. Int. J. Parasitol. 1996, 26, 741–747. [Google Scholar] [CrossRef]
- Islam, M.K.; Jabbar, A.; Campbell, B.E.; Cantacessi, C.; Gasser, R.B. Bovine theileriosis–An emerging problem in south-eastern Australia? Infect. Genet. Evol. 2011, 11, 2095–2097. [Google Scholar] [CrossRef]
- Gebrekidan, H.; Gasser, R.B.; Perera, P.K.; McGrath, S.; McGrath, S.; Stevenson, M.A.; Jabbar, A. Investigating the first outbreak of oriental theileriosis in cattle in South Australia using multiplexed tandem PCR (MT-PCR). Ticks Tick-Borne Dis. 2015, 6, 574–578. [Google Scholar] [CrossRef]
- Forshaw, D.; Alex, S.M.; Palmer, D.G.; Cotter, J.; Roberts, W.D.; Jenkins, C.; Hair, S. Theileria orientalis Ikeda genotype infection associated with anaemia, abortion and death in beef cattle in Western Australia. Aust. Vet. J. 2020, 98, 290–297. [Google Scholar] [CrossRef]
- Leong, C.-C.; Oskam, C.L.; Barbosa, A.D.; Aleri, J.W. Distribution and Prevalence of Theileria orientalis Genotypes in Adult Lactating Dairy Cows in South West Region of Western Australia. Pathogens 2023, 12, 125. [Google Scholar] [CrossRef]
- Lakew, B.T.; Kheravii, S.K.; Wu, S.-B.; Eastwood, S.; Andrew, N.R.; Jenkins, C.; Walkden-Brown, S.W. Endemic infection of cattle with multiple genotypes of Theileria orientalis on the Northern Tablelands of New South Wales despite limited presence of ticks. Ticks Tick-Borne Dis. 2020, 12, 101645. [Google Scholar] [CrossRef]
- Jenkins, C. Bovine theileriosis in Australia: A decade of disease. Microbiol. Aust. 2018, 39, 215. [Google Scholar] [CrossRef]
- Stewart, N.P.; Vos, A.J.; Shiels, I.; McGregor, W. The experimental transmission of Theileria buffeli of cattle in Australia by Haemaphysalis humerosa. Aust. Vet. J. 1987, 64, 81–83. [Google Scholar] [CrossRef] [PubMed]
- James, M.; Saunders, B.W.; Guy, L.A.; Brookbanks, E.O.; Charleston, W.A.G.; Uilenberg, G. Theileria orientalis, a blood parasite of cattle. First report in New Zealand. N. Z. Vet. J. 1984, 32, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, K.; McFadden, A.; Gias, E.; Pulford, D.; Pomroy, W. Epidemiology of the epidemic of bovine anaemia associated with Theileria orientalis (Ikeda) between August 2012 and March 2014. N. Z. Vet. J. 2015, 64, 38–47. [Google Scholar] [CrossRef] [PubMed]
- McFadden, A.; Gias, E.; Heuer, C.; McFadden, F.S.; Pulford, D. Prevalence and spatial distribution of cattle herds infected with Theileria orientalis in New Zealand between 2012 and 2013. N. Z. Vet. J. 2016, 64, 55–59. [Google Scholar] [CrossRef]
- Heath, A. Biology, ecology and distribution of the tick, Haemaphysalis longicornis Neumann (Acari: Ixodidae) in New Zealand. N. Z. Vet. J. 2016, 64, 10–20. [Google Scholar] [CrossRef]
- Lawrence, K.; Gedye, K.; Hickson, R.; Wang, B.; Carvalho, L.; Zhao, Y.; Pomroy, W. The role of sheep (Ovis aries) in maintaining Theileria orientalis Ikeda type infection. Vet. Parasitol. 2021, 291, 109391. [Google Scholar] [CrossRef]
- Perera, P.K.; Gasser, R.B.; Read, E.; Malmo, J.; Nguyen, H.; Nyein, S.; Cheng, A.; Jex, A.R.; Rawlin, G.; Spithill, T.W.; et al. Use of multiplexed tandem PCR to estimate the prevalence and intensity of Theileria orientalis infections in cattle. Infect. Genet. Evol. 2015, 32, 68–73. [Google Scholar] [CrossRef]
