
Characterization of Nematicidal Activity and Nematode-Toxic Metabolites of a Soilborne Brevundimonas bullata Isolate


Abstract
:1. Introduction
2. Results
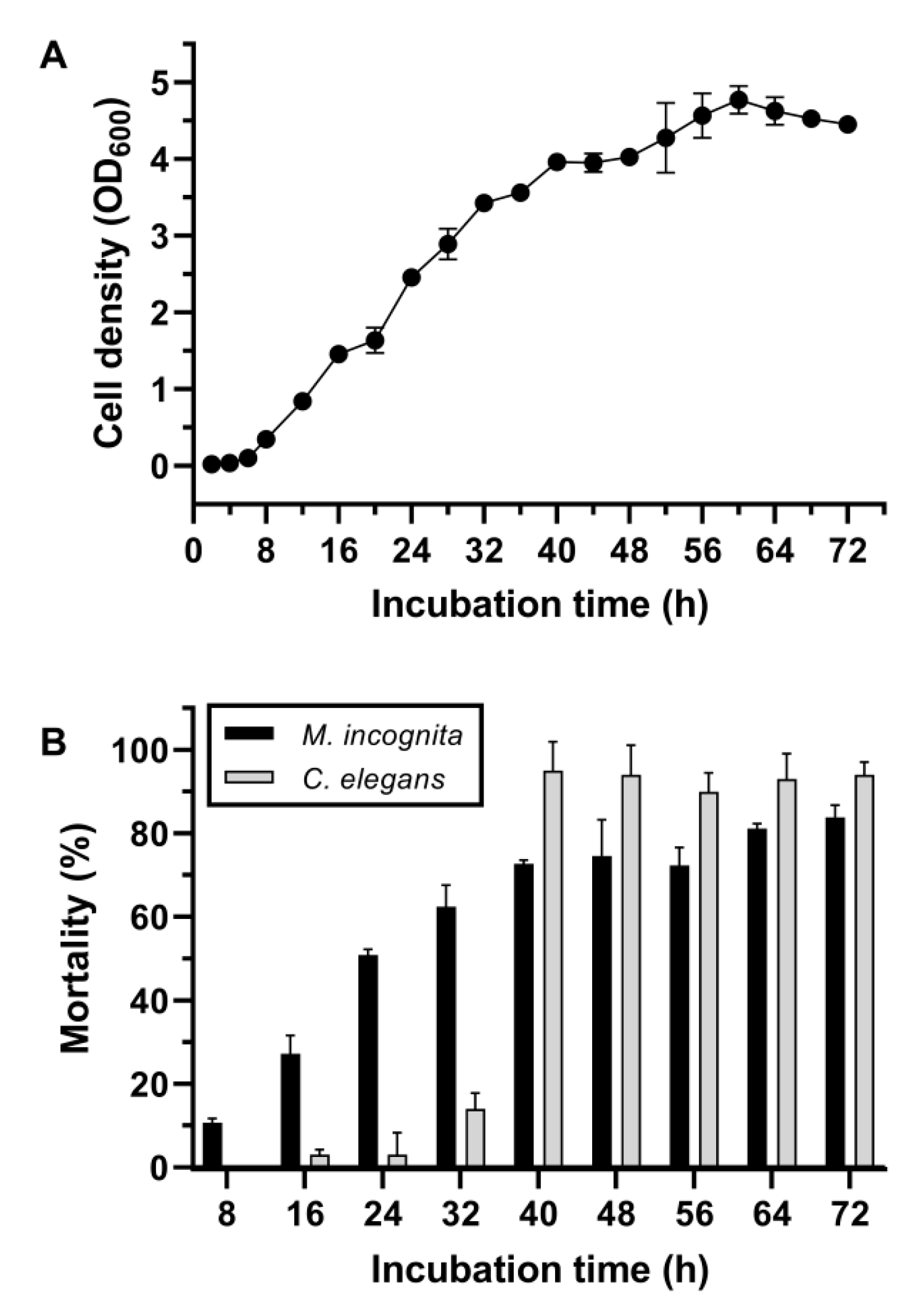
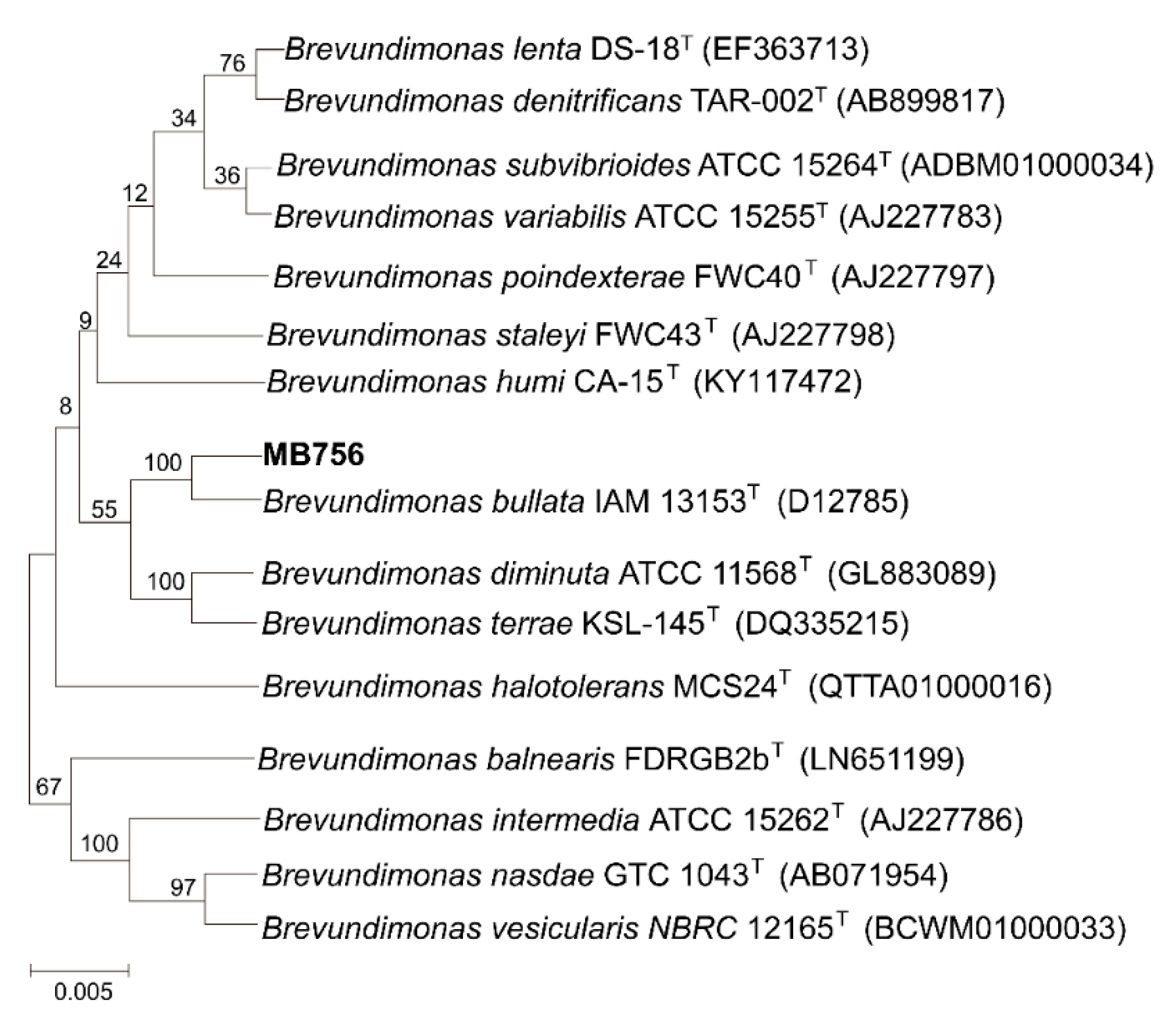
2.1. Screening and Identification of the Highly Nematicidal Brevundimonas Bullata MB756 and Its Nematicidal Efficacy
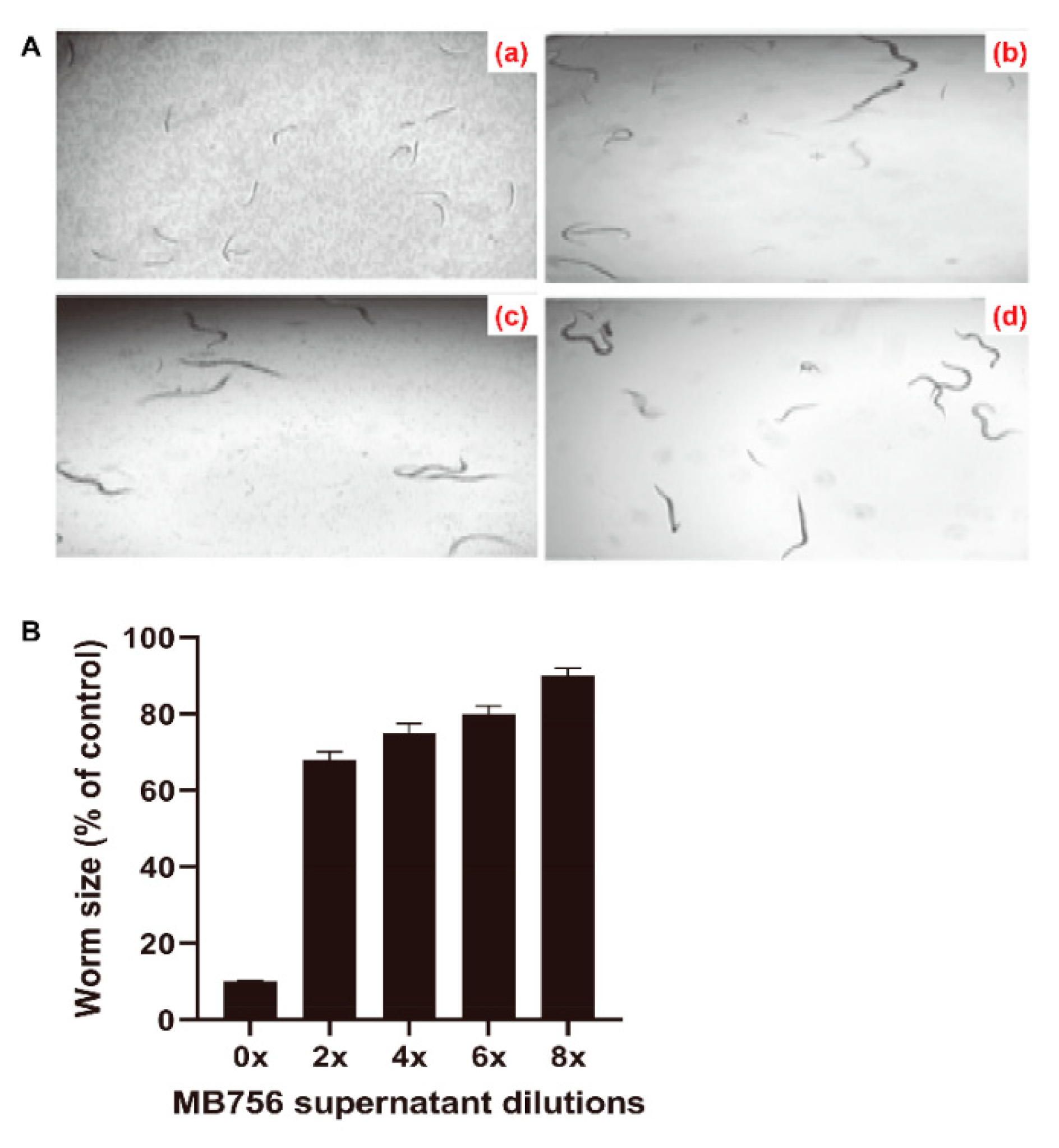
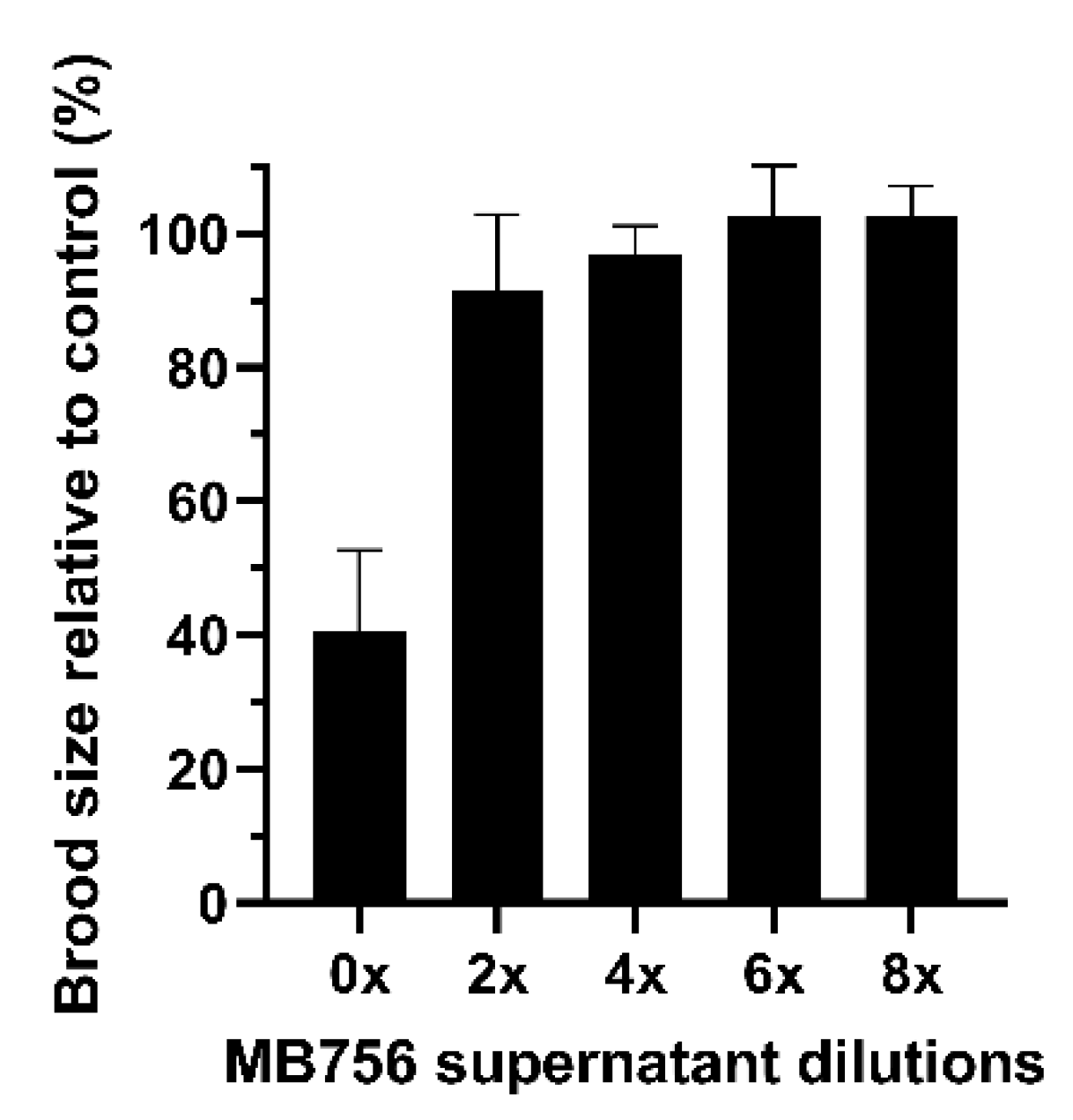
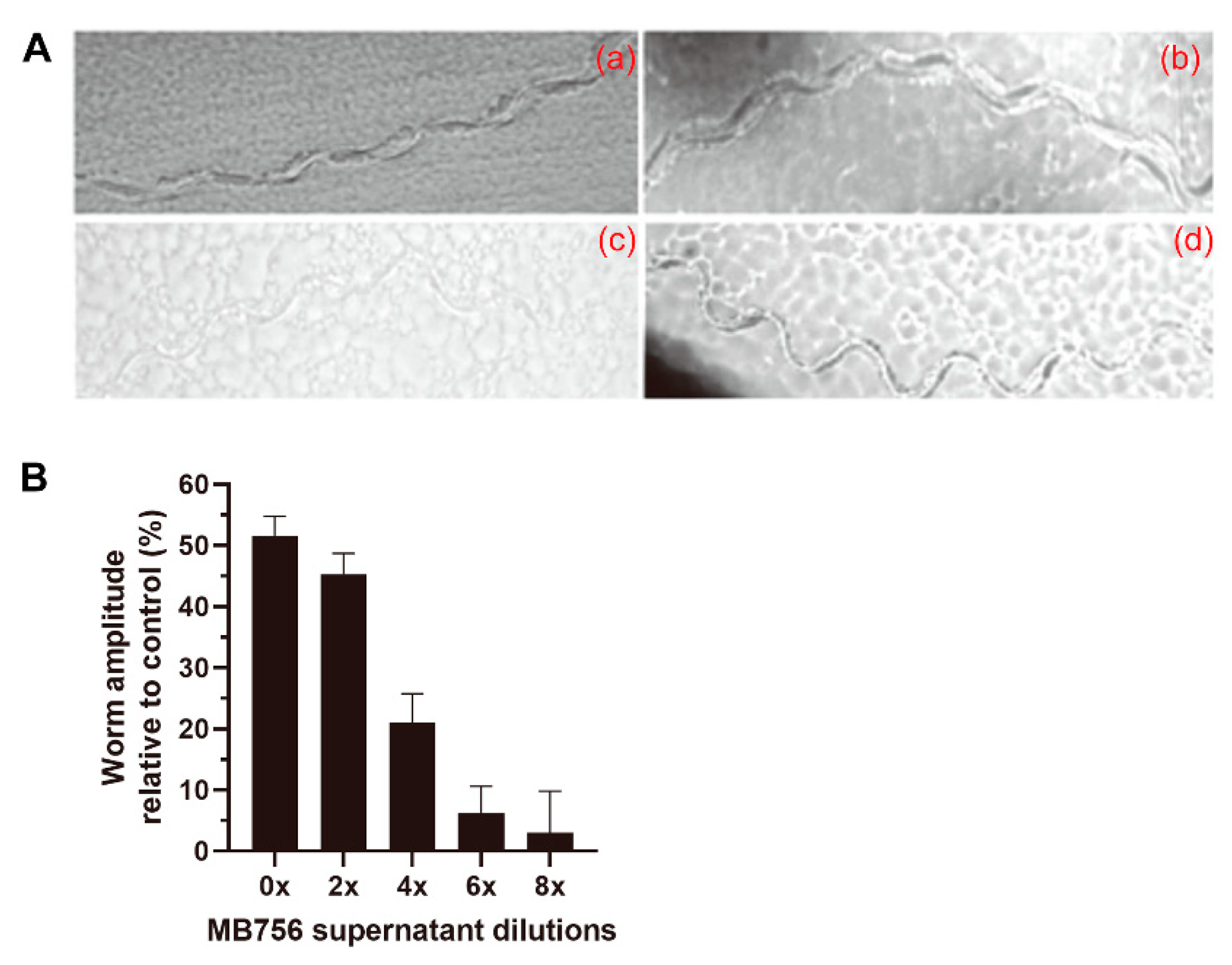
2.2. Effects of SCS of MB756 Cells on the Growth, Brood Size, and Mobility of C. elegans
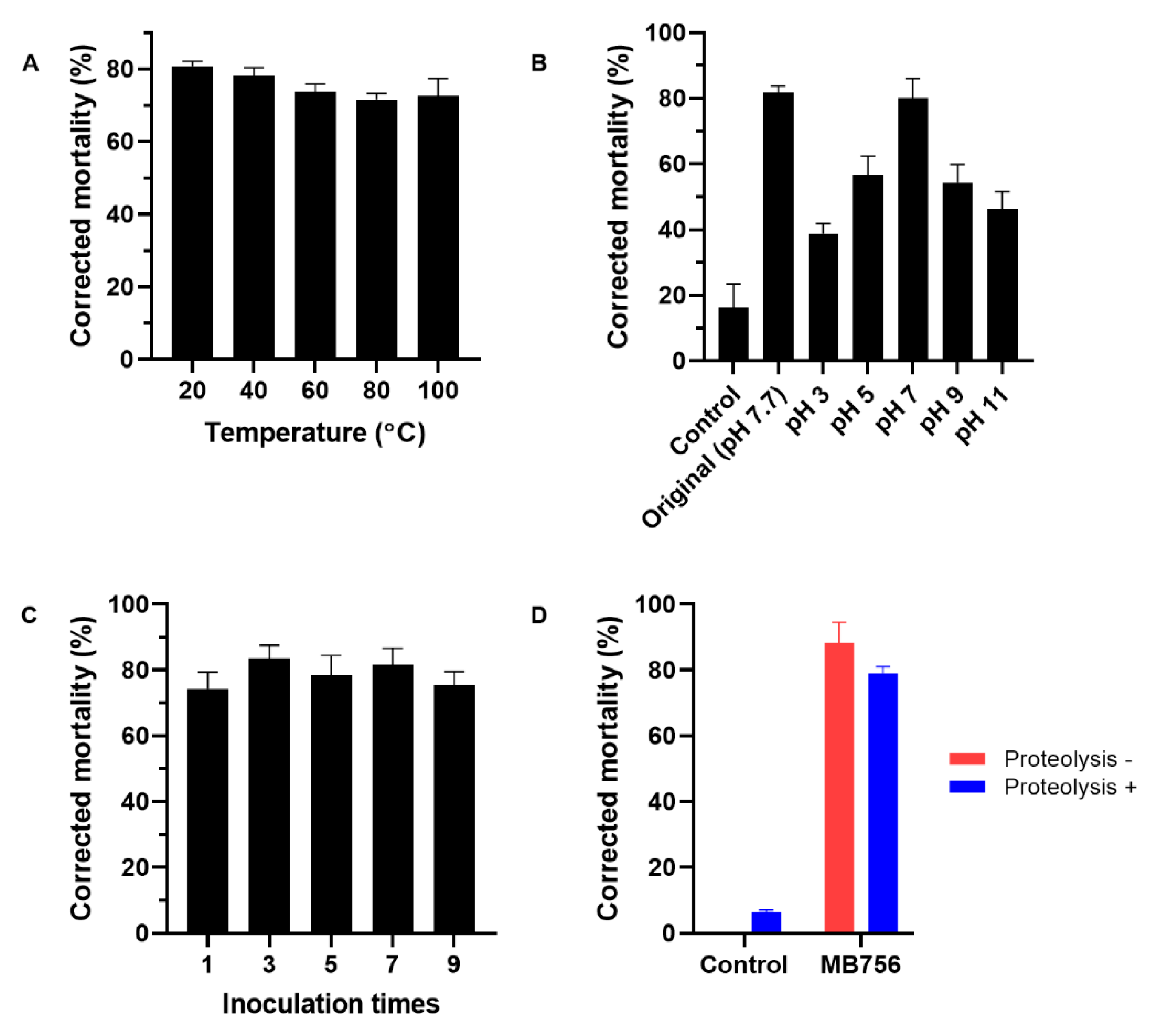
2.3. Effects of Different Temperatures, pH Values, Inoculation Times, and Proteolysis Hydrolysis Treatments on the Nematicidal Activity of the MB756 SCS against M. incognita
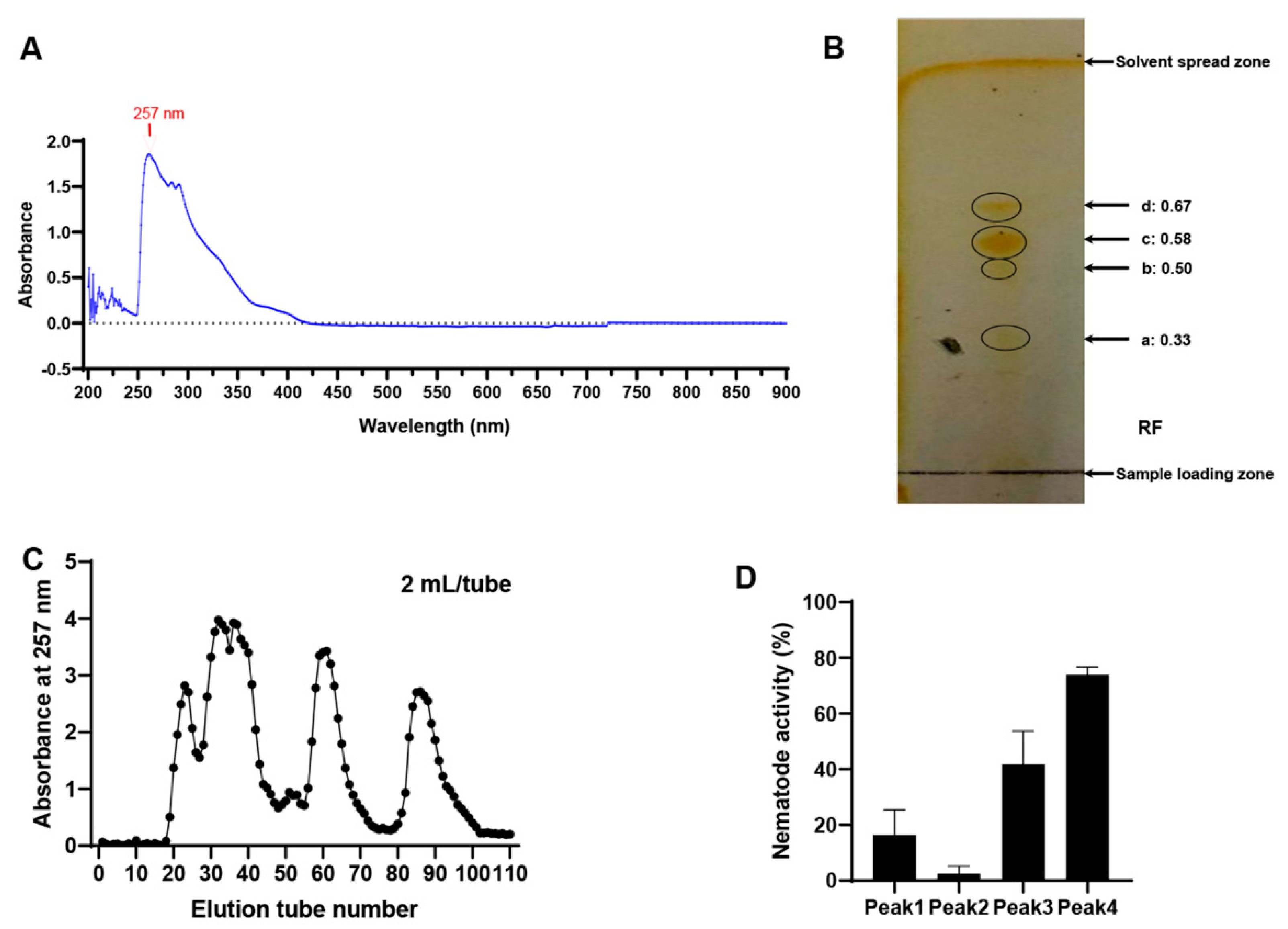
2.4. Purification and Preliminary Identification of the Active Substances in the MB756 SCS
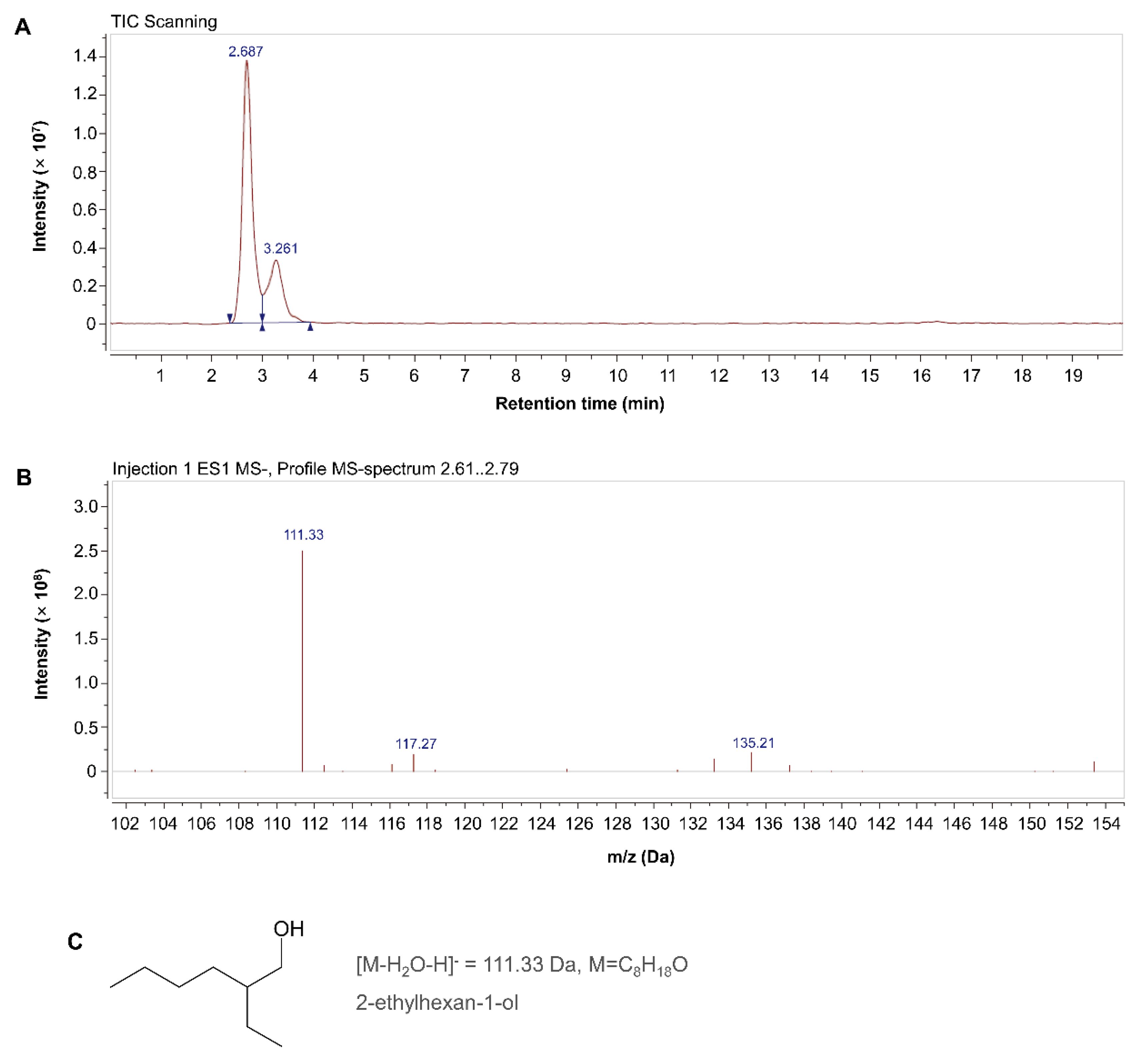
2.5. Preliminary Identification of the Main M. incognita-Toxic Substance in the MB756SCS
3. Discussion
4. Materials and Methods
4.1. Soil Sampling, Enrichment Culturing, and Isolation of Soilborne Bacteria
4.2. Bacterial Strains, Nematodes, Medium, and Culture Conditions
4.3. Bacterial Isolate Identification
4.4. Screening of Nematode-Lethal Bacterial Strains
4.5. Assays on the Growth, Brood Size, and Motility of C. elegans
4.6. Bioassays of the SCS of MB756 under Different Treatments
4.7. Organic Solvent Extraction of the SCS of MB756
4.8. Purification and Identification of M. incognita-Toxic Substances of the MB756 SCS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Abad, P.; Gouzy, J.; Aury, J.M.; Castagnone-Sereno, P.; Danchin, E.G.; Deleury, E.; Perfus-Barbeoch, L.; Anthouard, V.; Artiguenave, F.; Blok, V.C.; et al. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, D.; Handoo, Z. Taxonomy, identification and principal species. In Root-Knot Nematodes; Perry, M., Starr, J.L., Eds.; CABI Publishing: Wallingford, UK, 2009; pp. 55–97. [Google Scholar]
- Nicol, J.M.; Turner, S.J.; Coyne, D.; Den Nijs, L.; Hockland, S.; Maafi, Z.T. Current nematode threats to world agriculture. In Genomics and Molecular Genetics of Plant-Nematode Interactions; Jones, J., Gheysen, G., Fenoll, C., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 21–43. [Google Scholar]
