
Cytauxzoon felis: An Overview



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. History
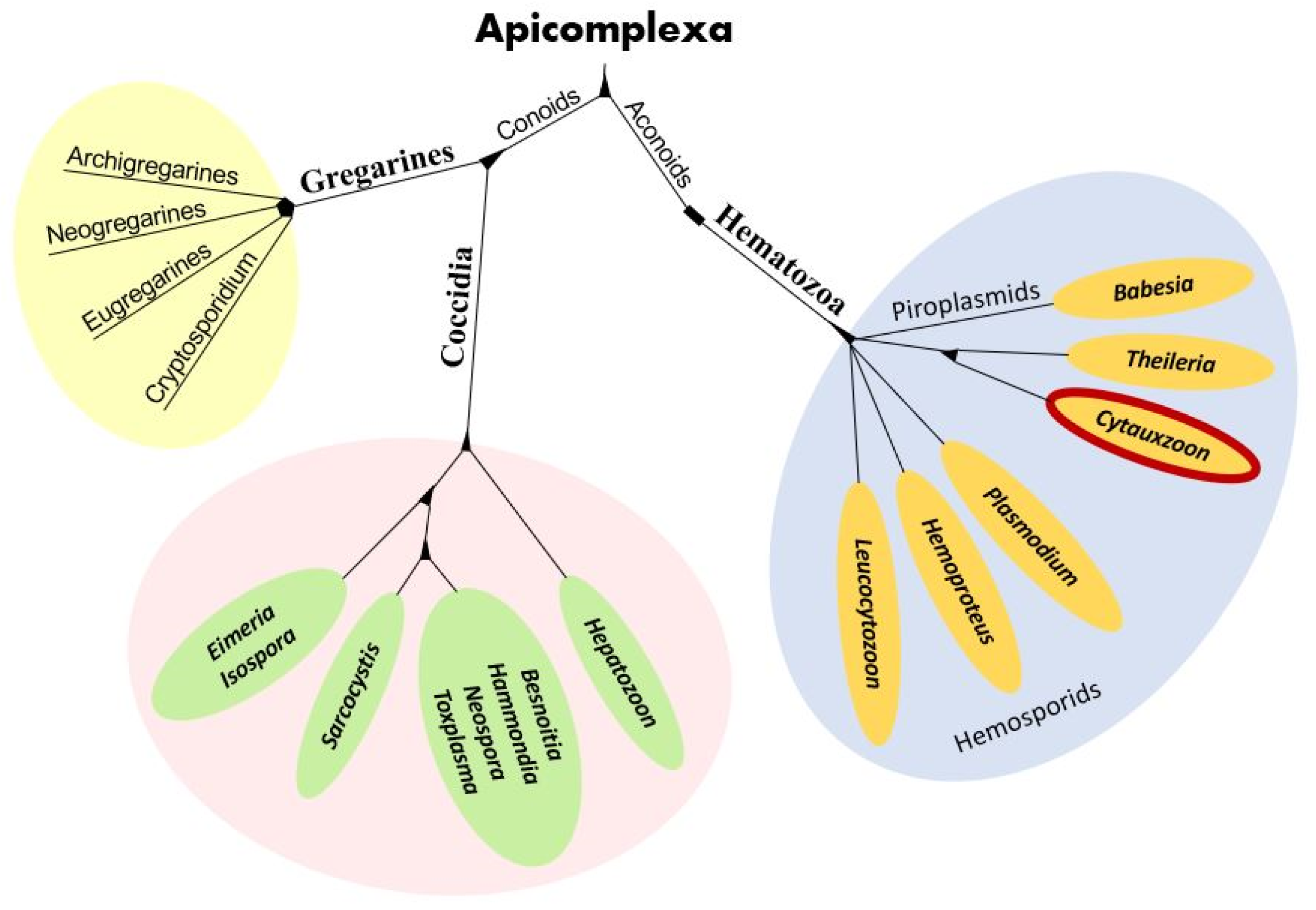
3. Phylogeny
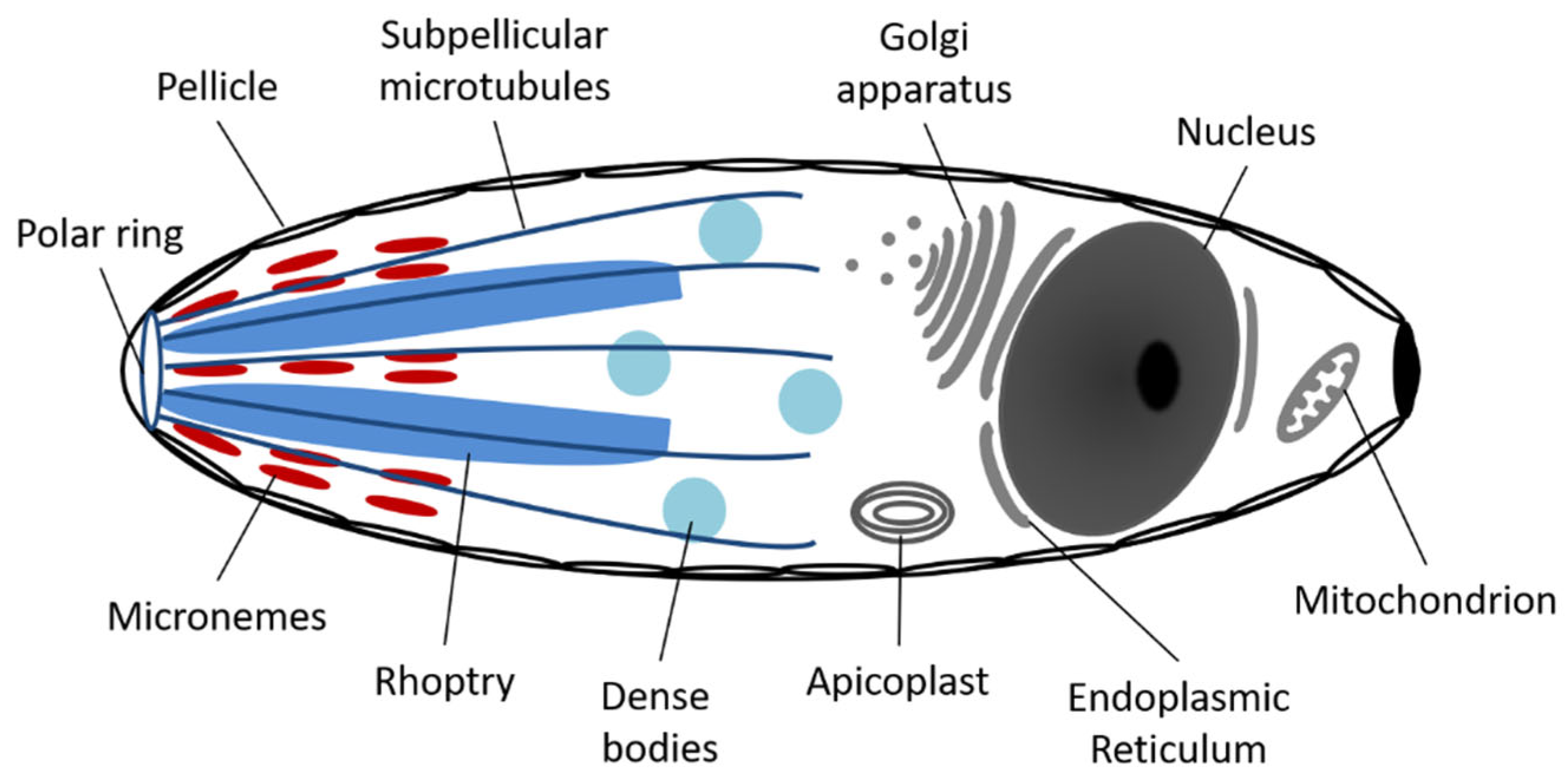
4. Anatomy
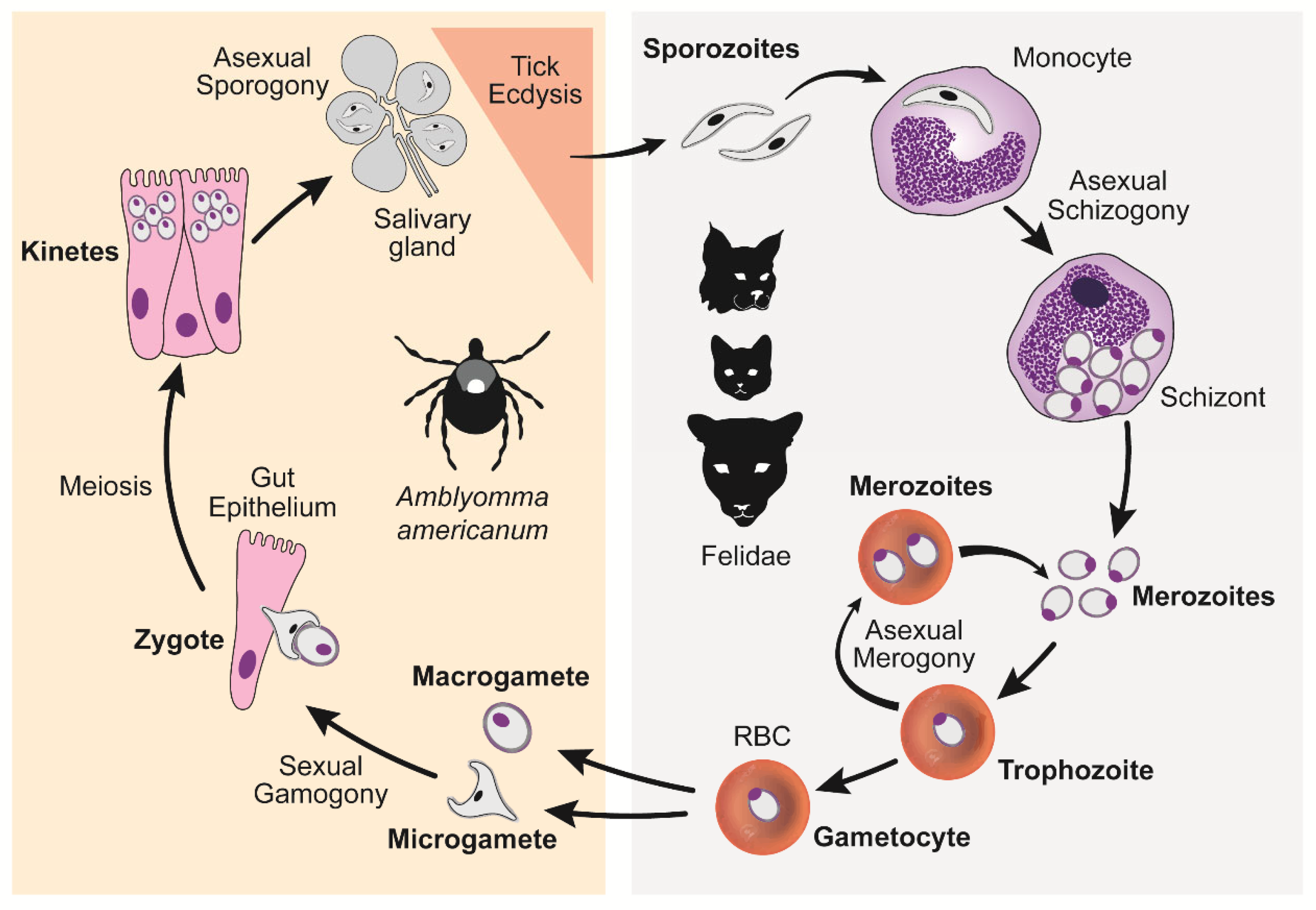
5. Life Cycle
5.1. Summary
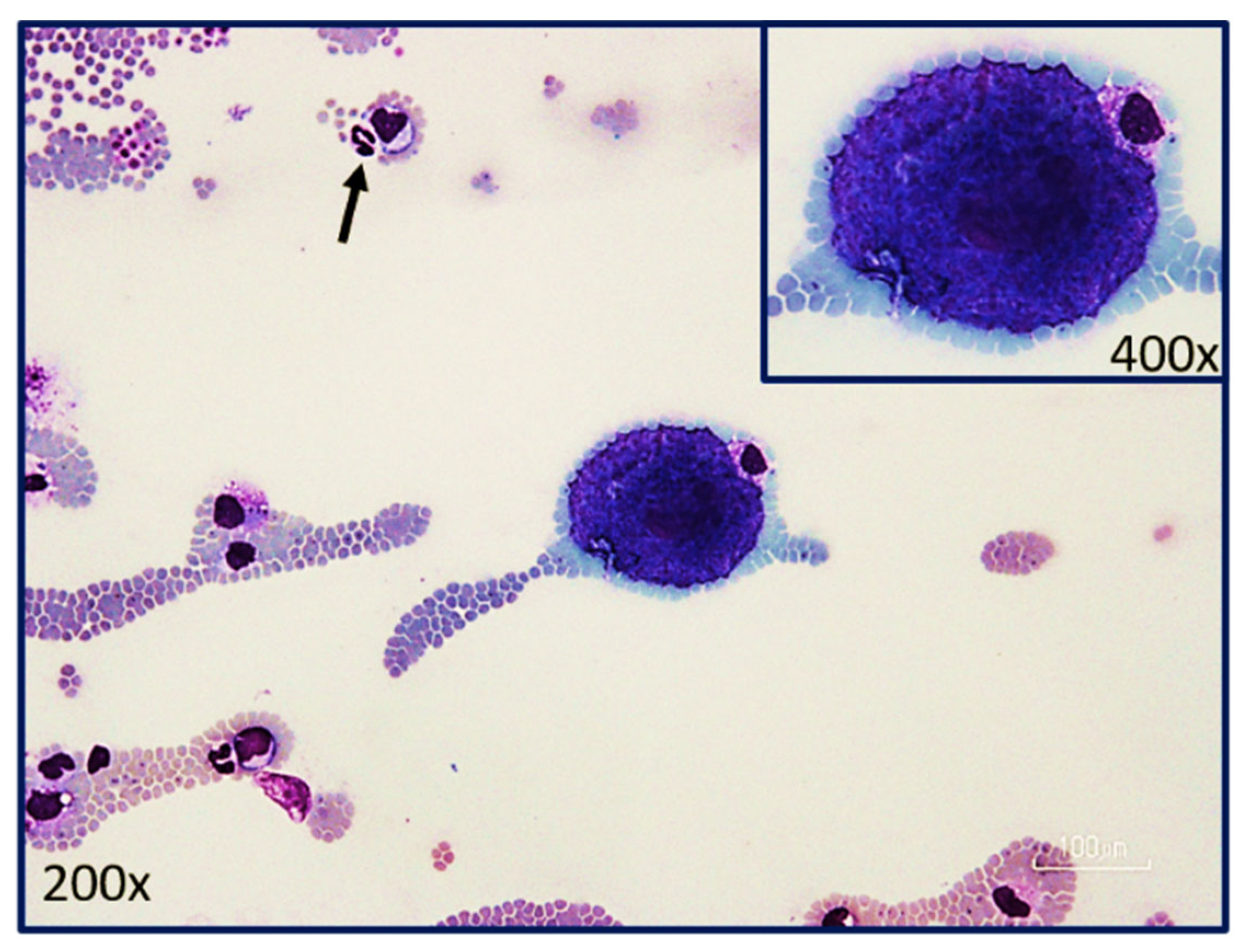
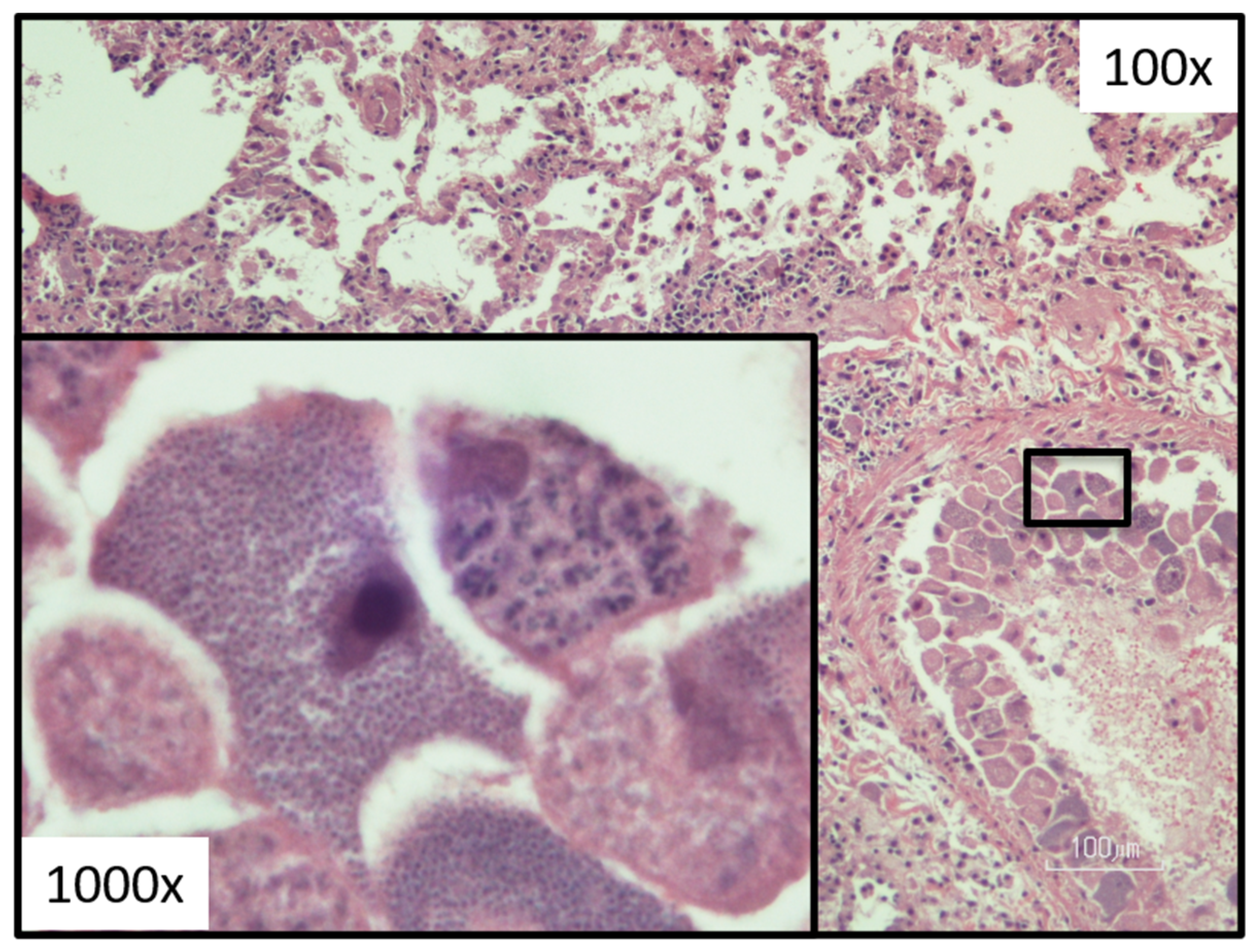
5.2. Asexual Schizogony
5.3. Asexual Merogony
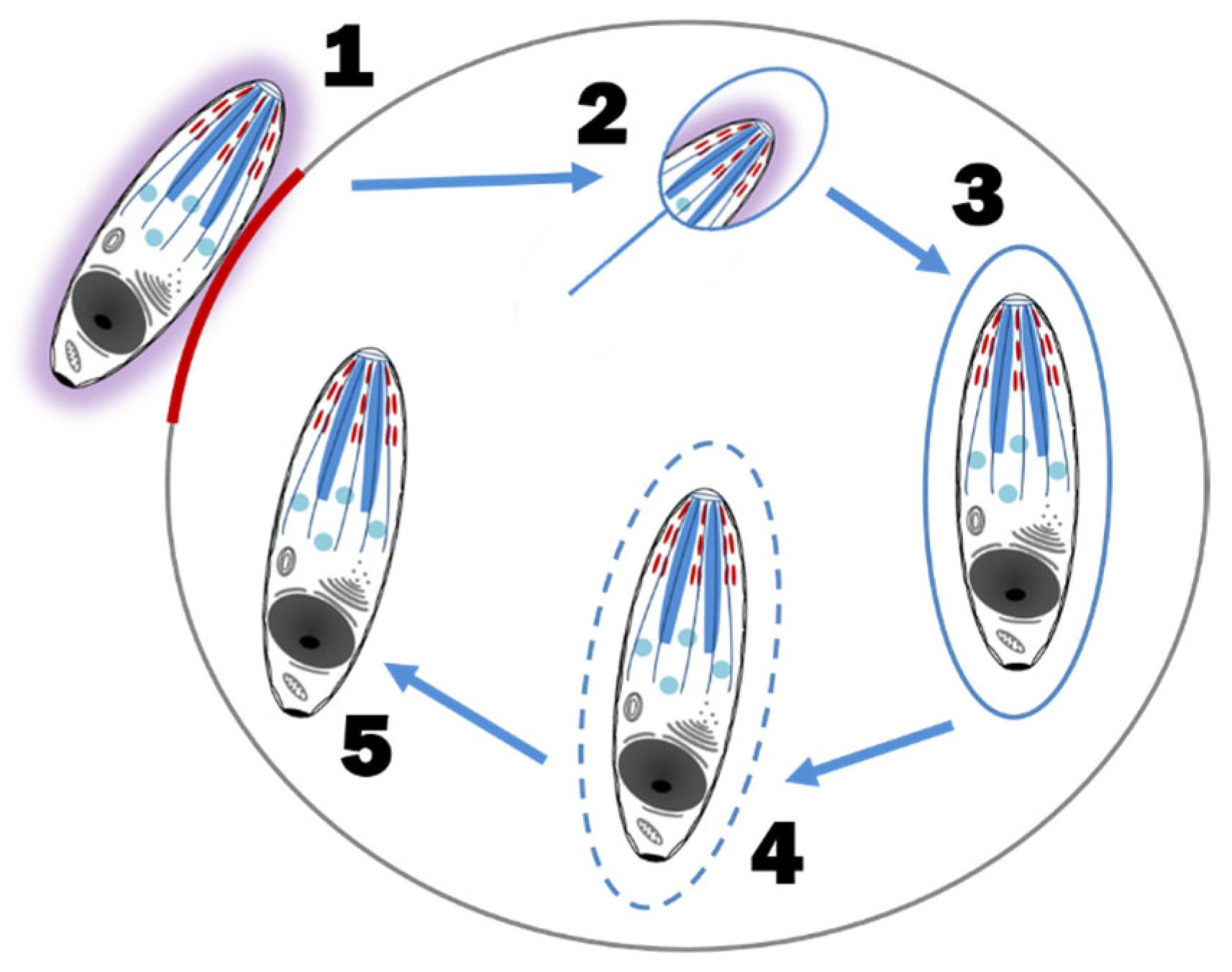
5.4. Sexual Reproduction
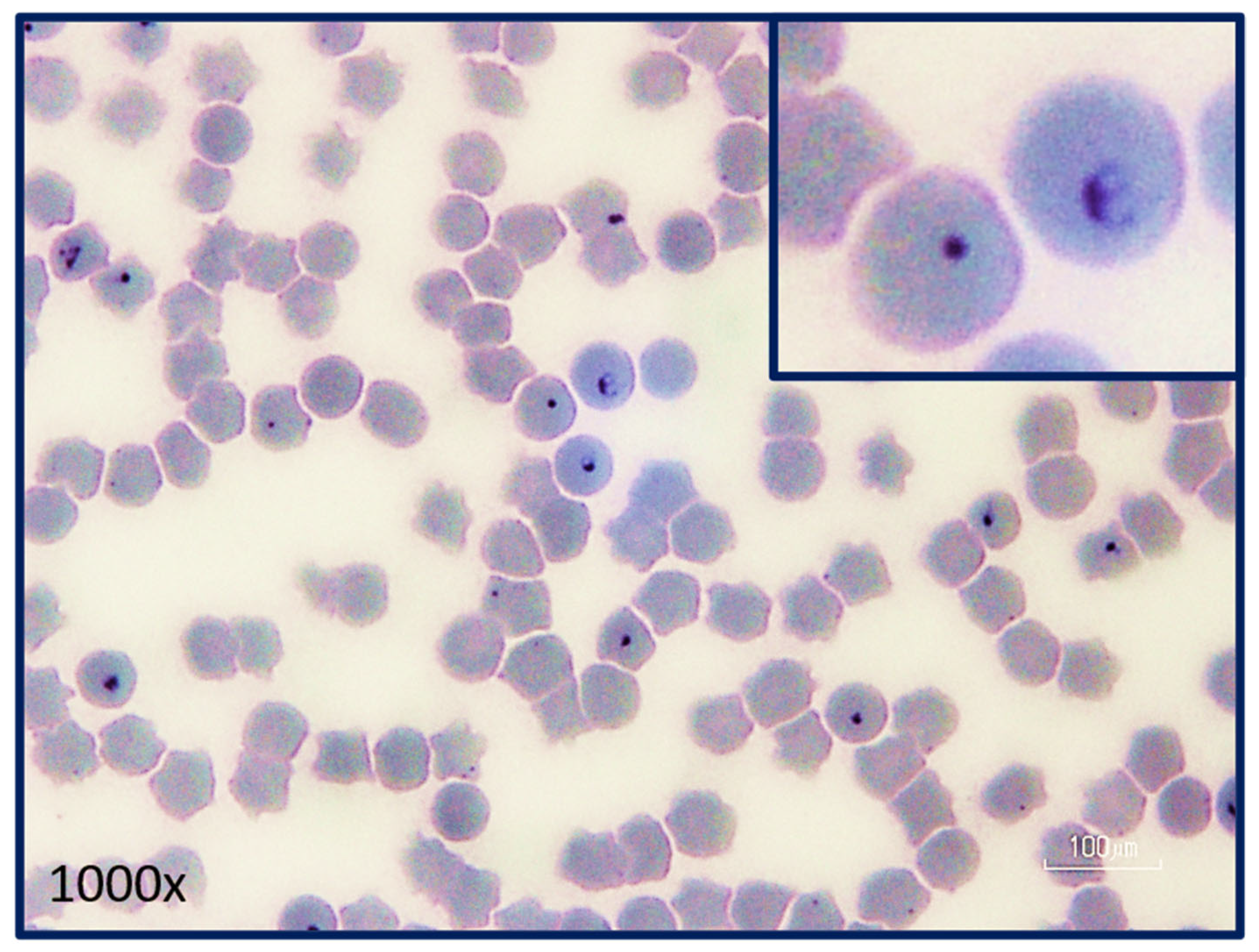
5.5. Asexual Sporogony
6. Transmission Vectors & Definitive Host
6.1. Amblyomma americanum
6.2. Dermacentor variabilis
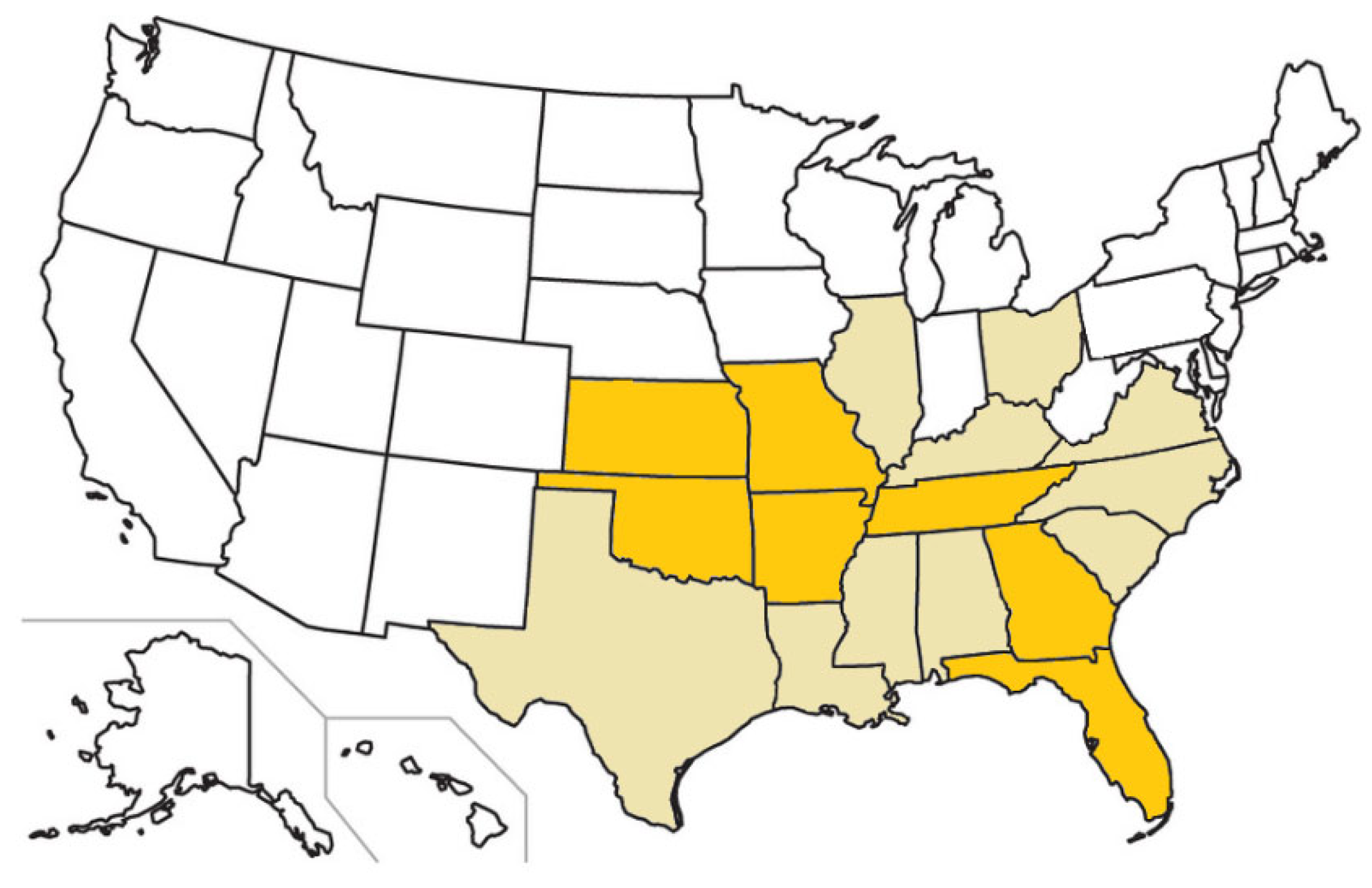
7. Felid Intermediate Hosts
8. Cytauxzoonosis
8.1. Acute Disease
8.2. Subclinical Disease
9. Diagnosis
10. Treatment Options
11. Control & Prevention
12. Summary & Future Considerations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Panait, L.C.; Mihalca, A.D.; Modry, D.; Jurankova, J.; Ionica, A.M.; Deak, G.; Gherman, C.M.; Heddergott, M.; Hodzic, A.; Veronesi, F.; et al. Three new species of Cytauxzoon in European wild felids. Vet. Parasitol. 2021, 290, 109344. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Li, T.T.; Liu, G.H.; Zhu, X.Q.; Yao, C. Two Tales of Cytauxzoon felis Infections in Domestic Cats. Clin. Microbiol. Rev. 2017, 30, 861–885. [Google Scholar] [CrossRef] [PubMed]
- Reichard, M.E.; Sanders, T.L.; Weerarathne, P.; Meinkoth, J.H.; Miller, C.A.; Scimeca, R.C.; Almazán, C. Cytauxzoonosis in North America. Pathogens 2021, 10, 1170. [Google Scholar] [CrossRef] [PubMed]
- Jalovecka, M.; Hajdusek, O.; Sojka, D.; Kopacek, P.; Malandrin, L. The Complexity of Piroplasms Life Cycles. Front. Cell. Infect. Microbiol. 2018, 8, 248. [Google Scholar] [CrossRef]
- Wikander, Y.; Anantatat, T.; Kang, Q.; Reif, K.R. Prevalence of Cytauxzoon felis Infection-Carriers in Eastern Kansas Domestic Cats. Pathogens 2020, 9, 854. [Google Scholar] [CrossRef]
- Neitz, W.O.; Thomas, A.D. Cytauxzoon sylvicaprae Gen. Nov., Spec. Nov., a Protozoon Responsible for a Hitherto Undescribed Disease in the Duiker, Sylvicapra grimmia (Linné). Onderstepoort J. Vet. Sci. Anim. Ind. 1948, 23, 63–76. [Google Scholar]
- Wagner, J.E. A Fatal Cytauxzoonosis like Disease in Cats. J. Am. Vet. Med. Assoc. 1976, 168, 585–588. [Google Scholar]