- Perera, P.K.; Gasser, R.B.; Anderson, G.A.; Jeffers, M.; Bell, C.M.; Jabbar, A. Epidemiological survey following oriental theileriosis outbreaks in Victoria, Australia, on selected cattle farms. Vet. Parasitol. 2013, 197, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Kamau, J.; de Vos, A.J.; Playford, M.; Salim, B.; Kinyanjui, P.; Sugimoto, C. Emergence of new types of Theileria orientalis in Australian cattle and possible cause of theileriosis outbreaks. Parasites Vectors 2011, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.; Micallef, M.; Alex, S.; Collins, D.; Djordjevic, S.; Bogema, D. Temporal dynamics and subpopulation analysis of Theileria orientalis genotypes in cattle. Infect. Genet. Evol. 2015, 32, 199–207. [Google Scholar] [CrossRef]
- Eamens, G.J.; Bailey, G.; Jenkins, C.; Gonsalves, J.R. Significance of Theileria orientalis types in individual affected beef herds in New South Wales based on clinical, smear and PCR findings. Vet. Parasitol. 2013, 196, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Kamau, J.; Salim, B.; Yokoyama, N.; Kinyanjui, P.; Sugimoto, C. Rapid discrimination and quantification of Theileria orientalis types using ribosomal DNA internal transcribed spacers. Infect. Genet. Evol. 2011, 11, 407–414. [Google Scholar] [CrossRef]
- Kubota, S.; Sugimoto, C.; Onuma, M. Population dynamics of Theileria sergenti in persistently infected cattle and vector ticks analysed by a polymerase chain reaction. Parasitology 1996, 112, 437–442. [Google Scholar] [CrossRef]
- Proctor, A.; Ball, M.; Freeman, P.; Jenkins, C.; Bogema, D.R. Prevalence of Theileria orientalis types in beef cattle herds on the North Coast of New South Wales. Aust. Vet. J. 2016, 94, 117–120. [Google Scholar] [CrossRef]
- Bogema, D.R.; Deutscher, A.T.; Fell, S.; Collins, D.; Eamens, G.J.; Jenkins, C. Development and Validation of a Quantitative PCR Assay Using Multiplexed Hydrolysis Probes for Detection and Quantification of Theileria orientalis Isolates and Differentiation of Clinically Relevant Subtypes. J. Clin. Microbiol. 2015, 53, 941–950. [Google Scholar] [CrossRef]
- Gachohi, J.; Skilton, R.; Hansen, F.; Ngumi, P.; Kitala, P. Epidemiology of East Coast fever (Theileria parva infection) in Kenya: Past, present and the future. Parasites Vectors 2012, 5, 194. [Google Scholar] [CrossRef]
- Swilks, E.; Jenkins, C.; Poynting, A.; Collins, D.; Krebs, G. Prevalence and effect of Theileria orientalis infection in homebred calves in the Gloucester region of New South Wales, Australia. Aust. Vet. J. 2017, 95, 211–216. [Google Scholar] [CrossRef]
- Storey-Lewis, B.; Mitrovic, A.; McParland, B. Molecular detection and characterisation of Babesia and Theileria in Australian hard ticks. Ticks Tick-Borne Dis. 2018, 9, 471–478. [Google Scholar] [CrossRef]
- Gebrekidan, H.; Nelson, L.; Smith, G.; Gasser, R.B.; Jabbar, A. An outbreak of oriental theileriosis in dairy cattle imported to Vietnam from Australia. Parasitology 2016, 144, 738–746. [Google Scholar] [CrossRef]
- Watts, J.; Playford, M.; Hickey, K. Theileria orientalis: A review. N. Z. Vet. J. 2016, 64, 3–9. [Google Scholar] [CrossRef]
- Thrusfield, M. Veterinary Epidemiology; John Wiley & Sons: Hoboken, NJ, USA, 2018. [Google Scholar]
