
A Murine Model of Waning Scrub Typhus Cross-Protection between Heterologous Strains of Orientia tsutsugamushi


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
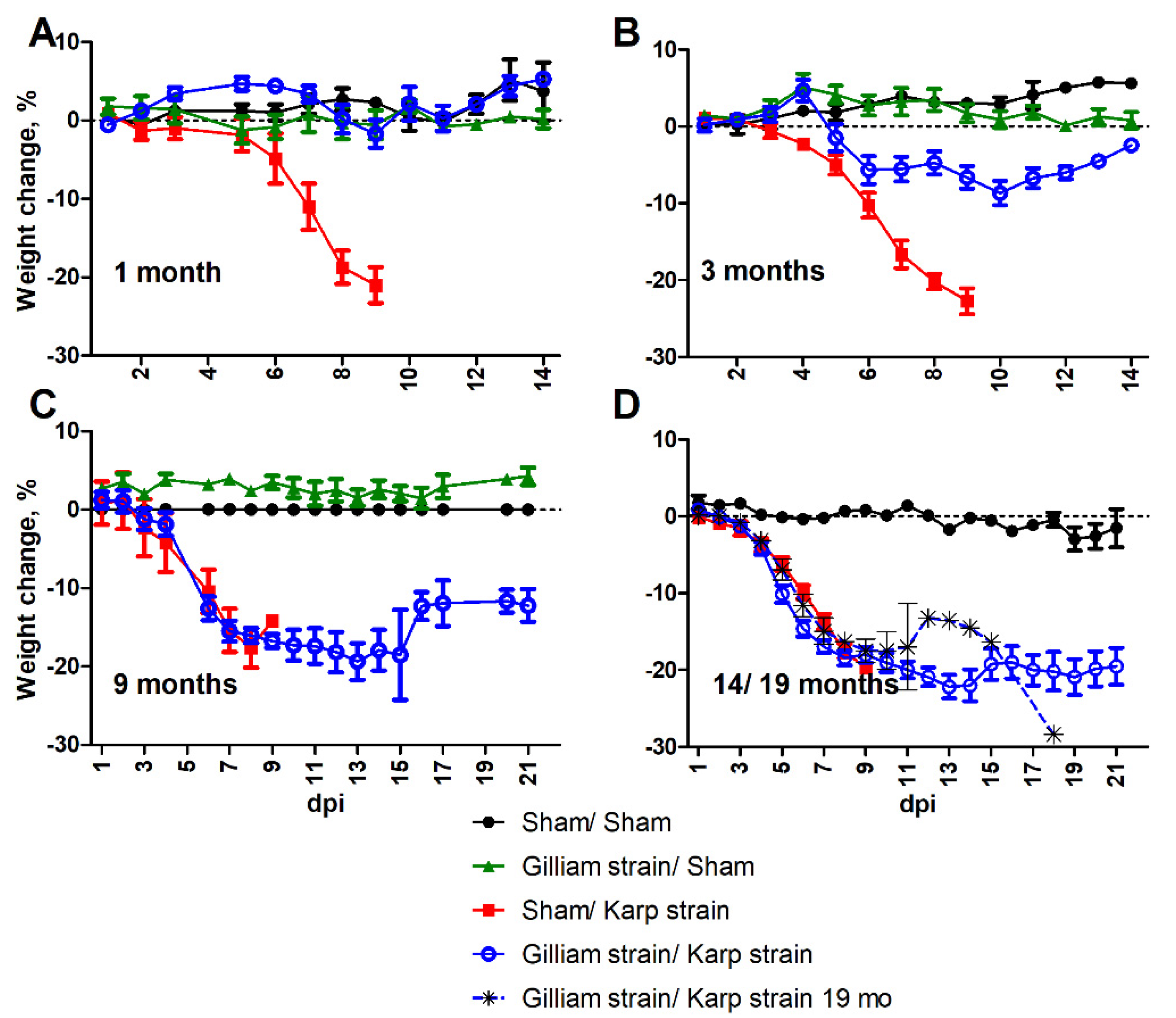
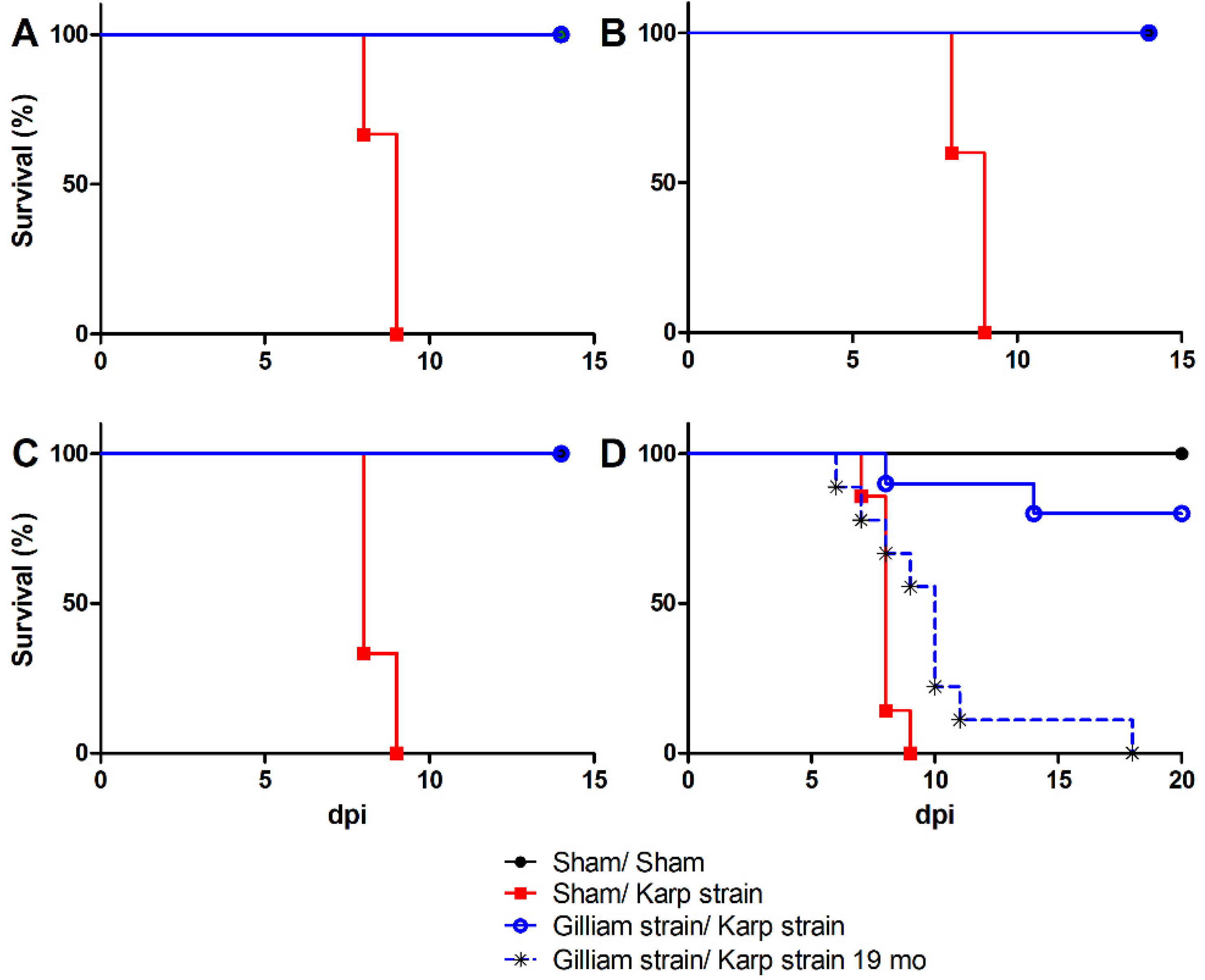
2.1. Loss of Protection and Development of Illness
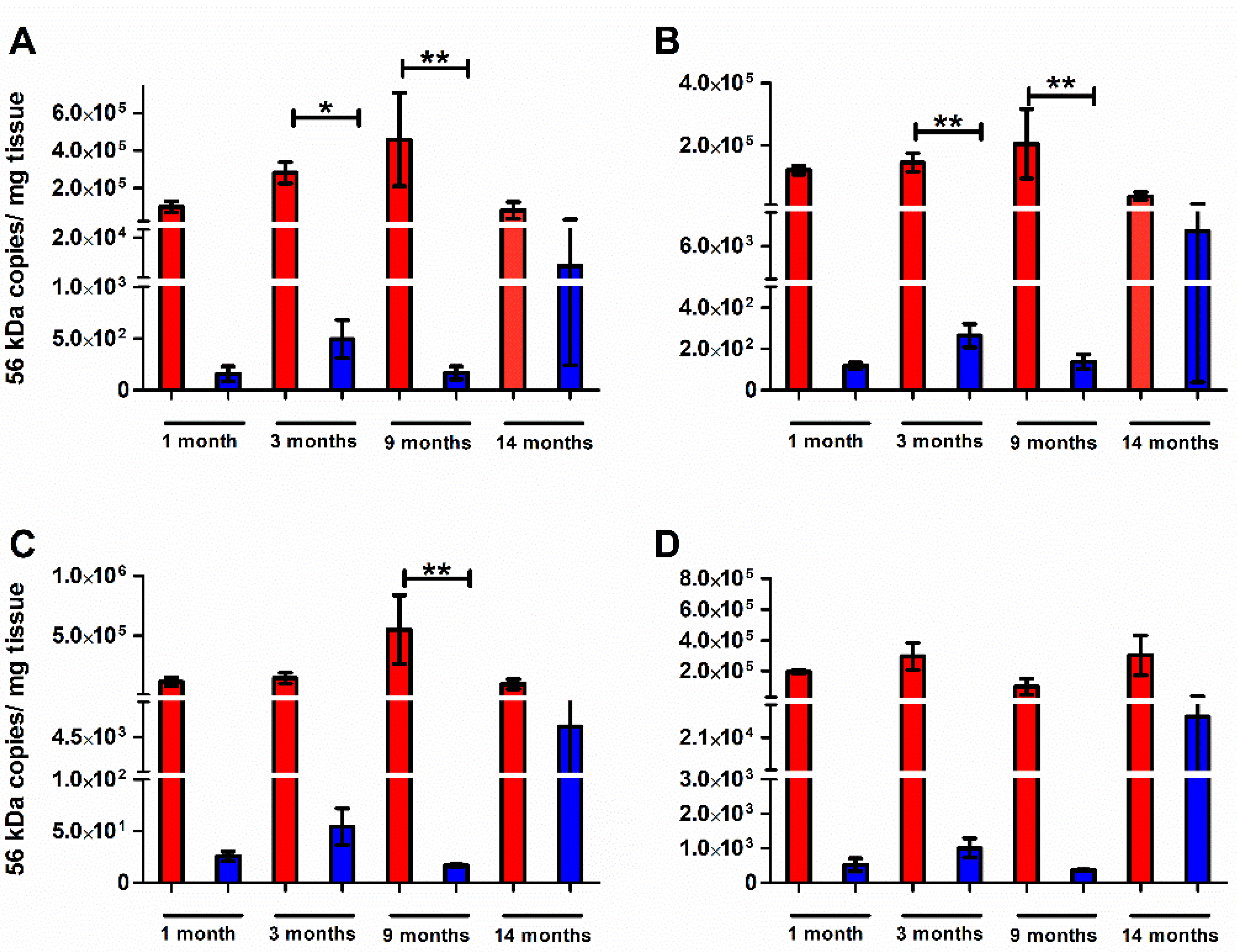
2.2. Bacterial Dissemination in Previously Challenged Mice
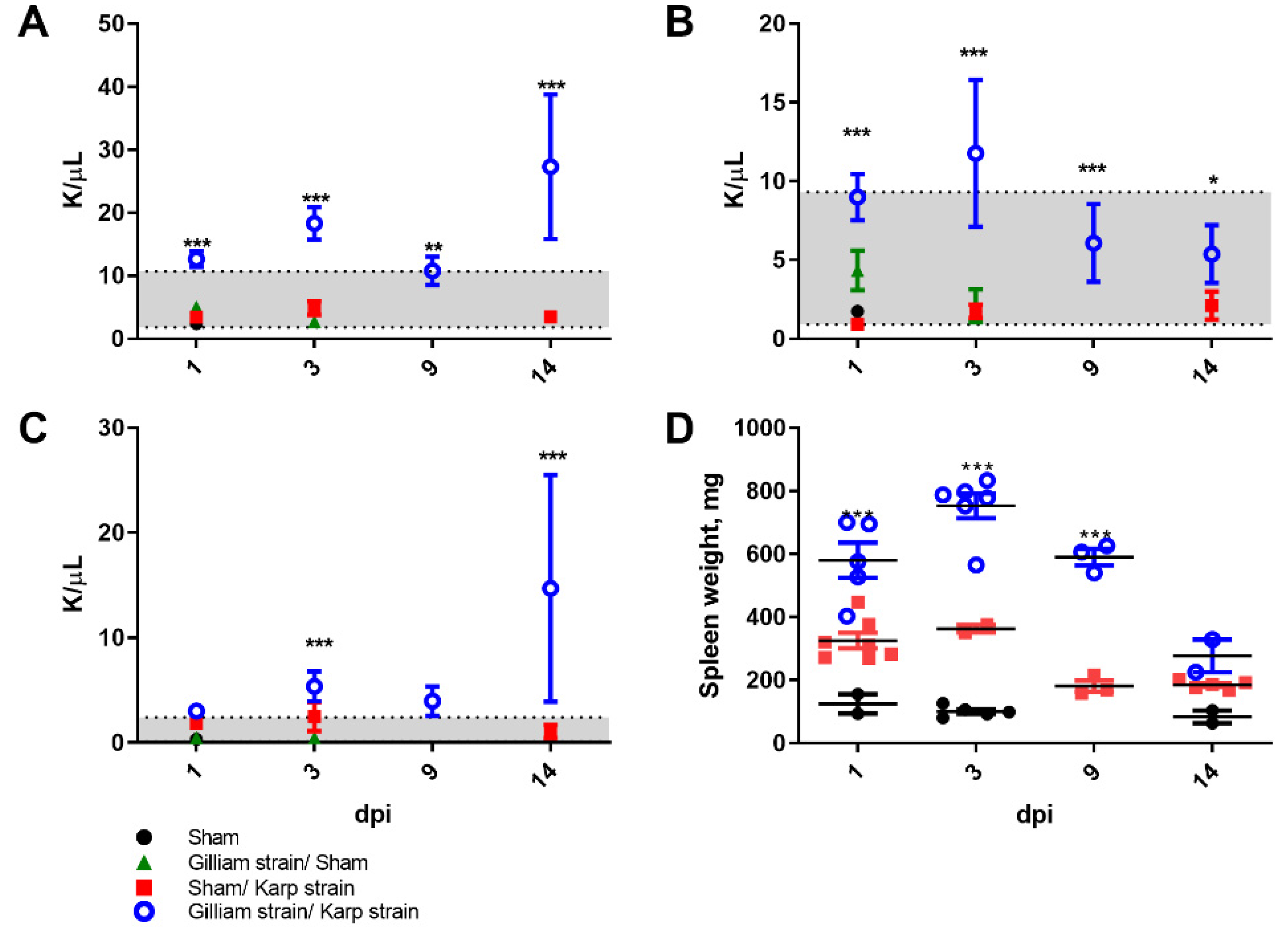
2.3. Physiologic Response to Heterologous Challenge
3. Discussion
4. Materials and Methods
4.1. Stock Propagation
4.2. Bacterial Viability and Load Determination
4.3. Mouse Infection
4.4. Hematologic Analyses
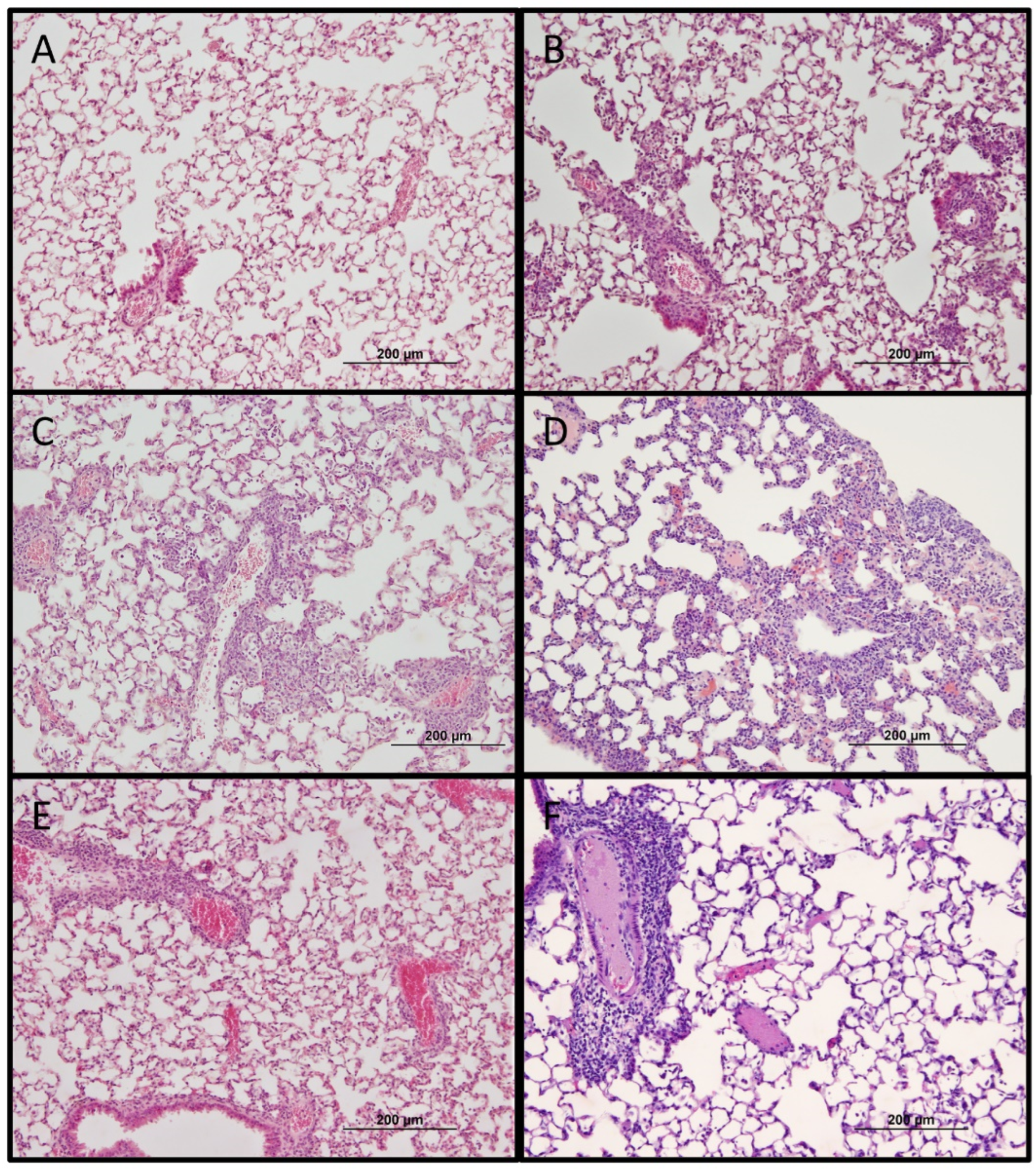
4.5. Histology
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kawamura, R. Studies on Tsutsushi Disease (Japanese flood fever). Med. Bull. Coll. Med. Univ. Cincinnati. 1926, 4, 1–229. [Google Scholar]
- Romeo, B.J. Convalescence from scrub typhus. Bull. United States Army Med. Dep. 1946, 6, 167–173. [Google Scholar]
