
Protozoan and Rickettsial Pathogens in Ticks Collected from Infested Cattle from Turkey


 ,
,  , ,
, ,  ,
,  and
and 
Abstract
:
1. Introduction
2. Results
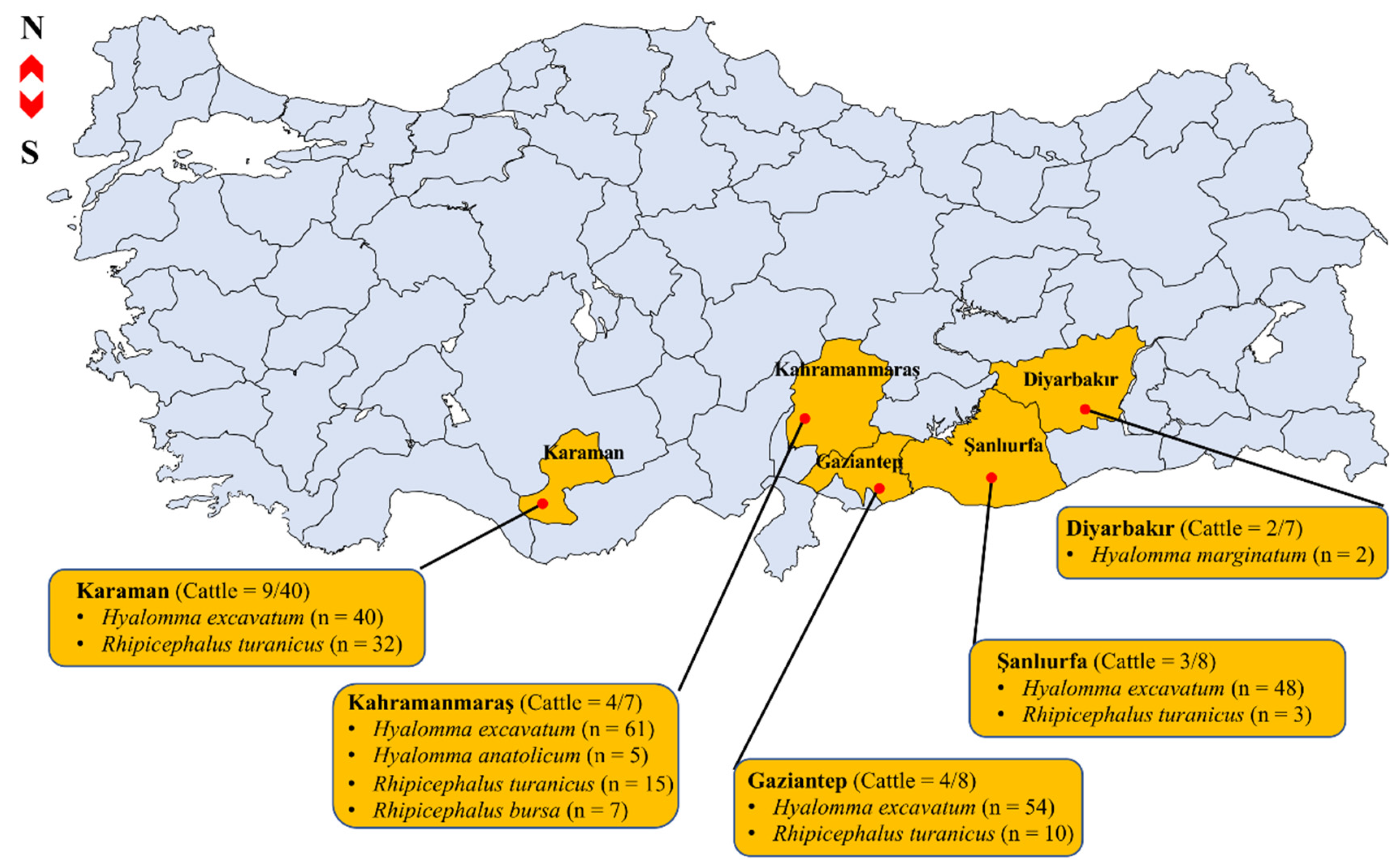
2.1. Tick Species Identification
2.2. Microorganisms Detected in Ticks and Minimum Detection Rates
2.3. Analyses of Tick-Borne Pathogen DNA Sequences
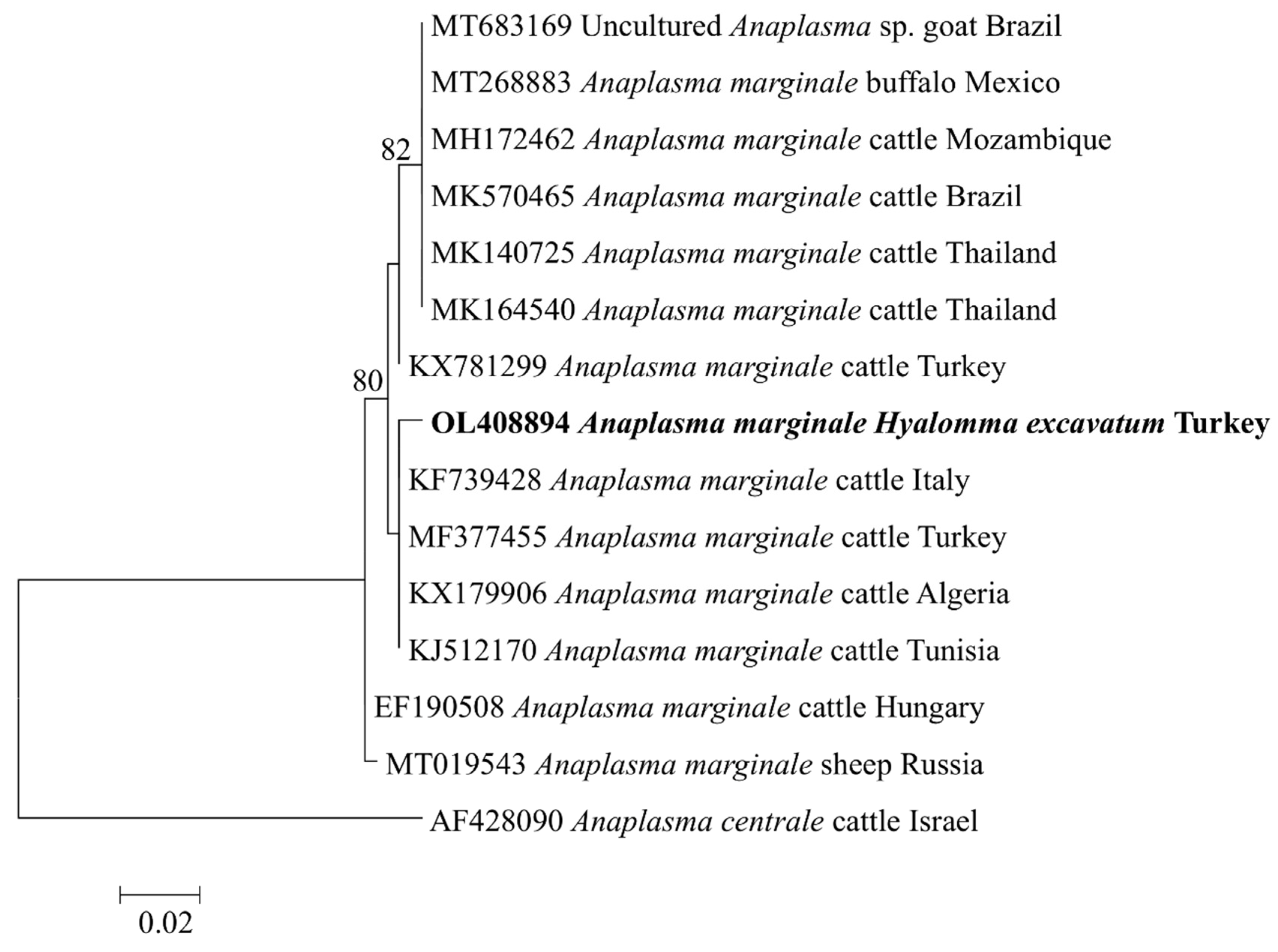
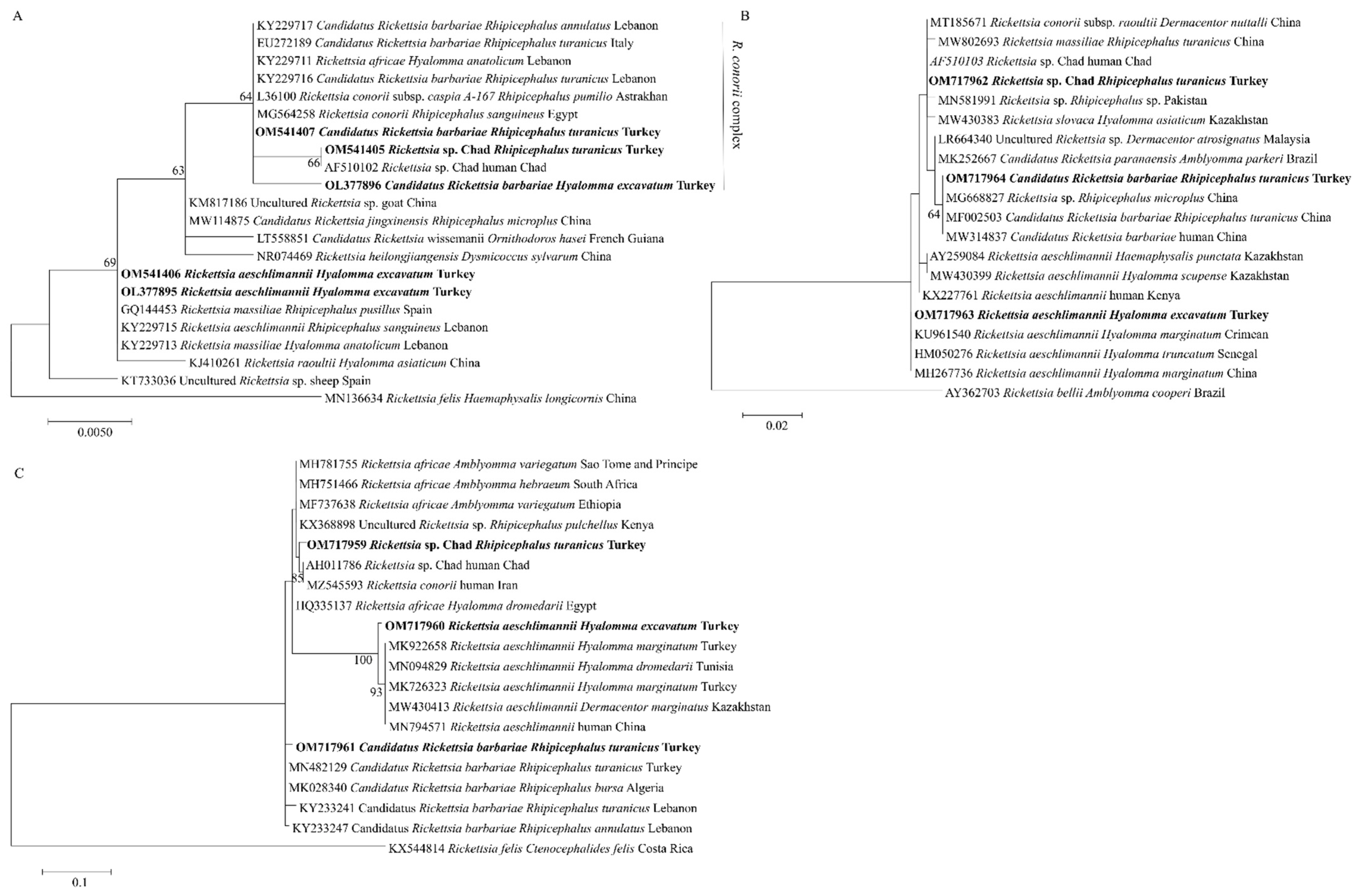
2.4. Phylogenetic Analyses of Rickettsial Agents
3. Discussion
4. Materials and Methods
4.1. Ethical Statements
4.2. Tick Samples and DNA Extraction
4.3. Molecular Characterization of Ticks
4.4. Detection and Characterization of Tick-Borne Pathogens
4.5. Cloning and Sequencing
4.6. Sequence Alignment and Phylogenetic Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- De la Fuente, J.; Estrada-Pena, A.; Venzal, J.M.; Kocan, K.M.; Sonenshine, D.E. Overview: Ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 2008, 13, 6938–6946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada-Peña, A.; Jongejan, F. Ticks feeding on humans: A review of records on human-biting Ixodoidea with special reference to pathogen transmission. Exp. Appl. Acarol. 1999, 23, 685–715. [Google Scholar] [CrossRef] [PubMed]
- Sparagano, O.A.; Allsopp, M.T.; Mank, R.A.; Rijpkema, S.G.; Figueroa, J.V.; Jongejan, F. Molecular detection of pathogen DNA in ticks (Acari: Ixodidae): A review. Exp. Appl. Acarol. 1999, 23, 929–960. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F.; Chomel, B.B.; Otranto, D. Ticks and tick-borne diseases: A One Health perspective. Trends. Parasitol. 2012, 28, 437–446. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, H.; Wang, T.; Sun, W.; Yang, X.; Liu, J. Tick-borne pathogens and the vector potential of ticks in China. Parasites Vectors 2015, 8, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dantas-Torres, F. Species Concepts: What about Ticks? Trends. Parasitol. 2018, 34, 1017–1026. [Google Scholar] [CrossRef]
- Sayin, F.; Dinçer, S.; Karaer, Z.; Cakmak, A.; Inci, A.; Yukari, B.A.; Eren, H.; Vatansever, Z.; Nalbantoglu, S. Studies on the epidemiology of tropical theileriosis (Theileria annulata infection) in cattle in Central Anatolia, Turkey. Trop. Anim. Health. Prod. 2003, 35, 521–539. [Google Scholar] [CrossRef]
- Vatansever, Z.; Gargili, A.; Aysul, N.S.; Sengoz, G.; Estrada-Peña, A. Ticks biting humans in the urban area of Istanbul. Parasitol. Res. 2008, 102, 551–553. [Google Scholar] [CrossRef]