- Mahoney, D.F.; Ross, D.R. Epizootiological factors in the control of bovine babesiosis. Aust. Vet. J. 1972, 48, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, D.; Wright, I.G.; Goodger, B.V.; Mirre, G.B.; Sutherst, R.W.; Utech, K.B.W. The transmission ofbabesia bovisin herds of european and zebu x european cattle infested with the tick, boophilus microplus. Aust. Vet. J. 1981, 57, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Swilks, E.; Fell, S.A.; Hammer, J.F.; Sales, N.; Krebs, G.L.; Jenkins, C. Transplacental transmission of Theileria orientalis occurs at a low rate in field-affected cattle: Infection in utero does not appear to be a major cause of abortion. Parasites Vectors 2017, 10, 1–9. [Google Scholar] [CrossRef]
- Randolph, S.E. Chapter 6 Epidemiological Consequences of the Ecological Physiology of Ticks. Adv. Insect Physiol. 2009, 37, 297–339. [Google Scholar] [CrossRef]
- Choi, K.-S.; Yu, D.-H.; Chae, J.-S.; Park, B.-K.; Yoo, J.-G.; Park, J. Seasonal changes in hemograms and Theileria orientalis infection rates among Holstein cattle pastured in the mountains in the Republic of Korea. Prev. Vet. Med. 2016, 127, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Fujisaki, K.; Ito, Y.; Kamio, T.; Kitaoka, S. The presence of Theileria sergenti in Haemaphysalis longicornis overwintering in pasture in Japan. Ann. Trop. Med. Parasitol. 1985, 79, 519–524. [Google Scholar] [CrossRef]
- McFadden, A.; Heuer, C.; Jackson, R.; West, D.; Parkinson, T. Reproductive performance of beef cow herds in New Zealand. N. Z. Vet. J. 2005, 53, 39–44. [Google Scholar] [CrossRef] [PubMed]
- VAN Hennekeler, K.; Jones, R.E.; Skerratt, L.F.; Muzari, M.O.; Fitzpatrick, L.A. Meteorological effects on the daily activity patterns of tabanid biting flies in northern Queensland, Australia. Med. Vet. Entomol. 2011, 25, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Baldacchino, F.; Desquesnes, M.; Mihok, S.; Foil, L.D.; Duvallet, G.; Jittapalapong, S. Tabanids: Neglected subjects of research, but important vectors of disease agents! Infect. Genet. Evol. 2014, 28, 596–615. [Google Scholar] [CrossRef] [PubMed]
- Lakew, B.T.; Kheravii, S.K.; Wu, S.-B.; Eastwood, S.; Andrew, N.R.; Nicholas, A.H.; Walkden-Brown, S.W. Detection and distribution of haematophagous flies and lice on cattle farms and potential role in the transmission of Theileria orientalis. Vet. Parasitol. 2021, 298, 109516. [Google Scholar] [CrossRef] [PubMed]
- Spratt, D. Comparative epidemiology of Dirofilaria roemeri infection in two regions of Queensland. Int. J. Parasitol. 1974, 4, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Okiwelu, S.; Samdi, S. Species diversity, abundance and seasonal occurrence of some biting flies in Southern Kaduna, Nigeria. Afr. J. Biomed. Res. 2006, 8, 113–118. [Google Scholar] [CrossRef]
- Emery, D.; Burgh, S.D.; Dinh, T.H.H.H.; Rolls, P.; Carter, P. Merozoites of Theileria Orientalis Buffeli Reduce Parasitosis Following Challenge by Ticks Infested with T. Orientalis Ikeda. Res. Sq. 2020, 14. [Google Scholar]
- Shaw, M.K. Theileria Development and Host Cell Invasion; Springer: Boston, MA, USA, 2002; pp. 1–22. [Google Scholar] [CrossRef]
