1. Introduction
Blackleg disease (phoma stem canker) is an economically important disease in many oilseed rape-growing areas in the world [
1]. The causal agent of the disease is a two-species complex:
Leptosphaeria maculans (anamorph =
Plenodomus lingam) and
Leptosphaeria biglobosa (anamorph =
Plenodomus biglobosus) [
2,
3]. Both species coexist in the field. However, the ratio of
L. maculans to
L. biglobosa in a region is decisive for disease severity, so that
L. maculans is mainly responsible for significant yield losses [
4].
L. maculans was reported to be dominant in Germany and other western European countries [
1].
Having a complex lifecycle with two reproduction systems and different dispersal mechanisms,
L. maculans has a high evolutionary potential that allows it to readily adapt to new conditions, such as introducing new
R genes in its host plant [
5]. Generally, the fungus survives saprophytically on stubbles of a previous season by means of the sexual fruiting bodies, pseudothecia. Once mature, pseudothecia eject wind-borne ascospores, resulting in a primary infection of host plants. Ascospores germinate on cotyledons and young leaves. Hyphae grow biotrophically to form characteristic phoma lesions, mostly with asexual fruiting bodies, pycnidia. The latter contain pycnidiospores, which can be spread by rain splashes. Their spread may finally result in a secondary infection. The fungus grows symptomless from leaves through petioles to stems. Once in the stem base, the fungus turns necrotrophic and causes canker [
1,
6]. Additionally,
L. maculans can be seed-transmitted [
7]. Studies describing a high diversity in
L. maculans populations have repeatedly emphasized the importance of sexual reproduction for generating wide genetic variation [
8,
9]. However, despite its importance, sexual reproduction might not be dominant in some seasons. Such cases were reported in western Canada, where ascospores were not detected in the air in some years, although infection was severe. Thus, it was concluded that ascospores did not form the main inoculum. Instead, pycnidiospores represented the alternative main source of inoculum [
10].
L. maculans has two mating types,
MAT1-1 and
MAT1-2 [
11]. According to the random mating hypothesis, the mating type frequencies in a population should be 1:1 under random sexual outcrossing [
12]. Defining the idiomorph ratios in a
L. maculans population helps to determine the source of the primary inoculum, and thus the speed expected for the pathogen to evolve new races in a specific region.
Integrated management of phoma stem canker includes stubble management, crop rotation, applying fungicides and using resistant cultivars [
13,
14]. Two types of resistance are known: polygenic, non-race specific, quantitative resistance and major gene, race-specific, qualitative resistance [
1,
15]. The immune system in plants is described by a “Zigzag” model of several phases [
16]. Once
L. maculans interacts with the host, the fungus initially confronts extracellular pattern recognition receptors (PRRs) which recognize pathogen-associated molecular patterns (PAMPs), and this recognition results in PAMP-triggered immunity (PTI). The pathogen excretes specific effectors to suppress PTI. If the plant cannot recognize these effectors, an effector-triggered susceptibility (ETS) will be initiated. Otherwise, a specific recognition of effectors will activate an effector-triggered immunity (ETI) [
16]. Effector-specific recognition happens according to the gene-for-gene concept, in which each
Avr gene in the pathogen has a counterpart major
R gene in the host [
17,
18]. The classic perception of the recognition process depicts it as a direct interaction between a plant receptor protein and a pathogen avirulence protein [
18,
19]. However, a more contemporary point of view speculates that
R gene(s) in a plant monitor the occurrence of modulations of host cell components that are targeted by the pathogen to prepare the cell environment for the invasion [
20,
21]. Through the co-evolution of pathogens and their hosts, natural selection allows a pathogen population to modify their
Avr profile to generate effectors able to successfully suppress PTI [
16].
New emerged isolates have previously been categorized into pathogenicity groups, including
L. biglobosa, until Shoemaker and Brun [
2] provided the final taxonomic evidence to separate
L. biglobosa as a distinct species. Since then, there has been a considerable gain of knowledge about major blackleg resistance genes, which have also been incorporated into actual tester sets, allowing the definition of races (up to 2
n). Characterization of
L. maculans by their races was suggested by Balesdent et al. [
22]. This characterization relies on differential cotyledon reactions of tester lines to individual isolates of
L. maculans based on
Avr–
R gene interactions.