- Ferris, D.H. A Progress Report on the Status of a New Disease of American Cats: Cytauxzoonosis. Comp. Immunol. Microbiol. Infect. Dis. 1979, 1, 269–276. [Google Scholar] [CrossRef]
- Nijhof, A.M.; Pillay, V.; Steyl, J.; Prozesky, L.; Stoltsz, W.H.; Lawrence, J.A.; Penzhorn, B.L.; Jongejan, F. Molecular Characterization of Theileria Species Associated with Mortality in Four Species of African Antelopes. J. Clin. Microbiol. 2005, 43, 5907–5911. [Google Scholar] [CrossRef]
- Kier, A.B.; Wightman, S.R.; Wagner, J.E. Interspecies Transmission of Cytauxzoon felis. Am. J. Vet. Res. 1982, 43, 102–105. [Google Scholar]
- Schreeg, M.E.; Marr, H.S.; Tarigo, J.L.; Cohn, L.A.; Bird, D.M.; Scholl, E.H.; Levy, M.G.; Wiegmann, B.M.; Birkenheuer, A.J. Mitochondrial Genome Sequences and Structures Aid in the Resolution of Piroplasmida Phylogeny. PLoS ONE 2016, 11, e0165702. [Google Scholar] [CrossRef] [PubMed]
- O’Donoghue, P. Haemoprotozoa: Making Biological Sense of Molecular Phylogenies. Int. J. Parasitol. Parasites Wildl. 2017, 6, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Ascencio, M.; Florin-Christensen, M.; Mamoun, C.; Weir, W.; Shiels, B.; Schnittger, L. Cysteine Proteinase C1A Paralog Profiles Correspond with Phylogenetic Lineages of Pathogenic Piroplasmids. Vet. Sci. 2018, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.J.; Williamson, D.H. Extrachromosomal DNA in the Apicomplexa. Microbiol. Mol. Biol. Rev. 1997, 61, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Tran, J.Q.; de Leon, J.C.; Li, C.; Huynh, M.-H.; Beatty, W.; Morrissette, N.S. RNG1 Is a Late Marker of the Apical Polar Ring in Toxoplasma gondii. Cytoskeleton 2010, 67, 586–598. [Google Scholar] [CrossRef]
- Sibley, L.D. How Apicomplexan Parasites Move in and out of Cells. Curr. Opin. Biotechnol. 2010, 21, 592–598. [Google Scholar] [CrossRef]
- Alexeyev, M.; Shokolenko, I.; Wilson, G.; LeDoux, S. The Maintenance of Mitochondrial DNA Integrity—Critical Analysis and Update. Cold Spring Harb. Perspect. Biol. 2013, 5, a012641. [Google Scholar] [CrossRef]
- Mather, M.; Henry, K.; Vaidya, A. Mitochondrial Drug Targets in Apicomplexan Parasites. Curr. Drug Targets 2006, 8, 49–60. [Google Scholar] [CrossRef]