- Nuttall, G.H.F.; Warburton, C.; Cooper, W.F.; Robinson, L.E. Ticks: A Monograph of the Ixodidea Part III. The Genus Haemaphysalis; Cambridge University Press: Cambridge, UK, 1915. [Google Scholar]
- Stewart, N.; Devos, A.; Shiels, I.; Jorgensen, W. Transmission of Theileria buffeli to cattle by Haemaphysalis bancrofti fed on Artificially Infected Mice. Vet. Parasitol. 1989, 34, 123–127. [Google Scholar] [CrossRef]
- Riek, R.F. Epidemiology and transmission of Theileria sp of cattle in Australia. Aust. Vet. J. 1982, 59, 89–92. [Google Scholar] [CrossRef]
- Marendy, D.; Baker, K.; Emery, D.; Rolls, P.; Stutchbury, R. Haemaphysalis longicornis: The life-cycle on dogs and cattle, with confirmation of its vector status for Theileria orientalis in Australia. Vet. Parasitol. 2019, 277, 100022. [Google Scholar] [CrossRef]
- Dinkel, K.D.; Herndon, D.R.; Noh, S.M.; Lahmers, K.K.; Todd, S.M.; Ueti, M.W.; Scoles, G.A.; Mason, K.L.; Fry, L.M. A U.S. isolate of Theileria orientalis, Ikeda genotype, is transmitted to cattle by the invasive Asian longhorned tick, Haemaphysalis longicornis. Parasites Vectors 2021, 14, 1–11. [Google Scholar] [CrossRef]
- Thompson, A.T.; White, S.; Shaw, D.; Egizi, A.; Lahmers, K.; Ruder, M.G.; Yabsley, M.J. Theileria orientalis Ikeda in host-seeking Haemaphysalis longicornis in Virginia, U.S.A. Ticks Tick-Borne Dis. 2020, 11, 101450. [Google Scholar] [CrossRef]
- Hammer, J.F.; Emery, D.; Bogema, D.R.; Jenkins, C. Detection of Theileria orientalis genotypes in Haemaphysalis longicornis ticks from southern Australia. Parasites Vectors 2015, 8, 1–8. [Google Scholar] [CrossRef]
- Yokoyama, N.; Sivakumar, T.; Ota, N.; Igarashi, I.; Nakamura, Y.; Yamashina, H.; Matsui, S.; Fukumoto, N.; Hata, H.; Kondo, S.; et al. Genetic diversity of Theileria orientalis in tick vectors detected in Hokkaido and Okinawa, Japan. Infect. Genet. Evol. 2012, 12, 1669–1675. [Google Scholar] [CrossRef] [PubMed]
- Altangerel, K.; Battsetseg, B.; Battur, B.; Sivakumar, T.; Batmagnai, E.; Javkhlan, G.; Tuvshintulga, B.; Igarashi, I.; Matsumoto, K.; Inokuma, H.; et al. The first survey of Theileria orientalis infection in Mongolian cattle. Vet. Parasitol. 2011, 182, 343–348. [Google Scholar] [CrossRef]
- Kumsa, B.; Signorini, M.; Teshale, S.; Tessarin, C.; Duguma, R.; Ayana, D.; Martini, M.; Cassini, R. Molecular detection of piroplasms in ixodid ticks infesting cattle and sheep in western Oromia, Ethiopia. Trop. Anim. Health Prod. 2013, 46, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Abichu, G.; Meli, M.L.; Tánczos, B.; Sulyok, K.M.; Gyuranecz, M.; Gönczi, E.; Farkas, R.; Hofmann-Lehmann, R. Influence of the Biotope on the Tick Infestation of Cattle and on the Tick-Borne Pathogen Repertoire of Cattle Ticks in Ethiopia. PLoS ONE 2014, 9, e106452. [Google Scholar] [CrossRef] [PubMed]
- Aktas, M.; Altay, K.; Ozubek, S.; Dumanli, N. A survey of ixodid ticks feeding on cattle and prevalence of tick-borne pathogens in the Black Sea region of Turkey. Vet. Parasitol. 2012, 187, 567–571. [Google Scholar] [CrossRef]
- Li, L.-H.; Wang, J.-Z.; Zhu, D.; Li, X.-S.; Lu, Y.; Yin, S.-Q.; Li, S.-G.; Zhang, Y.; Zhou, X.-N. Detection of novel piroplasmid species and Babesia microti and Theileria orientalis genotypes in hard ticks from Tengchong County, Southwest China. Parasitol. Res. 2020, 119, 1259–1269. [Google Scholar] [CrossRef]