Avr–
R gene interactions in the
L. maculans-
B. napus pathosystem can be more complex than involving two genes. On the one hand, a redundant recognition of a single
Avr gene can be displayed by two major
R genes [
23]. Larkan et al. demonstrated that the avirulence gene
Avrlm1 encodes effectors that can trigger the two major
R genes
LepR3 and
Rlm1 [
24]. On the other hand, there are cases where two
Avr genes must act together to be able to trigger one
R gene, the so-called two-gene-for-one-gene interaction. For example,
Rlm10-mediated recognition can be triggered only if both
Avrlm10A and
Avrlm10B are present together [
25]. Additionally, it was reported that the functional allele
Avrlm7 masks the recognition of
Avrlm9 and
Avrlm3 due to an epistatic interaction effect [
26,
27]. In
L. maculans, fourteen
Avr genes have been identified so far. Eight of them were cloned:
Avrlm1-L3 [
28],
Avrlm2 [
29],
Avrlm3 [
26],
Avrlm4-7 [
30],
Avrlm5-9 [
27,
31],
Avrlm6 [
32],
Avrlm10 [
25] and
Avrlm11 [
33]. On the side of the host, in
B. napus, an even larger number of corresponding major
R genes were described, such as
Rlm1, Rlm2, Rlm3, Rlm4, Rlm5, Rlm6, Rlm7, Rlm8, Rlm9, Rlm10, Rlm11, RlmS (BLMR1.2), Rlm13, LepR1, LepR2, LepR3 (BLMR1.1) and
LepR4. Only three of them,
Rlm2, Rlm9 and
LepR3, were cloned [
24,
34,
35,
36,
37,
38,
39,
40].
Sowing a specific commercial cultivar harboring a major
R gene in a region over years results in high natural selection pressure. Hence, new
L. maculans races evolve that can overcome the introduced major
R gene. This is called a “boom and bust” cycle [
5]. The amplitude of a “boom and bust” cycle of a major
R gene differs among fungal phytopathogens. For
L. maculans, several studies have documented the potential lifespan of a major
R gene when intensively deployed under experimental conditions or at a commercial level. For example, Brun et al. reported that
Rlm6 turned ineffective after three growing seasons in field experiments [
41]. On a commercial level, breakdowns of major
R genes have been observed within three to five years after their introduction into the market in many oilseed rape-growing countries, such as the efficacy loss of
Rlm1 in France,
Rlm3 in western Canada and “sylvestris”-derived resistance, namely
Rlm1 and
LepR3, in Australia [
42,
43,
44].
Setting a strategic regional plan to rotate cultivars harboring major
R genes is essential to expand the efficacy longevity of major
R genes. The reason is that using qualitative resistance is only reasonable as long as the corresponding
Avr gene is dominant in the population. However, a reliable plan for major
R gene rotation requires regular updating of the
Avr profile of regional
L. maculans populations [
45]. Monitoring of
Avr gene frequencies is thus crucial for practical recommendations for farmers and breeders.
The last study that investigated
Avr gene frequencies in
L. maculans populations in Germany was based on samples collected in the growing seasons of 2011 and 2012 [
46]. Here, we aimed not only to update the
Avr profile of
L. maculans populations, but also to expand the range of the tested
Avr genes by including
Avrlm6, Avrlm11, AvrlepR1, AvrlepR2 and
AvrlepR3, which have not been investigated in Germany so far. In addition, special attention was given to the change in
Avrlm7 frequency, since
Rlm7 has been known as the most effective commercialized major
R gene in Germany in recent years. Additionally, this study aimed to check whether
L. maculans population mating types deviate from the hypothesized 1:1 ratio in northern and central Germany.
3. Discussion
This is the first study identifying the frequencies of
AvrlepR1,
AvrlepR2,
AvrlepR3,
Avrlm6 and
Avrlm11 in the
L. maculans population in German fields. Besides, it has updated the
Avr profiles of
Avrlm1,
Avrlm2,
Avrlm3,
Avrlm4 and
Avrlm7, which have not been investigated in Germany since 2012 [
46]. Cotyledon tests showed that 100% of the isolates were virulent on differential lines harboring
Rlm2. The absence of
Avrlm2 was also described in northern Germany in isolates sampled in 2011–2012 [
46]. Similar results have previously been found in France in 2000–2001 [
49]. However, this situation is different on other continents. For example, in western Canada, a survey on samples collected in 2012–2014 showed that
Avrlm2 reached 80% [
50]. All our sampled isolates were virulent on the
Rlm9 differential. This can be explained by the use of the
Brassica napus trap cultivar NK-Bravour that carries the
Rlm9 R gene. This led to a preselection of
avrlm9-harboring isolates, whereas
Avrlm9-harboring isolates were counter-selected.