- Kawamura, R.; Kasahara, S.; Toyama, T.; Nishinarita, F.; Tsubaki, S. On the prevention of tsutsugamushi. Results of preventive inoculations for people in the endemic region, and laboratory tests with the Pescadores strain. Kitasato Arch. Exp. Med. 1939, 16, 93–109. [Google Scholar]
- Smadel, J.E.; Ley, H.L., Jr.; Diercks, F.H.; Traub, R. Immunity in scrub typhus: Resistance to induced reinfection. Arch. Pathol. 1950, 50, 847–861. [Google Scholar]
- Smadel, J.E.; Ley, H.L., Jr.; Diercks, F.H.; Paterson, P.Y.; Wisseman, C.L., Jr.; Traub, R. Immunization against scrub typhus: Duration of immunity in volunteers following combined living vaccine and chemoprophylaxis. Am. J. Trop. Med. Hyg. 1952, 1, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Smadel, J.E.; Ley, H.L., Jr.; Diercks, F.H.; Traub, R.; Tipton, V.J.; Frick, L.P. Immunization against scrub typhus. I. Combined living vaccine and chemoprophylaxis in volunteers. Am. J. Hyg. 1951, 53, 317–325. [Google Scholar]
- Rights, F.L.; Smadel, J.E. Studies on scrub typhus (tsutsugamushi Disease): III. Heterogenicity of strains of R. tsutsugamushi as demonstrated by cross-vaccination studies. J. Exp. Med. 1948, 87, 339. [Google Scholar] [CrossRef] [Green Version]
- Smadel, J.E.; Ley, H.L., Jr.; Diercks, F.H.; Cameron, J.A.P. Persistence of Rickettsia tsutsugamushi in tissues of patients recovered from scrub typhus. Am. J. Hyg. 1952, 56, 294–302. [Google Scholar]
- MacMillan, J.G.; Rice, R.M.; Jerrells, T.R. Development of antigen-specific cell-mediated immune responses after infection of cynomolgus monkeys (Macaca fascicularis) with Rickettsia tsutsugamushi. J. Infect. Dis. 1985, 152, 739–749. [Google Scholar] [CrossRef]
- Van den Ende, M.; Locket, S.; Hargreaves, W.H.; Niven, J. Accidental laboratory infection with tsutsugamushi rickettsia. Lancet 1946, 248, 4–7. [Google Scholar] [CrossRef]
- Fulton, F.; Joyner, L. Cultivation of Rickettsia tsutsugamushi in lungs of rodents. Preparation of a Scrub-Typhus Vaccine. Lancet 1945, 246, 729–734. [Google Scholar] [CrossRef]