- Hoogstraal, H.; Roberts, F.H.S.; Kohls, G.M.; Tipton, V.J. Review of Haemaphysalis (Kaiseriana) longicornis Neumann (Resurrected) of Australia, New Zealand, New Caledonia, Fiji, Japan, Korea, and Northeastern China and USSR, and Its Parthenogenetic and Bisexual Populations (Ixodoidea, Ixodidae). J. Parasitol. 1968, 54, 1197–1213. [Google Scholar] [CrossRef]
- Roberts, F. A systematic study of the Australian species of the genus Haemaphysalis Koch (Acarina: Ixodidae). Aust. J. Zool. 1963, 11, 35–80. [Google Scholar] [CrossRef]
- Roberts, F.H.S. Australian Ticks; CSIRO: Melbourne, Australia, 1970; 267p. [Google Scholar]
- Laan, B.; Handasyde, K.; Beveridge, I. Occurrence of the tick Haemaphysalis bancrofti Nuttall & Warburton, 1915 in Victoria with additional data on its distribution and with scanning electron micrographs of life cycle stages. Proc. R. Soc. Vic. 2011, 123, 189. [Google Scholar]
- Callow, L.L.; Hoyte, H.M.D. Transmission experiments using Babesia bigemina, Theileria mutans, Borrelia sp.; the cattle tick. Aust. Vet. J. 1961, 37, 381–390. [Google Scholar] [CrossRef]
- Onzere, C.K.; Herndon, D.R.; Hassan, A.; Oyen, K.; Poh, K.C.; Scoles, G.A.; Fry, L.M. A U.S. Isolate of Theileria orientalis Ikeda Is Not Transstadially Transmitted to Cattle by Rhipicephalus microplus. Pathogens 2023, 12, 559. [Google Scholar] [CrossRef]
- Foil, L.D.; Gorham, J.R. Mechanical Transmission of Disease Agents by Arthropods, in Medical Entomology; Springer: Berlin/Heidelberg, Germany, 2004; pp. 461–514. [Google Scholar]
- Biddle, A.; Eastwood, S.; Martin, L.; Freeman, P.; Druce, E. A survey to determine the prevalence of Theileria spp. in beef cattle in the northern tablelands of New South Wales. Aust. Vet. J. 2013, 91, 427–431. [Google Scholar] [CrossRef]
- Hammer, J.F.; Jenkins, C.; Bogema, D.; Emery, D. Mechanical transfer of Theileria orientalis: Possible roles of biting arthropods, colostrum and husbandry practices in disease transmission. Parasites Vectors 2016, 9, 1–9. [Google Scholar] [CrossRef]
- Fujisaki, K.; Kamio, T.; Kawazu, S.; Shimizu, S.; Shimura, K. Theileria sergenti: Experimental transmission by the long-nosed cattle louse, Linognathus vituli. Ann. Trop. Med. Parasitol. 1993, 87, 217–218. [Google Scholar] [CrossRef] [PubMed]
- De Marco, M.F.; Brugman, V.; Hernández-Triana, L.; Thorne, L.; Phipps, L.; Nikolova, N.; Fooks, A.; Johnson, N. Detection of Theileria orientalis in mosquito blood meals in the United Kingdom. Vet. Parasitol. 2016, 229, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Changbunjong, T.; Sungpradit, S.; Kanthasaewee, O.; Sedwisai, P.; Tangsudjai, S.; Ruangsittichai, J. Molecular Detection of Theileria and Babesia in a Diversity of Stomoxyini Flies (Diptera: Muscidae) from Khao Yai National Park, Thailand. Thai J. Vet. Med. 2016, 46, 227–234. [Google Scholar] [CrossRef]
- Hornok, S.; Takács, N.; Szekeres, S.; Szőke, K.; Kontschán, J.; Horváth, G.; Sugár, L. DNA of Theileria orientalis, T. equi and T. capreoli in stable flies (Stomoxys calcitrans). Parasites Vectors 2020, 13, 1–6. [Google Scholar] [CrossRef]