- Schreeg, M.E.; Marr, H.S.; Griffith, E.H.; Tarigo, J.L.; Bird, D.M.; Reichard, M.V.; Cohn, L.A.; Levy, M.G.; Birkenheuer, A.J. PCR Amplification of a Multi-Copy Mitochondrial Gene (Cox3) Improves Detection of Cytauxzoon felis Infection as Compared to a Ribosomal Gene (18S). Vet. Parasitol. 2016, 225, 123–130. [Google Scholar] [CrossRef]
- Kocan, A.A.; Kocan, K.M.; Blouin, E.F.; Mukolwe, S.W. A Redescription of Schizogony of Cytauxzoon Felis in the Domestic Cat. Ann. N. Y. Acad. Sci. 1992, 653, 161–167. [Google Scholar] [CrossRef]
- Snider, T.A.; Confer, A.W.; Payton, M.E. Pulmonary Histopathology of Cytauxzoon felis Infections in the Cat. Vet. Pathol. 2010, 47, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Lehane, M.J. Peritrophic Matrix Structure and Function. Annu. Rev. Entomol. 1997, 42, 525–550. [Google Scholar] [CrossRef]
- Hegedus, D.; Erlandson, M.; Gillott, C.; Toprak, U. New Insights into Peritrophic Matrix Synthesis, Architecture, and Function. Annu. Rev. Entomol. 2009, 54, 285–302. [Google Scholar] [CrossRef] [PubMed]
- Bolognesi, R.; Terra, W.R.; Ferreira, C. Peritrophic Membrane Role in Enhancing Digestive Efficiency. Theoretical and Experimental Models. J. Insect Physiol. 2008, 54, 1413–1422. [Google Scholar] [CrossRef] [PubMed]
- Reichard, M.V.; Edwards, A.C.; Meinkoth, J.H.; Snider, T.A.; Meinkoth, K.R.; Heinz, R.E.; Little, S.E. Confirmation of Amblyomma americanum (Acari: Ixodidae) as a Vector for Cytauxzoon felis (Piroplasmorida: Theileriidae) to Domestic Cats. J. Med. Entomol. 2010, 47, 890–896. [Google Scholar] [CrossRef]
- Thomas, J.E.; Ohmes, C.M.; Payton, M.E.; Hostetler, J.A.; Reichard, M.V. Minimum Transmission Time of Cytauxzoon felis by Amblyomma americanum to Domestic Cats in Relation to Duration of Infestation, and Investigation of Ingestion of Infected Ticks as a Potential Route of Transmission. J. Feline Med. Surg. 2018, 20, 67–72. [Google Scholar] [CrossRef]
- Allen, K.E.; Thomas, J.E.; Wohltjen, M.L.; Reichard, M.V. Transmission of Cytauxzoon felis to Domestic Cats by Amblyomma americanum Nymphs. Parasites Vectors 2019, 12, 28. [Google Scholar] [CrossRef]
- Nagamori, Y.; Slovak, J.E.; Reichard, M.V. Prevalence of Cytauxzoon felis Infection in Healthy Free-Roaming Cats in North-Central Oklahoma and Central Iowa. J. Feline Med. Surg. Open Rep. 2016, 2, 363. [Google Scholar] [CrossRef]