- Buckland, F.E.; Dudgeon, A.; Edward, D.G.; Henderson-Begg, A.; MacCallum, F.O.; Niven, J.S.F.; Rowlands, I.W.; Ende, M.; Bargmann, H.E.; Curtis, E.E. Scrub-typhus vaccine: Large-scale production. Lancet 1945, 246, 734–737. [Google Scholar] [CrossRef]
- Card, W.I.; Walker, J.M. Scrub-Typhus Vaccine: Field trial in South-East Asia. Lancet 1947, 249, 481–483. [Google Scholar] [CrossRef]
- Bengtson, I.A. Apparent serological heterogeneity among strains of tsutsugamushi disease (scrub typhus). Public Health Rep. 1945, 14, 1483–1488. [Google Scholar] [CrossRef]
- Bengtson, I.A. A serological study of 37 cases of tsutsugamushi disease (scrub typhus) occurring in Burma and the Philippine Islands. Public Health Rep. 1946, 61, 887–894. [Google Scholar] [CrossRef]
- Berge, T.O.; Gauld, R.L.; Kitaoka, M. A field trial of a vaccine prepared from the Volner strain of Rickettsia tsutsugamushi. Am. J. Hyg. 1949, 50, 337–342. [Google Scholar] [CrossRef]
- Eisenberg, G.H., Jr.; Osterman, J.V. Experimental scrub typhus immunogens: Gamma-irradiated and formalinized rickettsiae. Infect. Immun. 1977, 15, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, G.H., Jr.; Osterman, J.V. Gamma-irradiated scrub typhus immunogens: Development and duration of immunity. Infect. Immun. 1978, 22, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, G.H., Jr.; Osterman, J.V. Gamma-irradiated scrub typhus immunogens: Broad-spectrum immunity with combinations of rickettsial strains. Infect. Immun. 1979, 26, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Zarafonetis, C.J.D.; Snyder, J.C.; Murray, E.S. Immunity following para-aminobenzoic acid therapy in experimental tsutsugamushi disease (Scrub typhus). Proc. Soc. Exp. Biol. Med. 1946, 61, 240–242. [Google Scholar] [CrossRef]
- Topping, N.H. Cross immunity between four strains of tsutsugamushi disease (scrub typhus). Public Health Rep. 1945, 60, 945–947. [Google Scholar] [CrossRef]