- Jirapattharasate, C.; Changbunjong, T.; Sedwisai, P.; Weluwanarak, T. Molecular detection of piroplasms in haematophagus flies in the Nakhon Pathom and Kanchanaburi Provinces, Thailand. Vet. Integr. Sci. 2018, 16, 123–133. [Google Scholar]
- Foil, L.C.; Foil, C.S. Dipteran parasites of horses. Equine Pract. 1988, 10, 21–38. [Google Scholar]
- Foil, L.; Stage, D.; Adams, W.V.; Issel, C.J. Observations of tabanid feeding on mares and foals. Am. J. Vet. Res. 1985, 46, 1111–1113. [Google Scholar]
- Hollander, A.L.; Wright, R.E. Impact of Tahanids on Cattle: Blood Meal Size and Preferred Feeding Sites. J. Econ. Entomol. 1980, 73, 431–433. [Google Scholar] [CrossRef]
- Muzari, M.; Jones, R.; Skerratt, L.; Duran, T. Feeding success and trappability of horse flies evaluated with electrocuting nets and odour-baited traps. Vet. Parasitol. 2010, 171, 321–326. [Google Scholar] [CrossRef]
- Muzari, M.; Skerratt, L.; Jones, R.; Duran, T. Alighting and feeding behaviour of tabanid flies on horses, kangaroos and pigs. Vet. Parasitol. 2010, 170, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Foil, L. Tabanids as vectors of disease agents. Parasitol. Today 1989, 5, 88–96. [Google Scholar] [CrossRef]
- Barros, A.; Foil, L. The influence of distance on movement of tabanids (Diptera: Tabanidae) between horses. Vet. Parasitol. 2007, 144, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Foil, L.; Hogsette, J. Biology and control of tabanids, stable flies and horn flies. Rev. Sci. Tech.-Off. Int. Des Épizooties 1994, 13, 1125–1158. [Google Scholar] [CrossRef]
- Schowalter, T.; Klowden, M. Blood Meal Size of the Stable Fly, Stomoxys Calcitrans, Measured by the HiCN [Hemoglobin-Cyanide] Method. Mosq. News 1979, 39, 110–112. [Google Scholar]
- Holdsworth, P.A. Use of Macrocyclic Lactones to Control Cattle Parasites in Australia and New Zealand; CAB International: Wallingford, UK, 2002; pp. 288–301. [Google Scholar] [CrossRef]
- Bailey, G. Cattle Lice, Primefact 337, 2nd ed.; NSW Department of Primary Industries: Orange, Australia, 2015; pp. 1–4. [Google Scholar]
- Yu, Y.; Liu, J.; Liu, G.; Liu, Z.; Hao, B.; Yan, G.; Zhao, T. Ceratopogonidae of China. Insecta Diptera 2005, 1, 2. [Google Scholar]
- Venter, G.; Hamblin, C.; Paweska, J. Determination of the oral susceptibility of South African livestock-associated biting midges, Culicoides species, to bovine ephemeral fever virus. Med. Vet. Entomol. 2003, 17, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Standfast, H.A.; Dyce, A. Potential vectors of arboviruses of cattle and buffalo in Australia. Aust. Vet. J. 1972, 48, 224–227. [Google Scholar] [CrossRef]
- Bishop, A.L.; Barchia, I.M.; Spohr, L.J. Models for the dispersal in Australia of the arbovirus vector, Culicoides brevitarsis Kieffer (Diptera: Ceratopogonidae). Prev. Vet. Med. 2000, 47, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, K.; Gedye, K.; Pomroy, W. A longitudinal study of the effect of Theileria orientalis Ikeda type infection on three New Zealand dairy farms naturally infected at pasture. Vet. Parasitol. 2019, 276, 108977. [Google Scholar] [CrossRef]
- Baek, B.K.; Soo, K.B.; Kim, J.H.; Hur, J.; O Lee, B.; Jung, J.M.; Onuma, M.; O Oluoch, A.; Kim, C.-H.; Kakoma, I. Verification by polymerase chain reaction of vertical transmission of Theileria sergenti in cows. Can. J. Vet. Res. = Rev. Can. Rech. Vet. 2003, 67, 278–282. [Google Scholar]