- Jin, N.; Chen, Y.; Liu, Q.; Jian, H. Progress in occurrence, diagnoses, pathogenic mechanism and integrated management of root-knot nematode in China. J. Plant Prot. 2022, 49, 424–438. [Google Scholar] [CrossRef]
- Yang, J.; Liang, L.; Li, J.; Zhang, K.Q. Nematicidal enzymes from microorganisms and their applications. Appl. Microbiol. Biotechnol. 2013, 97, 7081–7095. [Google Scholar] [CrossRef]
- Faria, J.M.S.; Barbosa, P.; Vieira, P.; Vicente, C.S.L.; Figueiredo, A.C.; Mota, M. Phytochemicals as biopesticides against the pinewood nematode Bursaphelenchus xylophilus: A review on essential oils and their volatiles. Plants 2021, 10, 2614. [Google Scholar] [CrossRef] [PubMed]
- Desaeger, J.; Wram, C.; Zasada, I. New reduced-risk agricultural nematicides—Rationale and review. J. Nematol. 2020, 52, e2020–e2091. [Google Scholar] [CrossRef]
- Sikandar, A.; Zhang, M.Y.; Wang, Y.Y.; Zhu, X.F.; Lliu, X.Y.; Fan, H.Y.; Xuan, Y.H.; Chen, L.J.; Duan, Y.X. Root-knot nematode (Meloidogyne incognita) and its management: A review. Appl. Ecol. Environ. Res. 2020, 18, 1679–1690. [Google Scholar] [CrossRef]
- Engelbrecht, G.; Horak, I.; Jansen van Rensburg, P.J.; Claassens, S. Bacillus-based bionematicides: Development, modes of action and commercialisation. Biocontrol Sci. Technol. 2018, 28, 629–653. [Google Scholar] [CrossRef]
- Tian, B.; Yang, J.; Zhang, K.Q. Bacteria used in the biological control of plant-parasitic nematodes: Populations, mechanisms of action, and future prospects. FEMS Microbiol. Ecol. 2007, 61, 197–213. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Shao, Y.; Wan, J.; Feng, H.; Zhu, H.; Huang, H.; Zhou, Y. Isolation and characterization of a rhizobacterial antagonist of root-knot nematodes. PLoS ONE 2014, 9, e85988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, D.; Lin, J.; Huang, Q.; Zheng, W.; Liu, G.; Zheng, J.; Zhu, L.; Sun, M. A novel metalloproteinase virulence factor is involved in Bacillus thuringiensis pathogenesis in nematodes and insects. Environ. Microbiol. 2016, 18, 846–862. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Xiong, J.; Zhou, Q.; Xia, L.; Yu, Z. The effects of Bacillus thuringiensis Cry6A on the survival, growth, reproduction, locomotion, and behavioral response of Caenorhabditis elegans. Appl. Microbiol. Biotechnol. 2013, 97, 10135–10142. [Google Scholar] [CrossRef]
