
Stable Episomal Transfectant Leishmania infantum Promastigotes Over-Expressing the DEVH1 RNA Helicase Gene Down-Regulate Parasite Survival Genes

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasite Transfection
2.2. RNA Isolation, mRNA Amplification, and Indirect Labeling of cDNA with CYANINES
2.3. Microarray Hybridization Analysis
2.4. Identification of Differentially Regulated Genes
2.5. Validation by Real-Time Quantitative RT-PCR (qRT-PCR)
3. Results and Discussion
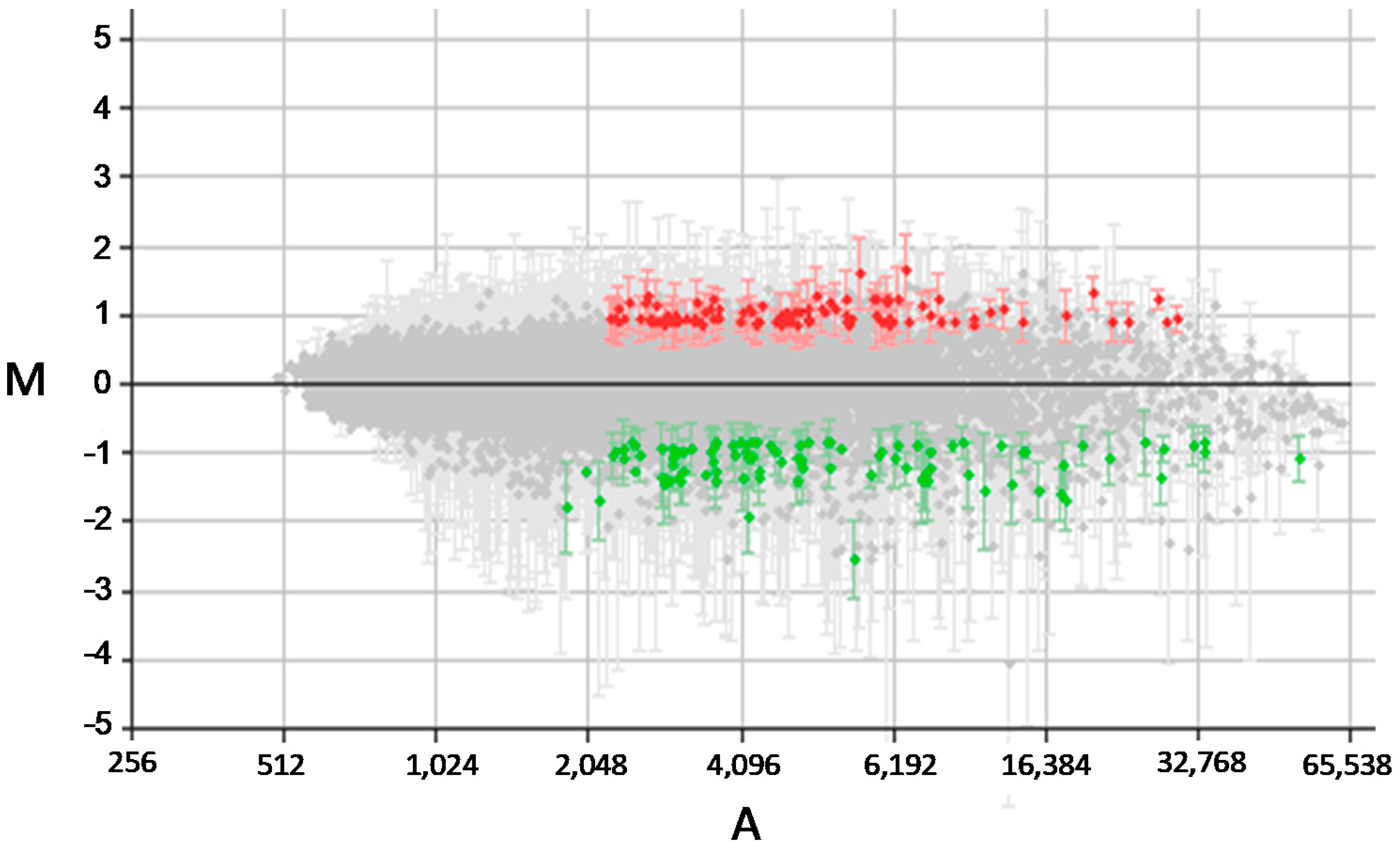
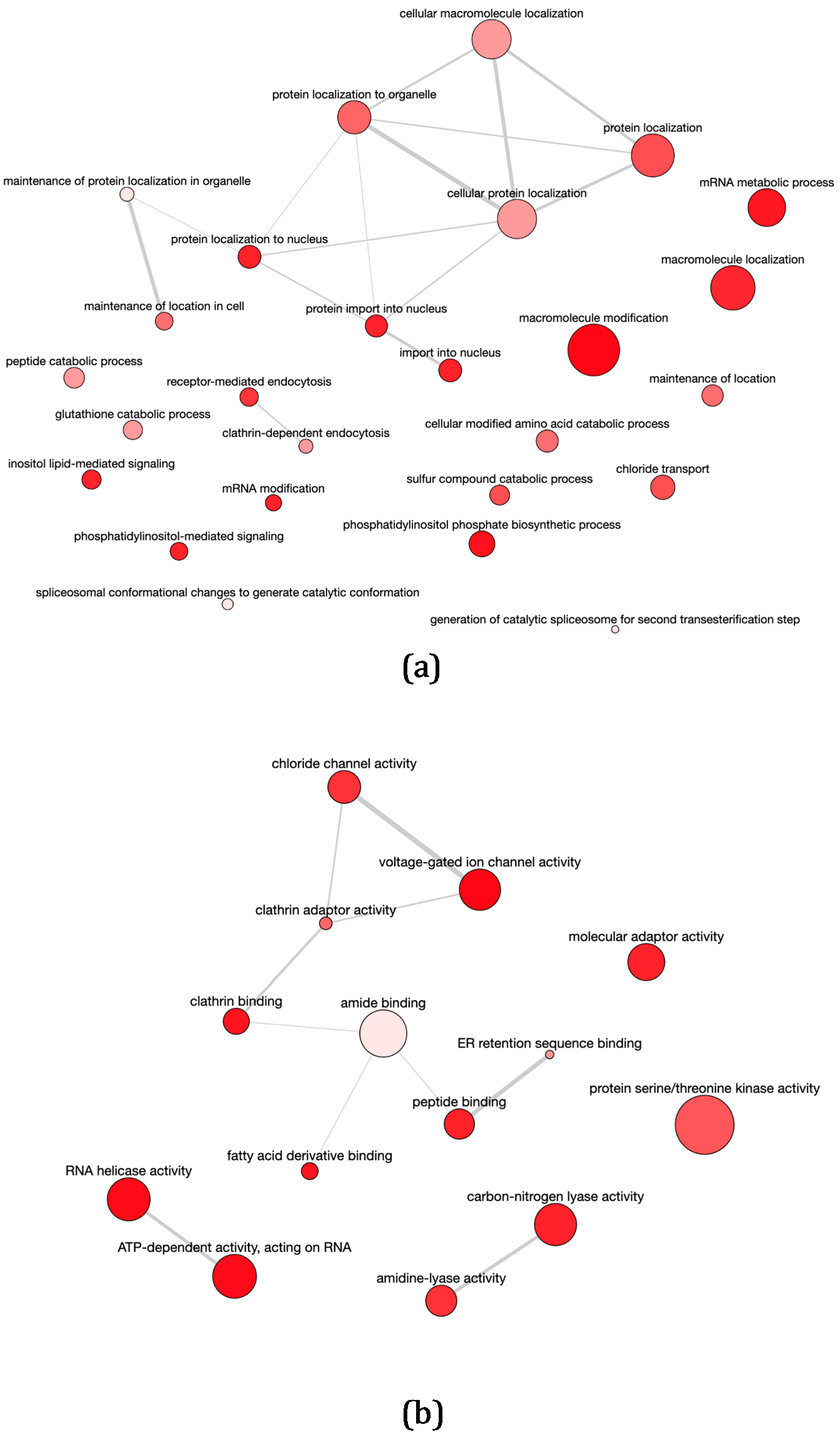
3.1. High-Throughput Differential Gene Expression Profiling of Stable Transfectant L. infantum Promastigotes Over-Expressing the DEVH1 Helicase Gene
3.2. Differential Transcript Abundance of Genes Involved in Gene Expression Regulation, Intracellular Signaling, Metabolism, Transport, and Movement in pTEX-DEVH1 L. infantum Promastigotes
3.3. pTEX-DEVH1 L. infantum Promastigotes Down-Regulate Genes Involved in Parasite Survival
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Report of a Meeting of the WHO Expert Committee on the Control of Leishmaniases; WHO: Geneva, Switzerland, 2010. [Google Scholar]
- Alvar, J.; Velez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; den Boer, M. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE 2012, 7, e35671. [Google Scholar]
- Gonzalez, E.; Jiménez, M.; Hernández, S.; Martín-Martín, I.; Molina, R. Phlebotomine sand fly survey in the focus of leishmaniasis in Madrid, Spain (2012–2014): Seasonal dynamics, Leishmania infantum infection rates and blood meal preferences. Parasit. Vectors 2017, 10, 368. [Google Scholar] [CrossRef]
- Gonzalez, E.; Jimenez, M.; Hernandez, S.; Martin-Martin, I.; Molina, R. Opportunistic feeding behaviour and Leishmania infantum detection in Phlebotomus perniciosus females collected in the human leishmaniasis focus of Madrid, Spain (2012–2018). PLoS Negl. Trop. Dis. 2021, 15, e0009240. [Google Scholar] [CrossRef]
- Molina, R.; Jiménez, M.I.; Cruz, I.; Iriso, A.; Martín-Martín, I.; Sevillano, O.; Melero, S.; Bernal, J. The hare (Lepus granatensis) as potential sylvatic reservoir of Leishmania infantum in Spain. Vet. Parasitol. 2012, 190, 268–271. [Google Scholar] [CrossRef]
- Bates, P.A. Transmission of Leishmania metacyclic promastigotes by phlebotomine sand flies. Int. J. Parasitol. 2007, 37, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Burchmore, R.J.; Barrett, M.P. Life in vacuoles—Nutrient acquisition by Leishmania amastigotes. Int. J. Parasitol. 2001, 31, 1311–1320. [Google Scholar] [CrossRef]
- Handman, E. Cell biology of Leishmania. Adv. Parasitol. 1999, 44, 1–39. [Google Scholar]
- Handman, E.; Bullen, D.V. Interaction of Leishmania with the host macrophage. Trends Parasitol. 2002, 18, 332–334. [Google Scholar] [CrossRef]
- Ivens, A.C.; Peacock, C.S.; Worthey, E.A.; Murphy, L.; Aggarwal, G.; Berriman, M.; Sisk, E.; Rajandream, M.A.; Adlem, E.; Aert, R.; et al. The genome of the kinetoplastid parasite, Leishmania major. Science 2005, 309, 436–442. [Google Scholar] [CrossRef] [Green Version]