- Lawrence, K.; Gedye, K.; McFadden, A.; Pulford, D.; Pomroy, W. An observational study of the vertical transmission of Theileria orientalis (Ikeda) in a New Zealand pastoral dairy herd. Vet. Parasitol. 2016, 218, 59–65. [Google Scholar] [CrossRef]
- Mekata, H.; Minamino, T.; Mikurino, Y.; Yamamoto, M.; Yoshida, A.; Nonaka, N.; Horii, Y. Evaluation of the natural vertical transmission of Theileria orientalis. Vet. Parasitol. 2018, 263, 1–4. [Google Scholar] [CrossRef]
- Reber, A.; Donovan, D.; Gabbard, J.; Galland, K.; Aceves-Avila, M.; Holbert, K.; Marshall, L.; Hurley, D. Transfer of maternal colostral leukocytes promotes development of the neonatal immune system: Part II. Effects on neonatal lymphocytes. Vet. Immunol. Immunopathol. 2008, 123, 305–313. [Google Scholar] [CrossRef]
- Donovan, D.C.; Reber, A.J.; Gabbard, J.D.; Aceves-Avila, M.; Galland, K.L.; Holbert, K.A.; Ely, L.O.; Hurley, D.J. Effect of maternal cells transferred with colostrum on cellular responses to pathogen antigens in neonatal calves. Am. J. Vet. Res. 2007, 68, 778–782. [Google Scholar] [CrossRef]
- Emery, D.L.; I Morrison, W.; Buscher, G.; Nelson, R.T. Generation of cell-mediated cytotoxicity to Theileria parva (East Coast fever) after inoculation of cattle with parasitized lymphoblasts. Pediatrics 1982, 128, 195–200. [Google Scholar] [CrossRef]
- Goddeeris, B.M.; Morrison, W.I. Cell-Mediated Immunity in Ruminants; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]

{kind=link}
{kind=link}
| Country | Tick Species | Source of Ticks | T. orientalis MPSP Genotypes Detected | References |
|---|---|---|---|---|
| Australia | H. longicornis | engorged ticks | Type 1, 2, and 3 | [27,70] |
| H. bancrofti | questing ticks | Type 1 and 3 | ||
| Japan | H. megaspinosa, | questing ticks | Type 1 and 2 | [71] |
| H. douglasi, | ||||
| Ixodes persulcatus, | ||||
| I. ovatus | ||||
| Mongolia | Dermacentor nuttalli | engorged ticks | Types 1, 3, 5, 7, and N-3 | [72] |
| Ethiopia | R. evertsi evertsi | engorged ticks | Type 3 | [73,74] |
| R. decoloratus | ||||
| Rh. praetextatus | ||||
| Turkey | Hyalomma excavatum Boophilus annulatus | engorged ticks | Type 3 | [75] |
| Vietnam | Boophilus microplus | engorged ticks | Types 5 and 7 | [14] |
| China | R. microplus | engorged ticks and questing ticks | Types 1–5, 7, and N3 | [76] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lakew, B.T.; Eastwood, S.; Walkden-Brown, S.W. Epidemiology and Transmission of Theileria orientalis in Australasia. Pathogens 2023, 12, 1187. https://doi.org/10.3390/pathogens12101187
Lakew BT, Eastwood S, Walkden-Brown SW. Epidemiology and Transmission of Theileria orientalis in Australasia. Pathogens. 2023; 12(10):1187. https://doi.org/10.3390/pathogens12101187
Chicago/Turabian StyleLakew, Biniam T., Steve Eastwood, and Stephen W. Walkden-Brown. 2023. "Epidemiology and Transmission of Theileria orientalis in Australasia" Pathogens 12, no. 10: 1187. https://doi.org/10.3390/pathogens12101187
APA StyleLakew, B. T., Eastwood, S., & Walkden-Brown, S. W. (2023). Epidemiology and Transmission of Theileria orientalis in Australasia. Pathogens, 12(10), 1187. https://doi.org/10.3390/pathogens12101187