- Bursali, A.; Keskin, A.; Tekin, S. A review of the ticks (Acari: Ixodida) of Turkey: Species diversity, hosts and geographical distribution. Exp. Appl. Acarol. 2012, 57, 91–104. [Google Scholar] [CrossRef]
- Ceylan, O.; Xuan, X.; Sevinc, F. Primary Tick-Borne Protozoan and Rickettsial Infections of Animals in Turkey. Pathogens 2021, 10, 231. [Google Scholar] [CrossRef]
- Ozubek, S.; Bastos, R.G.; Alzan, H.F.; Inci, A.; Aktas, M.; Suarez, C.E. Bovine babesiosis in Turkey: Impact, current gaps, and opportunities for intervention. Pathogens 2020, 9, 1041. [Google Scholar] [CrossRef] [PubMed]
- Altay, K.; Dumanli, N.; Holman, P.J.; Aktas, M. Detection of Theileria ovis in naturally infected sheep by nested PCR. Vet. Parasitol. 2005, 127, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Altay, K.; Dumanli, N.; Aktas, M. Molecular identification, genetic diversity and distribution of Theileria and Babesia species infecting small ruminants. Vet. Parasitol. 2007, 147, 161–165. [Google Scholar] [CrossRef]
- Altay, K.; Aydin, M.F.; Dumanli, N.; Aktas, M. Molecular detection of Theileria and Babesia infections in cattle. Vet. Parasitol. 2008, 158, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Aktas, M.; Altay, K.; Dumanli, N. Molecular detection and identification of Anaplasma and Ehrlichia species in cattle from Turkey. Ticks Tick Borne Dis. 2011, 2, 62–65. [Google Scholar] [CrossRef]
- Kocan, K.M.; de la Fuente, J.; Cabezas-Cruz, A. The genus Anaplasma: New challenges after reclassification. Rev. Sci. Tech. 2015, 34, 577–586. [Google Scholar] [CrossRef]
- Aydin, M.F.; Özübek, S.; Aktaş, M. Molecular survey of Anaplasma and Ehrlichia species in cattle from Karaman of Turkey, including a novel tandem report of Anaplasma marginale msp1a gene. Ankara Üniv. Vet. Fak. Derg. 2019, 66, 255–260. [Google Scholar] [CrossRef]
- De la Fuente, J.; Van Den Bussche, R.A.; Kocan, K.M. Molecular phylogeny and biogeography of North American isolates of Anaplasma marginale (Rickettsiaceae: Ehrlichieae). Vet. Parasitol. 2001, 97, 65–76. [Google Scholar] [CrossRef]
- De la Fuente, J.; Passos, L.M.; Van Den Bussche, R.A.; Ribeiro, M.F.; Facury-Filho, E.J.; Kocan, K.M. Genetic diversity and molecular phylogeny of Anaplasma marginale isolates from Minas Gerais, Brazil. Vet. Parasitol. 2004, 121, 307–316. [Google Scholar] [CrossRef]