- Myler, P.; Fasel, N. Leishmania—After the Genome; Caister Academic Press: Norfolk, UK, 2008. [Google Scholar]
- Clayton, C.; Shapira, M. Post-transcriptional regulation of gene expression in trypanosomes and leishmanias. Mol. Biochem. Parasitol. 2007, 156, 93–101. [Google Scholar] [CrossRef]
- Rogers, M.B.; Hilley, J.D.; Dickens, N.J.; Wilkes, J.; Bates, P.A.; Depledge, D.P.; Harris, D.; Her, Y.; Herzyk, P.; Imamura, H.; et al. Chromosome and gene copy number variation allow major structural change between species and strains of Leishmania. Genome Res. 2011, 21, 2129–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holzer, T.R.; McMaster, W.R.; Forney, J.D. Expression profiling by whole-genome interspecies microarray hybridization reveals differential gene expression in procyclic promastigotes, lesion-derived amastigotes, and axenic amastigotes in Leishmania mexicana. Mol. Biochem. Parasitol. 2006, 146, 198–218. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.; Lahav, T.; Holland, N.; Aggarwal, G.; Anupama, A.; Huang, Y.; Volpin, H.; Myler, P.J.; Zilberstein, D. Analysis of the Leishmania donovani transcriptome reveals an ordered progression of transient and permanent changes in gene expression during differentiation. Mol. Biochem. Parasitol. 2007, 152, 53–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcolea, P.J.; Alonso, A.; Gomez, M.J.; Sanchez-Gorostiaga, A.; Moreno-Paz, M.; Gonzalez-Pastor, E.; Torano, A.; Parro, V.; Larraga, V. Temperature increase prevails over acidification in gene expression modulation of amastigote differentiation in Leishmania infantum. BMC Genomics 2010, 11, 31. [Google Scholar] [CrossRef] [Green Version]
- Lahav, T.; Sivam, D.; Volpin, H.; Ronen, M.; Tsigankov, P.; Green, A.; Holland, N.; Kuzyk, M.; Borchers, C.; Zilberstein, D.; et al. Multiple levels of gene regulation mediate differentiation of the intracellular pathogen Leishmania. FASEB J. 2011, 25, 515–525. [Google Scholar] [CrossRef]
- Alcolea, P.J.; Alonso, A.; Molina, R.; Jimenez, M.; Myler, P.J.; Larraga, V. Functional genomics in sand fly-derived Leishmania promastigotes. PLoS Negl. Trop. Dis. 2019, 13, e0007288. [Google Scholar] [CrossRef] [Green Version]
- Alcolea, P.J.; Alonso, A.; Moreno-Izquierdo, M.A.; Degayon, M.A.; Moreno, I.; Larraga, V. Serum Removal from Culture Induces Growth Arrest, Ploidy Alteration, Decrease in Infectivity and Differential Expression of Crucial Genes in Leishmania infantum Promastigotes. PLoS ONE 2016, 11, e0150172. [Google Scholar]
- Rochette, A.; Raymond, F.; Corbeil, J.; Ouellette, M.; Papadopoulou, B. Whole-genome comparative RNA expression profiling of axenic and intracellular amastigote forms of Leishmania infantum. Mol. Biochem. Parasitol. 2009, 165, 32–47. [Google Scholar] [CrossRef]