- Liu, Z.; Budiharjo, A.; Wang, P.; Shi, H.; Fang, J.; Borriss, R.; Zhang, K.; Huang, X. The highly modified microcin peptide plantazolicin is associated with nematicidal activity of Bacillus amyloliquefaciens FZB42. Appl. Microbiol. Biotechnol. 2013, 97, 10081–10090. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Qi, G.; Yin, R.; Zhang, H.; Li, C.; Zhao, X. Bacillus cereus strain S2 shows high nematicidal activity against Meloidogyne incognita by producing sphingosine. Sci. Rep. 2016, 6, 28756. [Google Scholar] [CrossRef]
- Geng, C.; Nie, X.; Tang, Z.; Zhang, Y.; Lin, J.; Sun, M.; Peng, D. A novel serine protease, Sep1, from Bacillus firmus DS-1 has nematicidal activity and degrades multiple intestinal-associated nematode proteins. Sci. Rep. 2016, 6, 25012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, J.; Zhou, Q.; Luo, H.; Xia, L.; Li, L.; Sun, M.; Yu, Z. Systemic nematicidal activity and biocontrol efficacy of Bacillus firmus against the root-knot nematode Meloidogyne incognita. World J. Microbiol. Biotechnol. 2015, 31, 661–667. [Google Scholar] [CrossRef]
- Terefe, M.; Tefera, T.; Sakhuja, P. Effect of a formulation of Bacillus firmus on root-knot nematode Meloidogyne incognita infestation and the growth of tomato plants in the greenhouse and nursery. J. Invertebr. Pathol. 2009, 100, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.; Zhang, L.; Zhang, K.; Huang, X.; Hui, F.; Kan, Y.; Yao, L. Changes in intestinal microflora of Caenorhabditis elegans following Bacillus nematocida B16 infection. Sci. Rep. 2016, 6, 20178. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Jiang, H.; Cheng, Q.; Chen, J.; Wu, G.; Kumar, A.; Sun, M.; Liu, Z. Enhanced nematicidal potential of the chitinase pachi from Pseudomonas aeruginosa in association with Cry21Aa. Sci. Rep. 2015, 5, 14395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feinbaum, R.L.; Urbach, J.M.; Liberati, N.T.; Djonovic, S.; Adonizio, A.; Carvunis, A.-R.; Ausubel, F.M. Genome-wide identification of Pseudomonas aeruginosa virulence-related genes using a Caenorhabditis elegans infection model. PLoS Pathog. 2012, 8, e1002813. [Google Scholar] [CrossRef] [Green Version]