- Soong, L.; Shelite, T.R.; Xing, Y.; Kodakandla, H.; Liang, Y.; Trent, B.J.; Horton, P.; Smith, K.C.; Zhao, Z.; Sun, J. Type 1-skewed neuroinflammation and vascular damage associated with Orientia tsutsugamushi infection in mice. PLoS Negl. Trop. Dis. 2017, 11, e0005765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelite, T.R.; Saito, T.B.; Mendell, N.L.; Gong, B.; Xu, G.; Soong, L.; Valbuena, G.; Bouyer, D.H.; Walker, D.H. A hematogenously disseminated Orientia tsutsugamsushi-infected murine model of scrub typhus. PLoS Negl. Trop. Dis. 2014, 8, e2966. [Google Scholar] [CrossRef] [Green Version]
- Mendell, N.L.; Bouyer, D.H.; Walker, D.H. Murine models of scrub typhus associated with host control of Orientia tsutsugamushi infection. PLoS Negl. Trop. Dis. 2017, 11, e0005453. [Google Scholar] [CrossRef]
- Soong, L.; Mendell, N.L.; Olano, J.P.; Rockx-Brouwer, D.; Xu, G.; Goez-Rivillas, Y.; Drom, C.; Shelite, T.R.; Valbuena, G.; Walker, D.H. An intradermal inoculation mouse model for immunological investigations of acute scrub typhus and persistent infection. PLoS Negl. Trop. Dis. 2016, 10, e0004884. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendell, N.L.; Xu, G.; Shelite, T.R.; Bouyer, D.H.; Walker, D.H. A Murine Model of Waning Scrub Typhus Cross-Protection between Heterologous Strains of Orientia tsutsugamushi. Pathogens 2022, 11, 512. https://doi.org/10.3390/pathogens11050512
Mendell NL, Xu G, Shelite TR, Bouyer DH, Walker DH. A Murine Model of Waning Scrub Typhus Cross-Protection between Heterologous Strains of Orientia tsutsugamushi. Pathogens. 2022; 11(5):512. https://doi.org/10.3390/pathogens11050512
Chicago/Turabian StyleMendell, Nicole L., Guang Xu, Thomas R. Shelite, Donald H. Bouyer, and David H. Walker. 2022. "A Murine Model of Waning Scrub Typhus Cross-Protection between Heterologous Strains of Orientia tsutsugamushi" Pathogens 11, no. 5: 512. https://doi.org/10.3390/pathogens11050512
APA StyleMendell, N. L., Xu, G., Shelite, T. R., Bouyer, D. H., & Walker, D. H. (2022). A Murine Model of Waning Scrub Typhus Cross-Protection between Heterologous Strains of Orientia tsutsugamushi. Pathogens, 11(5), 512. https://doi.org/10.3390/pathogens11050512