- Ubeda, J.M.; Legare, D.; Raymond, F.; Ouameur, A.A.; Boisvert, S.; Rigault, P.; Corbeil, J.; Tremblay, M.J.; Olivier, M.; Papadopoulou, B.; et al. Modulation of gene expression in drug resistant Leishmania is associated with gene amplification, gene deletion and chromosome aneuploidy. Genome Biol. 2008, 9, R115. [Google Scholar] [CrossRef] [Green Version]
- Rastrojo, A.; Carrasco-Ramiro, F.; Martin, D.; Crespillo, A.; Reguera, R.M.; Aguado, B.; Requena, J.M. The transcriptome of Leishmania major in the axenic promastigote stage: Transcript annotation and relative expression levels by RNA-seq. BMC Genom. 2013, 14, 223. [Google Scholar] [CrossRef] [Green Version]
- Rastrojo, A.; Corvo, L.; Lombrana, R.; Solana, J.C.; Aguado, B.; Requena, J.M. Analysis by RNA-seq of transcriptomic changes elicited by heat shock in Leishmania major. Sci. Rep. 2019, 9, 6919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcolea, P.J.; Alonso, A.; Gomez, M.J.; Moreno, I.; Dominguez, M.; Parro, V.; Larraga, V. Transcriptomics throughout the life cycle of Leishmania infantum: High down-regulation rate in the amastigote stage. Int. J. Parasitol. 2010, 40, 1497–1516. [Google Scholar] [CrossRef] [PubMed]
- Kedersha, N.; Anderson, P. Mammalian stress granules and processing bodies. Methods Enzymol. 2007, 431, 61–81. [Google Scholar] [PubMed]
- Cassola, A.; de Gaudenzi, J.G.; Frasch, A.C. Recruitment of mRNAs to cytoplasmic ribonucleoprotein granules in trypanosomes. Mol. Microbiol. 2007, 65, 655–670. [Google Scholar] [CrossRef] [PubMed]
- Cassola, A. RNA Granules Living a Post-transcriptional Life: The Trypanosomes’ Case. Curr. Chem. Biol. 2011, 5, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Zinoviev, A.; Manor, S.; Shapira, M. Nutritional stress affects an atypical cap-binding protein in Leishmania. RNA Biol. 2012, 9, 1450–1460. [Google Scholar] [CrossRef] [Green Version]
- Pastro, L.; Smircich, P.; Di Paolo, A.; Becco, L.; Duhagon, M.A.; Sotelo-Silveira, J.; Garat, B. Nuclear Compartmentalization Contributes to Stage-Specific Gene Expression Control in Trypanosoma cruzi. Front. Cell Dev. Biol. 2017, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Kramer, S. RNA in development: How ribonucleoprotein granules regulate the life cycles of pathogenic protozoa. Wiley Interdiscip. Rev. RNA 2014, 5, 263–284. [Google Scholar] [CrossRef]
- Hooper, C.; Hilliker, A. Packing them up and dusting them off: RNA helicases and mRNA storage. Biochim. Biophys. Acta 2013, 1829, 824–834. [Google Scholar] [CrossRef]
- Linder, P. Dead-box proteins: A family affair—Active and passive players in RNP-remodeling. Nucleic Acids Res. 2006, 34, 4168–4180. [Google Scholar] [CrossRef]
- Gorbalenya, A.E.; Koonin, E.V.; Donchenko, A.P.; Blinov, V.M. Two related superfamilies of putative helicases involved in replication, recombination, repair and expression of DNA and RNA genomes. Nucleic Acids Res. 1989, 17, 4713–4730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilliker, A. Analysis of RNA helicases in P-bodies and stress granules. Methods Enzymol. 2012, 511, 323–346. [Google Scholar] [PubMed]