- Dubern, J.F.; Cigana, C.; De Simone, M.; Lazenby, J.; Juhas, M.; Schwager, S.; Bianconi, I.; Döring, G.; Eberl, L.; Williams, P. Integrated whole-genome screening for Pseudomonas aeruginosa virulence genes using multiple disease models reveals that pathogenicity is host specific. Environ. Microbiol. 2015, 17, 4379–4393. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Jing, X.; Peng, W.L.; Nie, Q.; Zhai, Y.; Shao, Z.; Zheng, L.; Cai, M.; Li, G.; Zuo, H.; et al. Comparative genomic and functional analyses: Unearthing the diversity and specificity of nematicidal factors in Pseudomonas putida strain 1A00316. Sci. Rep. 2016, 6, 29211. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, R.; Ding, M.; Liu, Y.; Li, L. Biocontrol of the root-knot nematode Meloidogyne incognita by a nematicidal bacterium Pseudomonas simiae MB751 with cyclic dipeptide. Pest Manag. Sci. 2021, 77, 4365–4374. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Sun, Y.; Xie, L.; Yu, H.; Bashir, A.; Li, L. The pathogenicity of Pseudomonas syringae MB03 against Caenorhabditis elegans and the transcriptional response of nematicidal genes upon different nutritional conditions. Front. Microbiol. 2016, 7, 805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashir, A.; Sun, Y.; Yu, X.; Sun, X.; Li, L. Nematicidal effects of 2-methyl-aconitate isomerase from the phytopathogen Pseudomonas syringae MB03 on the model nematode Caenorhabditis elegans. J. Invertebr. Pathol. 2021, 185, 107669. [Google Scholar] [CrossRef]
- Ali, M.; Gu, T.; Yu, X.; Bashir, A.; Wang, Z.; Sun, X.; Ashraf, N.; Li, L. Identification of the genes of the plant pathogen Pseudomonas syringae MB03 required for the nematicidal activity against Caenorhabditis elegans through an integrated approach. Front. Microbiol. 2022, 13, 826962. [Google Scholar] [CrossRef]