- Holderman, C.J.; Kaufman, P.E. Common Name: Lone Star Tick Scientific Name: Amblyomma americanum (Linnaeus) (Acari: Ixodidae). Available online: http://entnemdept.ufl.edu/creatures/urban/medical/lone_star_tick.htm (accessed on 15 March 2020).
- Adams, D.; Anderson, B.; Ammirati, C.; Helm, K. Identification And Diseases Of Common U.S. Ticks. Internet J. Dermatol. 2002, 2. Available online: http://ispub.com/IJD/2/1/7515 (accessed on 16 March 2020).
- Nuttall, P.A. Tick Saliva and Its Role in Pathogen Transmission. Wien. Klin. Wochenschr. Cental Eur. J. Med. 2019, 131. [Google Scholar] [CrossRef]
- Chmelar, J.; Kotál, J.; Kovaríková, A.; Kotsyfakis, M. The Use of Tick Salivary Proteins as Novel Therapeutics. Front. Physiol. 2019, 10, 812. [Google Scholar] [CrossRef]
- Chan, W.-H.; Kaufman, P.E. Common Name: American Dog Tick Scientific Name: Dermacentor variabilis (Say) (Arachnida: Ixodida: Ixodidae). Available online: http://entnemdept.ufl.edu/creatures/urban/medical/american_dog_tick.htm (accessed on 24 March 2020).
- Monzón, J.D.; Atkinson, E.G.; Henn, B.M.; Benach, J.L. Population and Evolutionary Genomics of Amblyomma americanum, an Expanding Arthropod Disease Vector. Genome Biol. Evol. 2016, 8, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.N.; Sundstrom, K.D.; Duncan, K.T.; Ientile, M.M.; Jordy, J.; Ghosh, P.; Little, S.E. Show Us Your Ticks: A Survey of Ticks Infesting Dogs and Cats across the USA. Parasites Vectors 2019, 12, 595. [Google Scholar] [CrossRef] [PubMed]
- Little, S.E.; Barrett, A.W.; Nagamori, Y.; Herrin, B.H.; Normile, D.; Heaney, K.; Armstrong, R. Ticks from Cats in the United States: Patterns of Infestation and Infection with Pathogens. Vet. Parasitol. 2018, 257, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, E.L.; Stone, N.E.; Scoles, G.A.; Hepp, C.M.; Busch, J.D.; Wagner, D.M. Range-Wide Genetic Analysis of Dermacentor variabilis and Its Francisella-like Endosymbionts Demonstrates Phylogeographic Concordance between Both Taxa. Parasites Vectors 2018, 11, 306. [Google Scholar] [CrossRef]