- Birdane, F.M.; Sevinc, F.; Derinbay, O. Anaplasma marginale infections in dairy cattle: Clinical disease with high seroprevalence. Bull. Vet. Inst. Pulawy. 2006, 50, 467–470. [Google Scholar]
- Aktas, M.; Altay, K.; Ozubek, S.; Dumanli, N. A survey of ixodid ticks feeding on cattle and prevalence of tick-borne pathogens in the Black Sea region of Turkey. Vet. Parasitol. 2012, 187, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Aktas, M. A survey of ixodid tick species and molecular identification of tick-borne pathogens. Vet. Parasitol. 2014, 200, 276–283. [Google Scholar] [CrossRef]
- Aktas, M.; Vatansever, Z.; Ozubek, S. Molecular evidence for trans-stadial and transovarial transmission of Babesia occultans in Hyalomma marginatum and Rhipicephalus turanicus in Turkey. Vet. Parasitol. 2014, 204, 369–371. [Google Scholar] [CrossRef] [PubMed]
- Aktas, M.; Özübek, S. Bovine anaplasmosis in Turkey: First laboratory confirmed clinical cases caused by Anaplasma phagocytophilum. Vet. Microbiol. 2015, 178, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.S.; de Vos, A.J. Studies on a bovine Babesia transmitted by Hyalomma marginatum rufipes Koch, 1844. Onderstepoort J. Vet. Res. 1981, 48, 215–223. [Google Scholar] [PubMed]
- Dipeolu, O.O.; Amoo, A. The presence of kinetes of a Babesia species in the haemolymph smears of engorged Hyalomma ticks in Nigeria. Vet. Parasitol. 1984, 17, 41–46. [Google Scholar] [CrossRef]
- Orkun, O. Molecular investigation of the natural transovarial transmission of tick-borne pathogens in Turkey. Vet. Parasitol. 2019, 273, 97–104. [Google Scholar] [CrossRef]
- Kilicoglu, Y.; Cagirgan, A.A.; Serdar; Kaya, S.; Durmaz, Y.; Gur, Y. Molecular investigation, isolation and phylogenetic analsysis of Coxiella burnetii from aborted fetus and ticks. Comp. Immunol. Microbiol. Infect. Dis. 2020, 73, 101571. [Google Scholar] [CrossRef]
- Altay, Ç.G.; Emre, Z.; Canpolat, S.; Vatansever, Y.; Düzgün, A. Detection of Coxiella burnetii from ticks by polymerase chain reaction and restriction fragment length polymorphism. Ankara. Univ. Vet. Fak. Derg. 2013, 60, 263–268. [Google Scholar] [CrossRef]
- Esemu, S.N.; Besong, W.O.; Ndip, R.N.; Ndip, L.M. Prevalence of Ehrlichia ruminantium in adult Amblyomma variegatum collected from cattle in Cameroon. Exp. Appl. Acarol. 2013, 59, 377–387. [Google Scholar] [CrossRef]