- Tedesco, P.; Di Schiavi, E.; Esposito, F.P.; de Pascale, D. Evaluation of Burkholderia cepacia complex bacteria pathogenicity using Caenorhabditis elegans. Bio Protoc. 2016, 6, e1964. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Yu, Z.; Zhang, S.; Xue, Z.; Huang, J.; Zhang, H.; Wan, X.; Chen, A.; Wang, J. Isolation and characterization of new 16-membered macrolides from the aveA3 gene replacement mutant strain Streptomyces avermitilis TM24 with acaricidal and nematicidal activities. J. Agric. Food Chem. 2019, 67, 4782–4792. [Google Scholar] [CrossRef]
- Huang, J.; Chen, A.L.; Zhang, H.; Yu, Z.; Li, M.H.; Li, N.; Lin, J.T.; Bai, H.; Wang, J.D.; Zheng, Y.G. Gene replacement for the generation of designed novel avermectin derivatives with enhanced acaricidal and nematicidal activities. Appl. Environ. Microbiol. 2015, 81, 5326–5334. [Google Scholar] [CrossRef] [Green Version]
- Phani, V.; Shivakumara, T.N.; Davies, K.G.; Rao, U. Meloidogyne incognita fatty acid- and retinol-binding protein (Mi-FAR-1) affects nematode infection of plant roots and the attachment of Pasteuria penetrans endospores. Front. Microbiol. 2017, 8, 2122. [Google Scholar] [CrossRef] [PubMed]
- Griffitts, J.S.; Haslam, S.M.; Yang, T.; Garczynski, S.F.; Mulloy, B.; Morris, H.; Cremer, P.S.; Dell, A.; Adang, M.J.; Aroian, R.V. Glycolipids as receptors for Bacillus thuringiensis crystal toxin. Science 2005, 307, 922–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Peng, D.; Zhang, F.; Ruan, L.; Sun, M. The Caenorhabditis elegans CUB-like-domain containing protein RBT-1 functions as a receptor for Bacillus thuringiensis Cry6Aa toxin. PLoS Pathog. 2020, 16, e1008501. [Google Scholar] [CrossRef]
- Manan, A.; Bazai, Z.A.; Fan, J.; Yu, H.; Li, L. The Nif3-family protein YqfO03 from Pseudomonas syringae MB03 has multiple nematicidal activities against Caenorhabditis elegans and Meloidogyne incognita. Int. J. Mol. Sci. 2018, 19, 3915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashir, A.; Tian, T.; Yu, X.; Meng, C.; Ali, M.; Li, L. Pyoverdine-mediated killing of Caenorhabditis elegans by Pseudomonas syringae MB03 and the role of iron in its pathogenicity. Int. J. Mol. Sci. 2020, 21, 2198. [Google Scholar] [CrossRef] [Green Version]