Epistatic effects, as a mechanism for evading recognition, were reported by
Avrlm7 toward
Avrlm3 [
26] and
Avrlm9 [
27]. Indeed, the results of our study supported that the presence of the functional
Avrlm7 masked the recognition of
Avrlm3. Investigations of epistatic mechanisms of
Avrlm7 toward
Avrlm3 and
Avrlm9 revealed that this suppression was caused by neither stopping the expression of
Avrlm3 and
Avrlm9, nor a physical interaction of the
Avr effector proteins of
Avrlm3,
Avrlm9 and
Avrlm4–
7 [
27,
51]. Using protein structure approaches, it was demonstrated that the three effector proteins of these genes belong to a new family of effectors, called
Leptosphaeria AviRulence-Supressing effectors (LARS). LARS are structurally analogue effectors that differ in their amino acid identities, although they share common targets in the plants [
52].
In our study, several compatible interactions on the
Rlm3 differential lines were detected among isolates lacking the functional
Avrlm7. This indicates that although
Avrlm7 masks the recognition of
Avrlm3, its absence does not mean that the pathogen has no other means to avoid recognition. A variety of virulence-gain mechanisms in plant pathogens have been described, such as deletion of the
Avr gene, point mutations that allow the pathogen to avoid recognition despite the presence of the
Avr gene, amino acid substitution and masking the
Avr protein through another
Avr protein [
26,
47,
53]. Gene silencing of
Avrlm3 proved that this
Avr gene is crucial in
L. maculans pathogenicity and has an important effect on its lifecycle in
B. napus [
51]. Thus, deletion of the gene can be ruled out by elucidating the virulence of
avrlm7 isolates on
Rlm3. Plissonneau et al. explained the virulence of
avrlm7 isolates toward
Rlm3 by the high allelic polymorphism of
Avrlm3, which allows a high level of possible protein isoforms. Therefore, it can be speculated that the alternative mechanism used by the pathogen when the epistatic effect disappears is to substitute an amino acid in the effector protein to allow a so-called “camouflage” type of escaping recognition [
51]. Setting strategies for
R gene management based on restoring the efficacy of
Rlm9 and
Rlm3 by losing the functional
Avrlm7 could mislead farmers into reducing caution in
Rlm7 deployment.
Our results showed that major
R genes
Rlm1, Rlm3, Rlm4 and
LepR3 are not able to provide sufficient resistance against phoma stem canker in German fields. In the explored regions,
Avrlm4 isolate frequencies ranged from 3% to 13%. This is quite close to the rate described in a large-scale survey throughout France in 2000–2001 (0–19.5%) [
49].
Avrlm4-harboring isolates turn virulent when glycine at position 120 in the
Avr protein is substituted with arginin [
54], while
Avrlm1 and
AvrlepR3, which are suspected to be alleles of the gene
Avrlm1-L3, turn virulent mainly due to deletion of the whole gene [
55,
56]. Our results showed that
Avrlm1 frequency ranged from 2% to 19%. A rapid adaptation of the
L. maculans population toward
Rlm1 was noticed in France, where the rate of
Avrlm1-harboring isolates made up 83% of the population in 1997–1998 and decreased dramatically to less than 13% in 1999–2000 [
42]. Similarly, in Canada,
Avrlm1 frequency did not exceed 5% [
45]. In Australia, the efficacy of
Rlm1 resistance in cultivar Surpass 400 notably decreased within three years after commercial release [
23], as well as the efficacy of
LepR3. This is explained by the fact that
Avrlm1 has dual specificity and can trigger both
R genes:
Rlm1 and
LepR3 [
24]. It can be concluded that wherever
Rlm1 efficacy is broken,
LepR3’s lifespan is shorter than that of other
R genes, in which the corresponding
Avr gene does not have dual specificity.