- Mtshali, K.; Khumalo, Z.; Nakao, R.; Grab, D.; Sugimoto, C.; Thekisoe, O. Molecular detection of zoonotic tick-borne pathogens from ticks collected from ruminants in four South African provinces. J. Vet. Med. Sci. 2015, 77, 1573–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, P.F.; Niu, Q.L.; Liu, Z.J.; Yang, J.F.; Chen, Z.; Guan, G.Q.; Liu, G.Y.; Luo, J.X.; Yin, H. Molecular epidemiological surveillance to assess emergence and re-emergence of tick-borne infections in tick samples from China evaluated by nested PCRs. Acta Trop. 2016, 158, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Orkun, O.; Çakmak, A.; Nalbantoğlu, S.; Karaer, Z. Turkey tick news: A molecular investigation into the presence of tick-borne pathogens in host-seeking ticks in Anatolia; Initial evidence of putative vectors and pathogens, and footsteps of a secretly rising vector tick, Haemaphysalis parva. Ticks Tick Borne Dis. 2020, 11, 101373. [Google Scholar] [CrossRef] [PubMed]
- Orkun, O.; Emir, H. Identification of tick-borne pathogens in ticks collected from wild animals in Turkey. Parasitol. Res. 2020, 119, 3083–3091. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.E.; Xeridat, B.; Raoult, D. Isolation of a Rickettsia related to Astrakhan fever rickettsia from a patient in Chad. Ann. N. Y. Acad. Sci. 2003, 990, 152–157. [Google Scholar] [CrossRef]
- Ferrolho, J.; Antunes, S.; Santos, A.S.; Velez, R.; Padre, L.; Cabezas-Cruz, A.; Santos-Silva, M.M.; Domingos, A. Detection and phylogenetic characterization of Theileria spp. and Anaplasma marginale in Rhipicephalus bursa in Portugal. Ticks Tick Borne Dis. 2016, 7, 443–448. [Google Scholar] [CrossRef]
- Duron, O.; Binetruy, F.; Noël, V.; Cremaschi, J.; McCoy, K.D.; Arnathau, C.; Plantard, O.; Goolsby, J.; Pérez de León, A.A.; Heylen, D.J.A.; et al. Evolutionary changes in symbiont community structure in ticks. Mol. Ecol. 2017, 26, 2905–2921. [Google Scholar] [CrossRef] [Green Version]
- Karim, S.; Budachetri, K.; Mukherjee, N.; Williams, J.; Kausar, A.; Hassan, M.J.; Adamson, S.; Dowd, S.E.; Apanskevich, D.; Arijo, A.; et al. A study of ticks and tick-borne livestock pathogens in Pakistan. PLoS Negl. Trop. Dis. 2017, 11, e0005681. [Google Scholar] [CrossRef]