- Holetz, F.B.; Correa, A.; Rodrigues Ávila, A.; Nakamura, C.V.; Krieger, M.A.; Goldenberg, S. Evidence of P-body-like structures in Trypanosoma cruzi. Biochem. Biophys. Res. Commun. 2007, 356, 1062–1067. [Google Scholar] [CrossRef] [PubMed]
- Afonso-Lehmann, R.N.; Thomas, M.C.; Santana-Morales, M.A.; Deniz, D.; Lopez, M.C.; Valladares, B.; Martinez-Carretero, E. A DEVH-box RNA Helicase from Leishmania braziliensis is Associated to mRNA Cytoplasmic Granules. Protist. 2015, 166, 457–467. [Google Scholar] [CrossRef]
- Kelly, J.M.; Ward, H.M.; Miles, M.A.; Kendall, G. A shuttle vector which facilitates the expression of transfected genes DEVH-box RNA Helicase from Leismania braziliensis in Trypanosoma cruzi and Leishmania. Nucleic Acids Res. 1992, 20, 3963–3969. [Google Scholar] [CrossRef] [Green Version]
- Alcolea, P.J.; Alonso, A.; Sanchez-Gorostiaga, A.; Moreno-Paz, M.; Gomez, M.J.; Ramos, I.; Parro, V.; Larraga, V. Genome-wide analysis reveals increased levels of transcripts related with infectivity in peanut lectin non-agglutinated promastigotes of Leishmania infantum. Genomics 2009, 93, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Aslett, M.; Aurrecoechea, C.; Berriman, M.; Brestelli, J.; Brunk, B.P.; Carrington, M.; Depledge, D.P.; Fischer, S.; Gajria, B.; Gao, X.; et al. TriTrypDB: A functional genomic resource for the Trypanosomatidae. Nucleic Acids Res. 2010, 38, D457–D462. [Google Scholar] [CrossRef] [Green Version]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [Green Version]
- Twyffels, L.; Gueydan, C.; Kruys, V. Transportin-1 and Transportin-2: Protein nuclear import and beyond. FEBS Lett. 2014, 588, 1857–1868. [Google Scholar] [CrossRef]
- McConville, M.J.; Mullin, K.A.; Ilgoutz, S.C.; Teasdale, R.D. Secretory pathway of trypanosomatid parasites. Microbiol. Mol. Biol. Rev. 2002, 66, 122–154. [Google Scholar] [CrossRef] [Green Version]
- Bogacz, M.; Krauth-Siegel, R.L. Tryparedoxin peroxidase-deficiency commits trypanosomes to ferroptosis-type cell death. Elife 2018, 7, e37503. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, M.; Moreno, I.; Aizpurua, C.; Torano, A. Early mechanisms of Leishmania infection in human blood. Microbes Infect. 2003, 5, 507–513. [Google Scholar] [CrossRef]
- Dominguez, M.; Moreno, I.; Lopez-Trascasa, M.; Torano, A. Complement interaction with trypanosomatid promastigotes in normal human serum. J. Exp. Med. 2002, 195, 451–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depledge, D.P.; Evans, K.J.; Ivens, A.C.; Aziz, N.; Maroof, A.; Kaye, P.M.; Smith, D.F. Comparative expression profiling of Leishmania: Modulation in gene expression between species and in different host genetic backgrounds. PLoS Negl. Trop. Dis. 2009, 3, e476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depledge, D.P.; MacLean, L.M.; Hodgkinson, M.R.; Smith, B.A.; Jackson, A.P.; Ma, S.; Uliana, S.R.; Smith, D.F. Leishmania-specific surface antigens show sub-genus sequence variation and immune recognition. PLoS Negl. Trop. Dis. 2010, 4, e829. [Google Scholar] [CrossRef] [Green Version]