- Baidoo, R.; Mengistu, T.M.; Brito, J.A.; McSorley, R.; Stamps, R.H.; Crow, W.T. Vertical distribution of Pasteuria penetrans parasitizing Meloidogyne incognita on Pittosporum tobira in Florida. J. Nematol. 2017, 49, 311–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokalis-Burelle, N. Pasteuria penetrans for control of Meloidogyne incognita on tomato and cucumber, and M. arenaria on Snapdragon. J. Nematol. 2015, 47, 207–213. [Google Scholar] [PubMed]
- Kaletta, T.; Hengartner, M.O. Finding function in novel targets: C. elegans as a model organism. Nat. Reviron. Drug Discov. 2006, 5, 387. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.J.; Choi, N.S.; Choi, J.H.; Lee, J.S.; Yoon, J.H.; Song, J.J. Brevundimonas naejangsanensis sp. nov., a proteolytic bacterium isolated from soil, and reclassification of Mycoplana bullata into the genus Brevundimonas as Brevundimonas bullata comb. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 3155–3160. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Li, G.; Wang, X.; Pan, W.; Li, L.; LV, H.; Liu, F.; Dang, L.; Mo, M.; Zhang, K. Nematicidal endophytic bacteria obtained from plants. Ann. Microbiol. 2008, 58, 569–572. [Google Scholar] [CrossRef]
- Niu, D.-D.; Zheng, Y.; Zheng, L.; Jiang, C.-H.; Zhou, D.-M.; Guo, J.-H. Application of PSX biocontrol preparation confers root-knot nematode management and increased fruit quality in tomato under field conditions. Biocontrol. Sci. Technol. 2016, 26, 174–180. [Google Scholar] [CrossRef]
- De Oliveira, J.L.; Fraceto, L.F.; Bravo, A.; Polanczyk, R.A. Encapsulation strategies for Bacillus thuringiensis: From now to the future. J. Agric. Food Chem. 2021, 69, 4564–4577. [Google Scholar] [CrossRef]
- Melo, A.L.; Soccol, V.T.; Soccol, C.R. Bacillus thuringiensis: Mechanism of action, resistance, and new applications: A review. Crit. Rev. Biotechnol. 2016, 36, 317–326. [Google Scholar] [CrossRef]
- Siddiqui, Z.A.; Mahmood, I. Role of bacteria in the management of plant parasitic nematodes: A review. Bioresour. Technol. 1999, 69, 167–179. [Google Scholar] [CrossRef]
- Khanna, K.; Kohli, S.K.; Ohri, P.; Bhardwaj, R. Plants-nematodes-microbes crosstalk within soil: A trade-off among friends or foes. Microbiol. Res. 2021, 248, 126755. [Google Scholar] [CrossRef] [PubMed]