- Ghafar, A.; Cabezas-Cruz, A.; Galon, C.; Obregon, D.; Gasser, R.B.; Moutailler, S.; Jabbar, A. Bovine ticks harbour a diverse array of microorganisms in Pakistan. Parasit. Vectors 2020, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Gottlieb, Y.; Lalzar, I.; Klasson, L. Distinctive Genome Reduction Rates Revealed by Genomic Analyses of Two Coxiella-Like Endosymbionts in Ticks. Genome Biol. Evol. 2015, 7, 1779–1796. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Peña, A.; Bouattour, A.; Camicas, J.L.; Walker, A.R. Ticks of Domestic Animals in the Mediterranean Region: A Guide to Identification of Species; University of Zaragoza: Zaragoza, Spain, 2004. [Google Scholar]
- Walker, A.R.; Bouattour, A.; Camicas, J.J.; Estrada-pena, A.; Horak, I.G.; Latif, A.A.; Pegram, R.G.; Preston, P.M. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; Bioscience Reports: Edinburgh, UK, 2003; pp. 1–1221. [Google Scholar]
- Adjou Moumouni, P.F.; Terkawi, M.A.; Jirapattharasate, C.; Cao, S.; Liu, M.; Nakao, R.; Umemiya-Shirafuji, R.; Yokoyama, N.; Sugimoto, C.; Fujisaki, K.; et al. Molecular detection of spotted fever group rickettsiae in Amblyomma variegatum ticks from Benin. Ticks Tick Borne Dis. 2016, 7, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Black, W.C.; Piesman, J. Phylogeny of hard- and soft-tick taxa (Acari: Ixodida) based on mitochondrial 16S rDNA sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 10034–10038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georges, K.; Loria, G.R.; Riili, S.; Greco, A.; Caracappa, S.; Jongejan, F. Detection of haemoparasites in cattle by reverse line blot hybridisation with a note on the distribution of ticks in Sicily. Vet. Parasitol. 2001, 99, 273–286. [Google Scholar] [CrossRef]
- Tay, S.T.; Koh, F.X.; Kho, K.L.; Ong, B.L. Molecular survey and sequence analysis of Anaplasma spp. in cattle and ticks in a Malaysian farm. Trop. Biomed. 2014, 31, 769–776. [Google Scholar] [PubMed]
- Pesquera, C.; Portillo, A.; Palomar, A.M.; Oteo, J.A. Investigation of tick-borne bacteria (Rickettsia spp., Anaplasma spp., Ehrlichia spp. and Borrelia spp.) in ticks collected from Andean tapirs, cattle and vegetation from a protected area in Ecuador. Parasit. Vectors 2015, 8, 46. [Google Scholar] [CrossRef] [Green Version]
- Terkawi, M.A.; Huyen, N.X.; Shinuo, C.; Inpankaew, T.; Maklon, K.; Aboulaila, M. Molecular and serological prevalence of Babesia bovis and Babesia bigemina in water buffaloes in the northeast region of Thailand. Vet. Parasitol. 2011, 178, 201–207. [Google Scholar] [CrossRef]