- McKean, P.G.; Denny, P.W.; Knuepfer, E.; Keen, J.K.; Smith, D.F. Phenotypic changes associated with deletion and overexpression of a stage-regulated gene family in Leishmania. Cell Microbiol. 2001, 3, 511–523. [Google Scholar] [CrossRef]
- Zhang, R.G.; Zhang, J.; Jing, B.Q. Virulence-associated gene profiling of different Leishmania spp. Zhongguo Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi 2009, 27, 307–311. [Google Scholar]
- Kumar, M.; Ranjan, K.; Singh, V.; Pathak, C.; Pappachan, A.; Singh, D.D. Hydrophilic Acylated Surface Protein A (HASPA) of Leishmania donovani: Expression, Purification and Biophysico-Chemical Characterization. Protein J. 2017, 36, 343–351. [Google Scholar] [CrossRef]
- Mortazavidehkordi, N.; Fallah, A.; Abdollahi, A.; Kia, V.; Khanahmad, H.; Najafabadi, Z.G.; Hashemi, N.; Estiri, B.; Roudbari, Z.; Najafi, A.; et al. A lentiviral vaccine expressing KMP11-HASPB fusion protein increases immune response to Leishmania major in BALB/C. Parasitol. Res. 2018, 117, 2265–2273. [Google Scholar] [CrossRef]
- Alcolea, P.J.; Alonso, A.; Domínguez, M.; Parro, V.; Jiménez, M.; Molina, R.; Larraga, V. Influence of the microenvironment in the transcriptome of Leishmania infantum promastigotes: Sand fly versus culture. PLoS Negl. Trop. Dis. 2016, 10, e0004693. [Google Scholar] [CrossRef]
- Alonso, A.; Larraga, V.; Alcolea, P.J. The contribution of DNA microarray technology to gene expression profiling in Leishmania spp.: A retrospective view. Acta Trop. 2018, 187, 129–139. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F | log2R ± SD | p | Id. | Gene Annotation (TriTrypDB) |
|---|---|---|---|---|
| 4.70 | 2.3 ± 0.4 | 0.003 | LINF_220021200 | ATP-dependent DEAD/H RNA helicase, putative |
| 3.04 | 1.6 ± 0.5 | 0.035 | LINF_300022400 | Hypothetical protein, conserved |
| 2.46 | 1.3 ± 0.2 | 0.011 | LINF_260024900 | Hypothetical protein, conserved |
| 2.39 | 1.3 ± 0.4 | 0.038 | LINF_230022400 | Dynein heavy chain, putative |
| 2.38 | 1.2 ± 0.1 | 0.002 | LINF_360020600 | N-terminal region of chorein—a TM vesicle-mediated sorter, putative |
| 2.36 | 1.2 ± 0.2 | 0.008 | LINF_340018000 | Hypothetical protein, conserved |
| 2.34 | 1.2 ± 0.1 | 0.004 | LINF_140005100 | PI3-kinase family—ras-binding domain/Phosphoinositide 3-kinase C2/Phosphoinositide 3-kinase family—accessory domain (PIK domain)/Phosphatidylinositol 3- and 4-kinase—putative |
| 2.32 | 1.2 ± 0.1 | 0.004 | LINF_340021000 | N-terminal region of chorein—a TM vesicle-mediated sorter/Protein of unknown function (DUF1162)—putative |
| 2.32 | 1.2 ± 0.4 | 0.043 | LINF_320041200 | Chloride channel protein, putative |
| 2.31 | 1.2 ± 0.1 | 0.002 | LINF_250008300 | Hypothetical protein, conserved |
| 2.29 | 1.2 ± 0.1 | 0.003 | LINF_310035300 | 3’−5’ exonuclease, putative |