- Minigan, J.N.; Hager, H.A.; Peregrine, A.S.; Newman, J.A. Current and Potential Future Distribution of the American Dog Tick (Dermacentor variabilis, Say) in North America. Ticks Tick. Borne. Dis. 2018, 9, 354–362. [Google Scholar] [CrossRef]
- Blouin, E.F.; Kocan, A.A.; Glenn, B.L.; Kocan, K.M.; Hair, J.A. Transmission of Cytauxzoon felis Kier, 1979 from Bobcats, Felis rufus (Schreber), to Domestic Cats by Dermacentor veriabilis (Say). J. Wildl. Dis. 1984, 20, 241–242. [Google Scholar] [CrossRef]
- Reichard, M.V.; Meinkoth, J.H.; Edwards, A.C.; Snider, T.A.; Kocan, K.M.; Blouin, E.F.; Little, S.E. Transmission of Cytauxzoon felis to a Domestic Cat by Amblyomma americanum. Vet. Parasitol. 2009, 161, 110–115. [Google Scholar] [CrossRef]
- Glenn, B.L.; Kocan, A.A.; Blouin, E.F. Cytauxzoonosis in Bobcats. J. Am. Vet. Med. Assoc. 1983, 183, 1155–1158. [Google Scholar]
- Blouin, E.F.; Kocan, A.A.; Kocan, K.M.; Hair, J. Evidence of a Limited Schizogonous Cycle for Cytauxzoon felis in Bobcats Following Exposure to Infected Ticks. J. Wildl. Dis. 1987, 23, 499–501. [Google Scholar] [CrossRef]
- Birkenheuer, A.J.; Marr, H.S.; Warren, C.; Acton, A.E.; Mucker, E.M.; Humphreys, J.G.; Tucker, M.D. Cytauxzoon felis Infections Are Present in Bobcats (Lynx rufus) in a Region Where Cytauxzoonosis Is Not Recognized in Domestic Cats. Vet. Parasitol. 2008, 153, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Zieman, E.A.; Nielsen, C.K.; Jiménez, F.A. Chronic Cytauxzoon felis Infections in Wild-Caught Bobcats (Lynx rufus). Vet. Parasitol. 2018, 252, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Shock, B.C.; Murphy, S.M.; Patton, L.L.; Shock, P.M.; Olfenbuttel, C.; Beringer, J.; Prange, S.; Grove, D.M.; Peek, M.; Butfiloski, J.W.; et al. Distribution and Prevalence of Cytauxzoon felis in Bobcats (Lynx rufus), the Natural Reservoir, and Other Wild Felids in Thirteen States. Vet. Parasitol. 2011, 175, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.M.; Lockhart, J.M.; Latimer, K.S.; Peterson, D.S. Identification and Genetic Characterization of Cytauxzoon felis in Asymptomatic Domestic Cats and Bobcats. Vet. Parasitol. 2010, 172, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.M.; Cohn, L.A.; Marr, H.S.; Birkenheuer, A.J. Diminazene Diaceturate for Treatment of Chronic Cytauxzoon felis Parasitemia in Naturally Infected Cats. J. Vet. Intern. Med. 2012, 26, 1490–1493. [Google Scholar] [CrossRef] [PubMed]