- Rosskopf, E.; Di Gioia, F.; Hong, J.C.; Pisani, C.; Kokalis-Burelle, N. Organic amendments for pathogen and nematode control. Annu. Rev. Phytopathol. 2020, 58, 277–311. [Google Scholar] [CrossRef]
- Bockenhoff, A.; Grundler, F. Studies on the nutrient uptake by the beet cyst nematode Heterodera schachtii by in situ microinjection of fluorescent probes into the feeding structures in Arabidopsis thaliana. Parasitology 1994, 109, 249–255. [Google Scholar] [CrossRef]
- Zhang, F.; Peng, D.; Ye, X.; Yu, Z.; Hu, Z.; Ruan, L.; Sun, M. In vitro uptake of 140 kDa Bacillus thuringiensis nematicidal crystal proteins by the second stage juvenile of Meloidogyne hapla. PLoS ONE 2012, 7, e38534. [Google Scholar] [CrossRef]
- Urwin, P.E.; Møller, S.G.; Lilley, C.J.; McPherson, M.J.; Atkinson, H.J. Continual green-fluorescent protein monitoring of cauliflower mosaic virus 35S promoter activity in nematode-induced feeding cells in Arabidopsis thaliana. Mol. Plant Microbe Interact. 1997, 10, 394–400. [Google Scholar] [CrossRef] [Green Version]
- Gross, H.; Loper, J.E. Genomics of secondary metabolite production by Pseudomonas spp. Nat. Prod. Rep. 2009, 26, 1408–1446. [Google Scholar] [CrossRef] [PubMed]
- Masschelein, J.; Jenner, M.; Challis, G.L. Antibiotics from Gram-negative bacteria: A comprehensive overview and selected biosynthetic highlights. Nat. Prod. Rep. 2017, 34, 712–783. [Google Scholar] [CrossRef]
- Barteczko, N.; Więcławik, J.; Tracz, A.; Pankalla, E.; Erfurt, K.; Latos, P.; Boncel, S.; Matuszek, K.; Chrobok, A. Dialkyl succinates and adipates as alternative plasticizers-even more efficient synthesis. Materials 2021, 14, 6219. [Google Scholar] [CrossRef]
- Perestrelo, R.; Silva, C.; Câmara, J.S. A useful approach for the differentiation of wines according to geographical origin based on global volatile patterns. J. Sep. Sci. 2014, 37, 1974–1981. [Google Scholar] [CrossRef] [PubMed]