| 2.24 | 1.2 ± 0.3 | 0.049 | LINF_270010200 | Calpain-like cysteine peptidase, putative |
| 2.21 | 1.1 ± 0.2 | 0.004 | LINF_040017400 | Hypothetical protein, conserved |
| 2.18 | 1.1 ± 0.3 | 0.031 | LINF_110011000 | Hypothetical protein, conserved |
| 2.15 | 1.1 ± 0.0 | 0.000 | LINF_330008300 | Glucose transporter/membrane transporter D2, putative |
| 2.12 | 1.1 ± 0.3 | 0.027 | LINF_070005500 | Alpha-adaptin-like protein |
| 2.11 | 1.1 ± 0.3 | 0.019 | LINF_330014800 | NLI interacting factor-like phosphatase |
| 2.11 | 1.1 ± 0.4 | 0.035 | LINF_270028900 | WD domain-G-beta repeat, putative |
| 2.10 | 1.1 ± 0.3 | 0.027 | LINF_160013300 | N-terminal region of chorein-A TM vesicle-mediated sorter, putative |
| 2.09 | 1.1 ± 0.3 | 0.021 | LINF_280011400 | ER lumen retaining receptor-like protein |
| 2.06 | 1.0 ± 0.2 | 0.010 | LINF_360054800 | Related to elongation factor−2 kinase efk−1b isoform-like protein |
| 2.06 | 1.0 ± 0.0 | 0.001 | LINF_360035100 | Transportin2-like protein |
| 2.05 | 1.0 ± 0.3 | 0.025 | LINF_330029900 | Glycerol phosphate mutase, putative |
| 2.01 | 1.0 ± 0.2 | 0.020 | LINF_310041000 | Phosphoglycan beta−1,3-galactosyltransferase 4 |
| 2.00 | 1.0 ± 0.4 | 0.049 | LINF_360034600 | Hypothetical protein, conserved |
| 1.96 | 1.0 ± 0.3 | 0.037 | LINF_350031700 | Hypothetical protein, conserved |
| 1.96 | 1.0 ± 0.3 | 0.035 | LINF_360020400 | Zn-finger in Ran binding protein and others, putative |
| 1.95 | 1.0 ± 0.3 | 0.028 | LINF_150005100 | Hypothetical protein, conserved |
| 1.95 | 1.0 ± 0.3 | 0.032 | LINF_350014800 | Casein kinase, putative |
| 1.93 | 0.9 ± 0.1 | 0.007 | LINF_150015300 | Hypothetical protein, conserved |
| 1.93 | 0.9 ± 0.3 | 0.039 | LINF_270023400 | Hypothetical protein, conserved |
| 1.92 | 0.9 ± 0.3 | 0.033 | LINF_220017900 | ChaC-like protein, putative |
| 1.91 | 0.9 ± 0.2 | 0.019 | LINF_350052900 | Hsp70 protein, putative |
| 1.91 | 0.9 ± 0.3 | 0.025 | LINF_240005600 | Hypothetical protein, conserved |
| 1.90 | 0.9 ± 0.2 | 0.016 | LINF_120012700 | Hypothetical protein, conserved |
| 1.90 | 0.9 ± 0.2 | 0.013 | LINF_350047900 | Hypothetical protein, conserved |
| 1.88 | 0.9 ± 0.4 | 0.049 | LINF_060014200 | Hypothetical protein, conserved |
| 1.88 | 0.9 ± 0.3 | 0.043 | LINF_280018500 | DnaJ domain-containing protein, putative |
| 1.87 | 0.9 ± 0.1 | 0.020 | LINF_140020800 | Hypothetical protein, conserved |
| 1.87 | 0.9 ± 0.2 | 0.010 | LINF_350045000 | U5 snRNA-associated splicing factor |
| 1.86 | 0.9 ± 0.1 | 0.005 | LINF_290031800 | Acyltransferase, putative |
| 1.86 | 0.9 ± 0.1 | 0.007 | LINF_310008600 | Amino acid transporter aATP11, putative |
| 1.85 | 0.9 ± 0.3 | 0.029 | LINF_070005100 | Isy1-like splicing family—putative |
| 1.84 | 0.9 ± 0.3 | 0.032 | LINF_170012400 | Acyl-CoA-binding protein |
| 1.84 | 0.9 ± 0.3 | 0.039 | LINF_100018200 | Hypothetical protein, conserved |
| 1.84 | 0.9 ± 0.3 | 0.045 | LINF_360071900 | Hypothetical protein, conserved |