- Butt, M.T.; Bowman, D.; Barr, M.C.; Roelke, M.E. Iatrogenic Transmistion of Cytauxzoon felis from a Florida Panther (Felix concolor coryi) to a Domestic Cat. J. Wildl. Dis. 1991, 27, 342–347. [Google Scholar] [CrossRef]
- Rotstein, D.S.; Taylor, S.K.; Harvey, J.W.; Bean, J. Hematologic Effects of Cytauxzoonosis in Florida Panthers and Texas Cougars in Florida. J. Wildl. Dis. 1999, 35, 613–617. [Google Scholar] [CrossRef]
- Harvey, J.W.; Dunbar, M.R.; Norton, T.M.; Yabsley, M.J. Laboratory Findings in Acute Cytauxzoon felis Infection in Cougars (Puma concolor couguar) in Florida. J. Zoo Wildl. Med. 2007, 38, 285–291. [Google Scholar] [CrossRef]
- Lewis, K.M.; Cohn, L.A.; Downey, M.E.; Whitney, M.S.; Birkenheuer, A.J. Evaluation of Cytauxzoon felis Infection Status in Captive-Born Wild Felids Housed in an Area Endemic for the Pathogen. J. Am. Vet. Med. Assoc. 2012, 241, 1088–1092. [Google Scholar] [CrossRef]
- Cerreta, A.J.; Yang, T.S.; Ramsay, D.C.; Birkenheuer, A.J.; Rahoi, D.; Qurollo, B.; Wilsn, J.; Cushing, A.C. Detection of Vector-borne Infections in Lions and Tigers at Two Zoos in Tennessee and Oklahoma, USA. J. Zoo Wildl. Med. 2022, 53, 50–59. [Google Scholar] [CrossRef]
- Rizzi, T.E.; Reichard, M.V.; Cohn, L.A.; Birkenheuer, A.J.; Taylor, J.D.; Meinkoth, J.H. Prevalence of Cytauxzoon felis Infection in Healthy Cats from Enzootic Areas in Arkansas, Missouri, and Oklahoma. Parasites Vectors 2015, 8, 13. [Google Scholar] [CrossRef]
- Nietfeld, J.C.; Pollock, C. Fatal Cytauxzoonosis in a Free-Ranging Bobcat (Lynx rufus). J. Wildl. Dis. 2002, 38, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Hoover, J.P.; Walker, D.B.; Hedges, J.D. Cytauxzoonosis in Cats: Eight Cases (1985–1992). J. Am. Vet. Med. Assoc. 1994, 205, 455–460. [Google Scholar] [PubMed]
- Meinkoth, J.H.; Kocan, A.A. Feline Cytauxzoonosis. Vet. Clin. N. Am. Small Anim. Pract. 2005, 35, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Meinkoth, J.; Kocan, A.A.; Whitworth, L.; Murphy, G.; Fox, J.C.; Woods, J.P. Cats Surviving Natural Infection with Cytauxzoon felis: 18 Cases (1997-1998). J. Vet. Intern. Med. 2000, 14, 521–525. [Google Scholar] [CrossRef]