- Ota, N.; Mizuno, D.; Kuboki, N.; Igarashi, I.; Nakamura, Y.; Yamashina, H. Epidemiological survey of Theileria orientalis infection in grazing cattle in the eastern part of Hokkaido, Japan. J. Vet. Med. Sci. 2009, 71, 937–944. [Google Scholar] [CrossRef] [Green Version]
- Martín-Sánchez, J.; Viseras, J.; Adroher, F.J.; García-Fernández, P. Nested polymerase chain reaction for detection of Theileria annulata and comparison with conventional diagnostic techniques: Its use in epidemiology studies. Parasitol. Res. 1999, 85, 243–245. [Google Scholar] [CrossRef]
- Ybañez, A.P.; Sivakumar, T.; Battsetseg, B.; Battur, B.; Altangerel, K.; Matsumoto, K.; Yokoyama, N.; Inokuma, H. Specific molecular detection and characterization of Anaplasma marginale in Mongolian cattle. J. Vet. Med. Sci. 2013, 75, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Tumwebaze, M.A.; Byamukama, B.; Tayebwa, D.S.; Byaruhanga, J.; Angwe, M.K.; Galon, E.M.; Liu, M.; Lee, S.H.; Ringo, A.E.; Adjou Moumouni, P.F.; et al. First molecular detection of Babesia ovis, Theileria spp., Anaplasma spp., and Ehrlichia ruminantium in goats from western Uganda. Pathogens 2020, 9, 895. [Google Scholar] [CrossRef]
- Zheng, W.; Liu, M.; Moumouni, P.F.; Liu, X.; Efstratiou, A.; Liu, Z.; Liu, Y.; Tao, H.; Guo, H.; Wang, G.; et al. First molecular detection of tick-borne pathogens in dogs from Jiangxi, China. J. Vet. Med. Sci. 2017, 79, 248–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labruna, M.B.; Whitworth, T.; Horta, M.C.; Bouyer, D.H.; McBride, J.W.; Pinter, A.; Popov, V.; Gennari, S.M.; Walker, D.H. Rickettsia species infecting Amblyomma cooperi ticks from an area in the state of São Paulo, Brazil, where Brazilian spotted fever is endemic. J. Clin. Microbiol. 2004, 42, 90–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anstead, C.A.; Chilton, N.B. A novel Rickettsia species detected in Vole Ticks (Ixodes angustus) from Western Canada. Appl. Environ. Microbiol. 2013, 79, 7583–7589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringo, A.E.; Adjou Moumouni, P.F.; Lee, S.H.; Liu, M.; Khamis, Y.H.; Gao, Y.; Guo, H.; Zheng, W.; Efstratiou, A.; Galon, E.M.; et al. Molecular detection and characterization of tick-borne protozoan and rickettsial pathogens isolated from cattle on Pemba Island, Tanzania. Ticks Tick Borne Dis. 2018, 9, 1437–1445. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Rannala, B. Molecular phylogenetics: Principles and practice. Nat. Rev. Genet. 2012, 13, 303–314. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Province | Tick Species (Positive Pools/Total Pools) | Microorganism Detected (No. of Pools) | Minimum Detection Rate |