| 1.84 | 0.9 ± 0.3 | 0.045 | LINF_270014400 | Right-handed beta helix region/Periplasmic copper-binding protein (NosD), putative |
| 1.84 | 0.9 ± 0.0 | 0.000 | LINF_340042800 | LicD family, putative |
| 1.83 | 0.9 ± 0.3 | 0.033 | LINF_140013700 | Hypothetical protein, conserved |
| 1.83 | 0.9 ± 0.2 | 0.017 | LINF_170008500 | Kinesin motor domain-containing protein, putative |
| 1.81 | 0.9 ± 0.3 | 0.036 | LINF_230022400 | Dynein, heavy chain, putative |
| 1.80 | 0.8 ± 0.3 | 0.043 | LINF_210006700 | Serine/threonine protein kinase, putative |
| −1.80 | −0.8 ± 0.2 | 0.014 | LINF_080011900 | Amastin-like protein |
| −1.80 | −0.9 ± 0.3 | 0.049 | LINF_330021300 | Glutamine aminotransferase, putative |
| −1.81 | −0.9 ± 0.2 | 0.025 | LINF_310013600 | C2 domain protein, putative |
| −1.82 | −0.9 ± 0.2 | 0.014 | LINF_220013600/700 | NADH-cytochrome b5 reductase |
| −1.83 | −0.9 ± 0.5 | 0.044 | LINF_060005100 | Hypothetical protein, conserved |
| −1.83 | −0.9 ± 0.1 | 0.007 | LINF_190020900 | Mitogen-activated protein kinase 4, putative |
| −1.83 | −0.9 ± 0.3 | 0.029 | LINF_120007800 | Hypothetical protein, conserved |
| −1.83 | −0.9 ± 0.3 | 0.028 | LINF_120007800 | Hypothetical protein, conserved |
| −1.83 | −0.9 ± 0.2 | 0.019 | LINF_270022500 | Hypothetical protein, conserved |
| −1.84 | −0.9 ± 0.3 | 0.042 | LINF_290014500 | Phytanoyl-CoA dioxygenase (PhyH), putative |
| −1.85 | −0.9 ± 0.3 | 0.038 | LINF_310023200 | Hypothetical protein, conserved |
| −1.87 | −0.9 ± 0.1 | 0.004 | LINF_320040300 | Hypothetical protein, conserved |
| −1.88 | −0.9 ± 0.0 | 0.010 | LINF_190005300 | Histone H2B (H2B) |
| −1.92 | −0.9 ± 0.2 | 0.016 | LINF_230006200 | Concanavalin A-like lectin/glucanases superfamily/Beige/BEACH domain-containing protein, putative |
| −1.93 | −0.9 ± 0.4 | 0.033 | LINF_350043500 | Hypothetical protein, conserved |
| −1.94 | −1.0 ± 0.0 | 0.049 | LINF_200012700 | Tubulin/FtsZ family, putative |
| −1.95 | −1.0 ± 0.0 | 0.012 | LINF_260013100 | Type II (glutathione peroxidase-like) tryparedoxin peroxidase |
| −1.98 | −1.0 ± 0.0 | 0.049 | LINF_360061200/300 | Vacuolar sorting protein-associated protein-like protein/Aldehyde dehydrogenase, putative |
| −2.02 | −1.0 ± 0.2 | 0.014 | LINF_170008100 | Tetratricopeptide repeat—putative |
| −2.02 | −1.0 ± 0.4 | 0.049 | LINF_180018900 | Hypothetical protein, conserved |
| −2.05 | −1.0 ± 0.4 | 0.039 | LINF_170005200 | Hypothetical protein, conserved |
| −2.12 | −1.1 ± 0.4 | 0.049 | LINF_360045700 | Mitogen-activated protein kinase-like |
| −2.14 | −1.1 ± 0.3 | 0.031 | LINF_330016400 | Hypothetical protein, conserved |
| −2.16 | −1.1 ± 0.4 | 0.040 | LINF_310022900 | Hypothetical protein, conserved |
| −2.18 | −1.1 ± 0.2 | 0.015 | LINF_360037500 | GDP-forming succinyl-CoA ligase b chain, putative |
| −2.20 | −1.1 ± 0.3 | 0.022 | LINF_150012500 | Ecotin, putative |
| −2.28 | −1.2 ± 0.0 | 0.046 | LINF_160015600 | Protein of unknown function (DUF3184), putative |
| −2.33 | −1.2 ± 0.3 | 0.014 | LINF_270032600 | 3-oxoacyl-ACP reductase, putative (KAR1) |