- Jackson, C.B.; Fisher, T. Fatal Cytauxzoonosis in a Kentucky Cat (Felis Domesticus). Vet. Parasitol. 2006, 139, 192–195. [Google Scholar] [CrossRef]
- Haber, M.D.; Tucker, M.D.; Marr, H.S.; Levy, J.K.; Burgess, J.; Lappin, M.R.; Birkenheuer, A.J. The Detection of Cytauxzoon felis in Apparently Healthy Free-Roaming Cats in the USA. Vet. Parasitol. 2007, 146, 316–320. [Google Scholar] [CrossRef]
- Brown, H.M.; Latimer, K.S.; Erikson, L.E.; Cashwell, M.E.; Britt, J.O.; Peterson, D.S. Detection of Persistent Cytauxzoon felis Infection by Polymerase Chain Reaction in Three Asymptomatic Domestic Cats. J. Vet. Diagn. Investig. 2008, 20, 485–488. [Google Scholar] [CrossRef]
- MacNeill, A.L.; Barger, A.M.; Skowronski, M.C.; Lanka, S.; Maddox, C.W. Identification of Cytauxzoon felis Infection in Domestic Cats from Southern Illinois. J. Feline Med. Surg. 2015, 17, 1069–1072. [Google Scholar] [CrossRef]
- Reichard, M.V.; Baum, K.A.; Cadenhead, S.C.; Snider, T.A. Temporal Occurrence and Environmental Risk Factors Associated with Cytauxzoonosis in Domestic Cats. Vet. Parasitol. 2008, 152, 314–320. [Google Scholar] [CrossRef]
- Birkenheuer, A.J.; Le, J.A.; Valenzisi, A.M.; Tucker, M.D.; Levy, M.G.; Breitschwerdt, E.B. Cytauxzoon felis Infection in Cats in the Mid-Atlantic States: 34 Cases (1998–2004). J. Am. Vet. Med. Assoc. 2006, 228, 568–571. [Google Scholar] [CrossRef]
- Kao, Y.; Peake, B.; Madden, R.; Cowan, S.R.; Scimeca, R.C.; Thomas, J.E.; Reichard, M.V.; Ramachandran, A.; Miller, C.A. A probe-based droplet digital polymerase chain reaction assay for early detection of feline acute cytauxzoonosis. J. Vet. Parasit. 2021, 292, 109413. [Google Scholar] [CrossRef] [PubMed]
- Frontera-Acevedo, K. Feline Immune Response To Infection With Cytauxzoon felis and the Role of CD18 in the Pathogenesis of Cytauxzoonosis. Ph.D. Thesis, University of Georgia, Athens, GA, USA, 2013. [Google Scholar]
- Frontera-Acevedo, K.; Sakamoto, K. Local Pulmonary Immune Responses in Domestic Cats Naturally Infected with Cytauxzoon felis. Vet. Immunol. Immunopathol. 2015, 163, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ridgway, M.D. Feline Cytauxzoonosis. Available online: https://vetmed.illinois.edu/wp-content/uploads/2015/08/22.-Feline-Cytauxzoonosis.pdf (accessed on 3 September 2018).