- Borowiak-Resterna, A.; Cierpiszewski, R.; Prochaska, K. Kinetic and equilibrium studies of the removal of cadmium ions from acidic chloride solutions by hydrophobic pyridinecarboxamide extractants. J. Hazard. Mater. 2010, 179, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Bischof, L.J.; Huffman, D.L.; Aroian, R.V. Assays for toxicity studies in C. elegans with Bt crystal proteins. In Methods in Molecular Biology; Strange, K., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2006; Volume 351, pp. 139–154. [Google Scholar]
- Smith, H.R.; Brown, A. Benson’s Microbiological Applications Laboratory Manual, 15th ed.; McGraw Hill Companies: New York, NY, USA, 2021. [Google Scholar]
- Cai, M.; Dong, X. Systematic Identification Manual of Common Bacteria; Science Press: Beijing, China, 2001. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory: Laurel Hollow, NY, USA, 2001. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Marmur, J.; Doty, P. Determination of the base composition of deoxyribonucleic acid from its thermal denaturation temperature. J. Mol. Biol. 1962, 5, 109–118. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extractants | Corrected Mortality (%) 1 |
|---|---|
| Petroleum ether | 42.89 ± 5.55 |
| Chloroform phase | 12.55 ± 6.62 |
| Ethyl acetate phase | 84.57 ± 8.33 |
| n-butanol phase | 44.41 ± 2.09 |
| Aqueous phase | 27.55 ± 4.27 |
| MB756 SCS stock | 88.27 ± 6.38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Ding, M.; Sun, X.; Li, Z.; Xu, L.; Li, L. Characterization of Nematicidal Activity and Nematode-Toxic Metabolites of a Soilborne Brevundimonas bullata Isolate. Pathogens 2022, 11, 708. https://doi.org/10.3390/pathogens11060708
Li J, Ding M, Sun X, Li Z, Xu L, Li L. Characterization of Nematicidal Activity and Nematode-Toxic Metabolites of a Soilborne Brevundimonas bullata Isolate. Pathogens. 2022; 11(6):708. https://doi.org/10.3390/pathogens11060708
Chicago/Turabian StyleLi, Jiaoqing, Meijuan Ding, Xiaowen Sun, Zhe Li, Liangzheng Xu, and Lin Li. 2022. "Characterization of Nematicidal Activity and Nematode-Toxic Metabolites of a Soilborne Brevundimonas bullata Isolate" Pathogens 11, no. 6: 708. https://doi.org/10.3390/pathogens11060708
APA StyleLi, J., Ding, M., Sun, X., Li, Z., Xu, L., & Li, L. (2022). Characterization of Nematicidal Activity and Nematode-Toxic Metabolites of a Soilborne Brevundimonas bullata Isolate. Pathogens, 11(6), 708. https://doi.org/10.3390/pathogens11060708