|---|---|---|---|
| Gaziantep | Hyalomma excavatum (11/17) | Anaplasma marginale (4) | |
| Babesia bovis (8) | |||
| Ehrlichia sp. (3) | |||
| Theileria annulata (4) | A. marginale 2.5% (7/277) | ||
| Rhipicephalus turanicus (1/8) | Ehrlichia sp. (1) | ||
| Kahramanmaraş | H. excavatum (16/16) | A. marginale (1) | |
| B. bovis (11) | |||
| Coxiella sp. (6) | B. bovis 7.9% (22/277) | ||
| Ehrlichia sp. (4) | |||
| Rickettsia sp. (3) | |||
| T. annulata (4) | |||
| Rh. turanicus (4/6) | A. marginale (1) | B. occultans 0.7% (2/277) | |
| B. bovis (1) | |||
| Coxiella sp. (2) | |||
| Ehrlichia sp. (1) | |||
| Rickettsia sp. (2) | Coxiella sp. 3.3% (9/277) | ||
| Rh. bursa (2/2) | B. bovis (1) | ||
| Coxiella sp. (1) | |||
| Ehrlichia sp. (1) | |||
| H. anatolicum (2/2) | B. bovis (1) | Ehrlichia sp. 7.2% (20/277) | |
| Ehrlichia sp. (1) | |||
| T. annulata (2) | |||
| Şanlıurfa | H. excavatum (8/13) | B. occultans (1) | |
| Ehrlichia sp. (6) | Rickettsia sp. 2.5% (7/277) | ||
| T. annulata (5) | |||
| Rh. turanicus (2/3) | Ehrlichia sp. (1) | ||
| T. annulata (1) | |||
| Karaman | H. excavatum (1/13) | B. occultans (1) | T. annulata 5.8% (16/277) |
| Rh. turanicus (5/9) | A. marginale (1) | ||
| Ehrlichia sp. (2) | |||
| Rickettsia sp. (2) |
| DNA Sequences | High BLASTn Match | ||||||
|---|---|---|---|---|---|---|---|
| Pathogen | Target Gene | Accession Number | Length (bp) | Query Cover (%) | Identity (%) | Reference Strains (Accession No.) | Source |
| Anaplasma marginale | msp4 | OL408894 | 350 | 99 | 100 | A. marginale (MF377455) | Cattle, Turkey |
| Babesia bovis | sbp4 | OL408893 | 512 | 100 | 100 | B. bovis (MN870661) | Buffalo, Egypt |
| B. occultans | 18S rRNA (V4) | OL377855 | 403 | 100 | 100 | B. occultans (KP745626) | Cattle, Turkey |
| Coxiella sp. | 16S rRNA | OL413002 | 1,453 | 100 | 99.66 | Candidatus Coxiella mudrowiae (CP011126) | Rh. turanicus, Israel |
| Ehrlichia sp. | pCS20 | OL408895 | 279 | 100 | 99.28 | Uncultured Ehrlichia sp. (KT362172) | Rhipicephalus microplus, China |