- Conner, B.J.; Hanel, R.M.; Brooks, M.B.; Cohn, L.A.; Birkenheuer, A.J. Coagulation Abnormalities in 5 Cats with Naturally Occurring Cytauxzoonosis. J. Vet. Emerg. Crit. Care 2015, 25, 538–545. [Google Scholar] [CrossRef]
- Clarke, L.L.; Rissi, D.R. Neuropathology of Natural Cytauxzoon felis Infection in Domestic Cats. Vet. Pathol. 2015, 52, 1167–1171. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.L.; Krimer, P.M.; Rissi, D.R. Glial Changes and Evidence for Apoptosis in the Brain of Cats Infected by Cytauxzoon felis. J. Comp. Pathol. 2017, 156, 147–151. [Google Scholar] [CrossRef]
- Cohn, L.A.; Shaw, D.; Shoemake, C.; Birkenheuer, A.J. Second Illness Due to Subsequent Cytauxzoon felis Infection in a Domestic Cat. J. Feline Med. Surg. Open Rep. 2020, 6, 1–5. [Google Scholar] [CrossRef]
- Lewis, K. Cytauxzoon felis: An Emerging Feline Pathogen and Potential Therapy; University of Missouri: Columbia, MO, USA, 2011. [Google Scholar]
- Sleznikow, C.R.; Granick, J.L.; Cohn, L.A.; Nafe, L.A.; Rendahl, A.; Burton, E.N. Evaluation of various sample sources for the cytologic diagnosis of Cytauxzoon felis. J. Vet. Int. Med. 2021, 36, 126–132. [Google Scholar] [CrossRef]
- Sykes, J.E. Feline Hemotropic Mycoplasmas. Vet. Clin. N. Am. Small Anim. Pract. 2010, 40, 1157–1170. [Google Scholar] [CrossRef]
- Penzhorn, B.L.; Oosthuizen, M.C. Babesia Species of Domestic Cats: Molecular Characterization Has Opened Pandora’s Box. Front. Vet. Sci. 2020, 7, 134. [Google Scholar] [CrossRef]
- Motzel, S.L.; Wagner, J.E. Treatment of Experimentally Induced Cytauxzoonosis in Cats with Parvaquone and Buparvaquone. Vet. Parasitol. 1990, 35, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.M.; Modaresi, S.M.; Cook, J.L.; Latimer, K.S.; Peterson, D.S. Genetic Variability of Archived Cytauxzoon felis Histologic Specimens from Domestic Cats in Georgia, 1995-2007. J. Vet. Diagn. Investig. 2009, 21, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.M.; Cohn, L.A.; Marr, H.S.; Birkenheuer, A.J. Failure of Efficacy and Adverse Events Associated with Dose-Intense Diminazene Diaceturate Treatment of Chronic Cytauxzoon felis Infection in Five Cats. J. Feline Med. Surg. 2014, 16, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Cohn, L.A.; Birkenheuer, A.J.; Brunker, J.D.; Ratcliff, E.R.; Craig, A.W. Efficacy of Atovaquone and Azithromycin or Imidocarb Dipropionate in Cats with Acute Cytauxzoonosis. J. Vet. Intern. Med. 2011, 25, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, A.B.; Mather, M.W. Mitochondrial Evolution and Functions in Malaria Parasites. Annu. Rev. Microbiol. 2009, 63, 249–267. [Google Scholar] [CrossRef] [PubMed]
- Schreeg, M.E.; Marr, H.S.; Tarigo, J.; Cohn, L.A.; Levy, M.G.; Birkenheuer, A.J. Pharmacogenomics of Cytauxzoon felis Cytochrome b: Implications for Atovaquone and Azithromycin Therapy in Domestic Cats with Cytauxzoonosis. J. Clin. Microbiol. 2013, 51, 3066–3069. [Google Scholar] [CrossRef]
- Schreeg, M.E.; Marr, H.S.; Tarigo, J.L.; Cohn, L.A.; Levy, M.G.; Birkenheuer, A.J. Rapid High-Resolution Melt Analysis of Cytauxzoon felis Cytochrome b to Aid in the Prognosis of Cytauxzoonosis. J. Clin. Microbiol. 2015, 53, 2517–2524. [Google Scholar] [CrossRef]
- Srivastava, I.K.; Rottenberg, H.; Vaidya, A.B. Atovaquone, a Broad Spectrum Antiparasitic Drug, Collapses Mitochondrial Membrane Potential in a Malarial Parasite. J. Biol. Chem. 1997, 272, 3961–3966. [Google Scholar] [CrossRef]
- Hikosaka, K.; Komatsuya, K.; Suzuki, S.; Kira, K. Mitochondria of Malaria Parasites as a Drug Target. In An Overview of Tropical Diseases; Intech Open: London, UK, 2015; pp. 17–38. [Google Scholar] [CrossRef]
- Tarigo, J. The Cytauxzoon felis Genome: A Guide to Vaccine Candidate Antigen Discovery for Cytauxzoonosis. Ph.D. Thesis, North Carolina State University, Raleigh, NC, USA, 2013. [Google Scholar]
- Pollard, D.A.; Reichard, M.V.; Cohn, L.A.; James, A.M.; Holman, P.J. Genetic Variability of Cloned Cytauxzoon felis Ribosomal RNA ITS1 and ITS2 Genomic Regions from Domestic Cats with Varied Clinical Outcomes from Five States. Vet. Parasitol. 2017, 244, 136–143. [Google Scholar] [CrossRef]
- Tarigo, J.L.; Scholl, E.H.; Bird, D.M.K.; Brown, C.C.; Cohn, L.A.; Dean, G.A.; Levy, M.G.; Doolan, D.L.; Trieu, A.; Nordone, S.K.; et al. A Novel Candidate Vaccine for Cytauxzoonosis Inferred from Comparative Apicomplexan Genomics. PLoS ONE 2013, 8, e71233. [Google Scholar] [CrossRef]
- Tarigo, J.L.; Kelly, L.S.; Brown, H.M.; Peterson, D.S. Limited Genetic Variability of Cytauxzoon felis Apical Membrane Antigen-1 (Ama1) from Domestic Cats and Bobcats. Parasites Vectors 2019, 12, 115. [Google Scholar] [CrossRef] [PubMed]
- Reichard, M.V.; Thomas, J.E.; Arther, R.G.; Hostetler, J.A.; Raetzel, K.L.; Meinkoth, J.H.; Little, S.E. Efficacy of an Imidacloprid 10%/Flumethrin 4.5% Collar (Seresto®, Bayer) for Preventing the Transmission of Cytauxzoon felis to Domestic Cats by Amblyomma americanum. Parasitol. Res. 2013, 112, S11–S20. [Google Scholar] [CrossRef] [PubMed]
- Reichard, M.V.; Rugg, J.J.; Thomas, J.E.; Allen, K.E.; Barrett, A.W.; Murray, J.K.; Herrin, B.H.; Beam, R.A.; King, V.L.; Vatta, A.F. Efficacy of a Topical Formulation of Selamectin plus Sarolaner against Induced Infestations of Amblyomma americanum on Cats and Prevention of Cytauxzoon felis Transmission. Vet. Parasitol. 2019, 270, S31–S37. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wikander, Y.M.; Reif, K.E. Cytauxzoon felis: An Overview. Pathogens 2023, 12, 133. https://doi.org/10.3390/pathogens12010133
Wikander YM, Reif KE. Cytauxzoon felis: An Overview. Pathogens. 2023; 12(1):133. https://doi.org/10.3390/pathogens12010133
Chicago/Turabian StyleWikander, Yvonne M., and Kathryn E. Reif. 2023. "Cytauxzoon felis: An Overview" Pathogens 12, no. 1: 133. https://doi.org/10.3390/pathogens12010133
APA StyleWikander, Y. M., & Reif, K. E. (2023). Cytauxzoon felis: An Overview. Pathogens, 12(1), 133. https://doi.org/10.3390/pathogens12010133