| −2.34 | −1.2 ± 0.3 | 0.018 | LINF 320037400 | Hypothetical protein, conserved |
| −2.38 | −1.2 ± 0.5 | 0.048 | LINF_350007000 | NLI interacting factor-like phosphatase—putative |
| −2.40 | −1.2 ± 0.5 | 0.048 | LINF_090014800 | Hypothetical protein, conserved |
| −2.40 | −1.3 ± 0.4 | 0.026 | LINF_130007800 | Alpha tubulin |
| −2.44 | −1.2 ± 0.0 | 0.048 | LINF_300039100 | 60S ribosomal protein L9, putative |
| −2.52 | −1.3 ± 0.0 | 0.033 | LINF_120015800/900 | Putative integral membrane protein conserved region (DUF2404), putative |
| −2.53 | −1.3 ± 0.5 | 0.046 | LINF_360026000 | Inosine-guanosine transporter |
| −2.56 | −1.4 ± 0.5 | 0.036 | LINF_290036400 | 40S ribosomal protein S19-like protein |
| −2.57 | −1.4 ± 0.7 | 0.039 | LINF_050014100 | CPSF A subunit region-containing protein, putative |
| −2.59 | −1.4 ± 0.4 | 0.028 | LINF_290017500 | Tryparedoxin 1, putative |
| −2.65 | −1.4 ± 0.5 | 0.049 | LINF_280027900 | Cullin 2, putative |
| −2.67 | −1.4 ± 0.1 | 0.010 | LINF_230018600 | Hydrophilic surface protein A (HASPA1) |
| −2.69 | −1.4 ± 0.3 | 0.019 | LINF_040008500 | ADP ribosylation factor, putative |
| −2.72 | −1.4 ± 0.5 | 0.046 | LINF_360026300 | Phosphomannomutase, putative |
| −2.93 | −1.5 ± 0.4 | 0.023 | LINF_300028400 | Hypothetical protein, conserved |
| −3.82 | −1.9 ± 0.5 | 0.021 | LINF_230018700 | Hydrophilic surface protein B (HASPB) |
| −3.82 | −1.9 ± 0.5 | 0.021 | LINF_230018800 | Small hydrophilic endoplasmic-reticulum-associated protein (SHERP) |
| −3.82 | −1.9 ± 0.5 | 0.021 | LINF_230018900 | Hydrophilic surface protein A (HASPA2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonso, A.; Larraga, J.; Loayza, F.J.; Martínez, E.; Valladares, B.; Larraga, V.; Alcolea, P.J. Stable Episomal Transfectant Leishmania infantum Promastigotes Over-Expressing the DEVH1 RNA Helicase Gene Down-Regulate Parasite Survival Genes. Pathogens 2022, 11, 761. https://doi.org/10.3390/pathogens11070761
Alonso A, Larraga J, Loayza FJ, Martínez E, Valladares B, Larraga V, Alcolea PJ. Stable Episomal Transfectant Leishmania infantum Promastigotes Over-Expressing the DEVH1 RNA Helicase Gene Down-Regulate Parasite Survival Genes. Pathogens. 2022; 11(7):761. https://doi.org/10.3390/pathogens11070761
Chicago/Turabian StyleAlonso, Ana, Jaime Larraga, Francisco Javier Loayza, Enrique Martínez, Basilio Valladares, Vicente Larraga, and Pedro José Alcolea. 2022. "Stable Episomal Transfectant Leishmania infantum Promastigotes Over-Expressing the DEVH1 RNA Helicase Gene Down-Regulate Parasite Survival Genes" Pathogens 11, no. 7: 761. https://doi.org/10.3390/pathogens11070761
APA StyleAlonso, A., Larraga, J., Loayza, F. J., Martínez, E., Valladares, B., Larraga, V., & Alcolea, P. J. (2022). Stable Episomal Transfectant Leishmania infantum Promastigotes Over-Expressing the DEVH1 RNA Helicase Gene Down-Regulate Parasite Survival Genes. Pathogens, 11(7), 761. https://doi.org/10.3390/pathogens11070761