| Rickettsia sp. | 16S rRNA | OL377895 | 436 | 99 | 99.77 | R. aeschlimannii (KY229715) | Rh. sanguineus, Lebanon |
| OL377896 | 439 | 100 | 99.77 | Candidatus Rickettsia barbariae (EU272189) | Rh. turanicus, Italy | ||
| OM541405 | 366 | 98 | 100 | Rickettsia sp. Chad (AF510102) | Human, Chad | ||
| OM541406 | 436 | 98 | 99.29 | R. aeschlimannii (KY229715) | Rh. sanguineus, Lebanon | ||
| OM541407 | 439 | 99 | 99.77 | Candidatus Rickettsia barbariae (EU272189) | Rh. turanicus, Italy | ||
| gltA | OM717962 | 401 | 100 | 100 | Rickettsia sp. Chad (AF510103) | Human, Chad | |
| OM717963 | 401 | 100 | 100 | R. aeschlimannii (KU961540) | H. marginatum, Crimea | ||
| OM717964 | 401 | 100 | 100 | Candidatus Rickettsia barbariae (MF002503) | Rh. turanicus, China | ||
| ompA | OM717959 | 210 | 97 | 99.05 | Rickettsia sp. Chad (AH011786) | Human, Chad | |
| OM717960 | 210 | 98 | 99.52 | R. aeschlimannii (MK922658) | H. marginatum, Turkey | ||
| OM717961 | 210 | 99 | 100 | Candidatus Rickettsia barbariae (MN482129) | Rh. turanicus, Turkey | ||
| Theileria annulata | Tams-1 | OL408892 | 452 | 95 | 99.07 | T. annulata (KX130956) | Dog, Tunisia |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, S.; Ceylan, O.; Ma, Z.; Galon, E.M.; Zafar, I.; Li, H.; Hasegawa, Y.; Sevinc, M.; Masatani, T.; Iguchi, A.; et al. Protozoan and Rickettsial Pathogens in Ticks Collected from Infested Cattle from Turkey. Pathogens 2022, 11, 500. https://doi.org/10.3390/pathogens11050500
Ji S, Ceylan O, Ma Z, Galon EM, Zafar I, Li H, Hasegawa Y, Sevinc M, Masatani T, Iguchi A, et al. Protozoan and Rickettsial Pathogens in Ticks Collected from Infested Cattle from Turkey. Pathogens. 2022; 11(5):500. https://doi.org/10.3390/pathogens11050500
Chicago/Turabian StyleJi, Shengwei, Onur Ceylan, Zhuowei Ma, Eloiza May Galon, Iqra Zafar, Hang Li, Yae Hasegawa, Mutlu Sevinc, Tatsunori Masatani, Aiko Iguchi, and et al. 2022. "Protozoan and Rickettsial Pathogens in Ticks Collected from Infested Cattle from Turkey" Pathogens 11, no. 5: 500. https://doi.org/10.3390/pathogens11050500
APA StyleJi, S., Ceylan, O., Ma, Z., Galon, E. M., Zafar, I., Li, H., Hasegawa, Y., Sevinc, M., Masatani, T., Iguchi, A., Kawase, O., Umemiya-Shirafuji, R., Asada, M., Sevinc, F., & Xuan, X. (2022). Protozoan and Rickettsial Pathogens in Ticks Collected from Infested Cattle from Turkey. Pathogens, 11(5), 500. https://doi.org/10.3390/pathogens11050500