We identified a dramatic increase in the frequency of
avrlm7 isolates compared with the 2011–2012 season. The frequency of
avrlm7 isolates increased within five to seven years from 0.9% to 17.6% in fields located in central and northern Germany, and the highest presence of them was in Peine, where the frequency reached 28%. Winter and Koopmann stated that
Rlm7 was the only still effective major
R gene used in commercial cultivars in Germany [
46]. The potential speed of
Rlm7 breakdown was experimentally tested by applying maximum pressure on the
L. maculans population through sowing cultivars with
Rlm7 for four years, without applying crop rotation or ploughing in the crop debris. Under these experimental conditions, the results revealed that
avrlm7 isolates frequency increased from 0 to 36% within 4 years [
54]. In 2006, a study at the European level showed that the
Avrlm7 frequency was 100% in France, Germany, Sweden and Poland [
57]. However, the intense deployment of
Rlm7 resulted in a rapid reduction of
Avrlm7 not only in Germany, but also in France. There,
Rlm7 was introduced commercially in 2004. By 2013, the frequency of
Avrlm7 isolates reached an average of 19.5% and a maximum of 45%, depending on the region [
58]. In contrast,
Rlm7 is still effective in Canada, with an
Avrlm7 frequency exceeding 98% in 2018–2019 [
45]. Hence, the present study calls for a more cautious deployment of
Rlm7, especially in Europe.
Some major
R genes in oilseed rape were derived from related
Brassica species. For instance, while
Rlm11,
LepR1, LepR2 and
LepR3 were introduced into
B. napus from
B. rapa [
33,
34,
35],
Rlm6 originated from
B. juncea [
59]. Our survey provided strong evidence that
LepR1 is the most effective major
R gene in Germany, with 100% frequency of the isolates harboring
AvrlepR1 in all regions investigated, except in Peine, where 2% of the isolates were virulent on
LepR1. Although the proportion of the virulent isolates toward
LepR1 was very low, and such isolates were found only in one region, the former observation of the rapid decline in efficacy of
Rlm7 within 5–7 years should prompt to take measures to preserve the efficacy of
LepR1.
In France,
AvrlepR1, AvrlepR2, Avrlm6 and
Avrlm11 frequencies in the
L. maculans population were shown to still be high enough to control the disease in the field [
60]. This is in line with our findings in Germany. A significant constraint in the targeted use of
R genes in the field is a lack of knowledge about their presence in several commercial cultivars registered in Germany.
LepR2, which is believed to be the same gene or an allelic form of
RlmS, was reported in cultivars recently registered in France and Germany [
61]. Considering the frequency of
avrlepR2 in Germany of 9–37%, as shown in our study, an annual monitoring seems necessary in regions where cultivars harboring
LepR2 are grown. This may help to avoid a situation such as that in Canada, where
AvrlepR1,
AvrlepR2 and
AvrlepR3 occur already at low frequencies of ca. 19%, 5% and 5–28%, respectively [
45].
The major mechanism of virulence gain of
Avrlm6 is a deletion of the gene [
47]. Van de Wouw et al. studied isolates collected in Australia over two decades (from 1987 to 2017) and observed fluctuating frequencies of
Avrlm6 from 21% to 80% depending on the year. This behavior appeared to be independent from the commercial introduction of
Rlm6 in Australia in 2010 [
62]. Hence, the study speculated that
Avrlm6 frequency might also be influenced by the intensive deployment of other major
R genes in Australia [
62]. In France,
Rlm6 was not introduced commercially after the French National Institute for Agricultural Research (INRA) decided to keep it as a research tool [
41]. As a result,
Avrlm6 frequency was 100% in France [
49], similar to Canada, where
Avrlm6 frequency exceeds 98% [
45]. In our study, a fluctuation in frequency between 88% and 100% depending on the region was observed, however the use of
Rlm6 in Germany is not clear.
Avrlm11 has been reported to occur in high frequencies in many regions of the world. For instance, its frequency was >95% in France and 100% in Canada [
33,
45]. In our study, the rate of isolates harboring
Avrlm11 ranged from 66% to 95%. Despite the high frequency of
Avrlm11, it was found to be located on a dispensable mini-chromosome of
L. maculans that was occasionally lost by meiosis. Thus, the loss of
Avrlm11 is related to the loss of this mini-chromosome in
L. maculans. [
33]. In spite of its dispensability, this chromosome turned out to be influential for the viability of ascospores, and thus, its loss has fitness costs [
33].
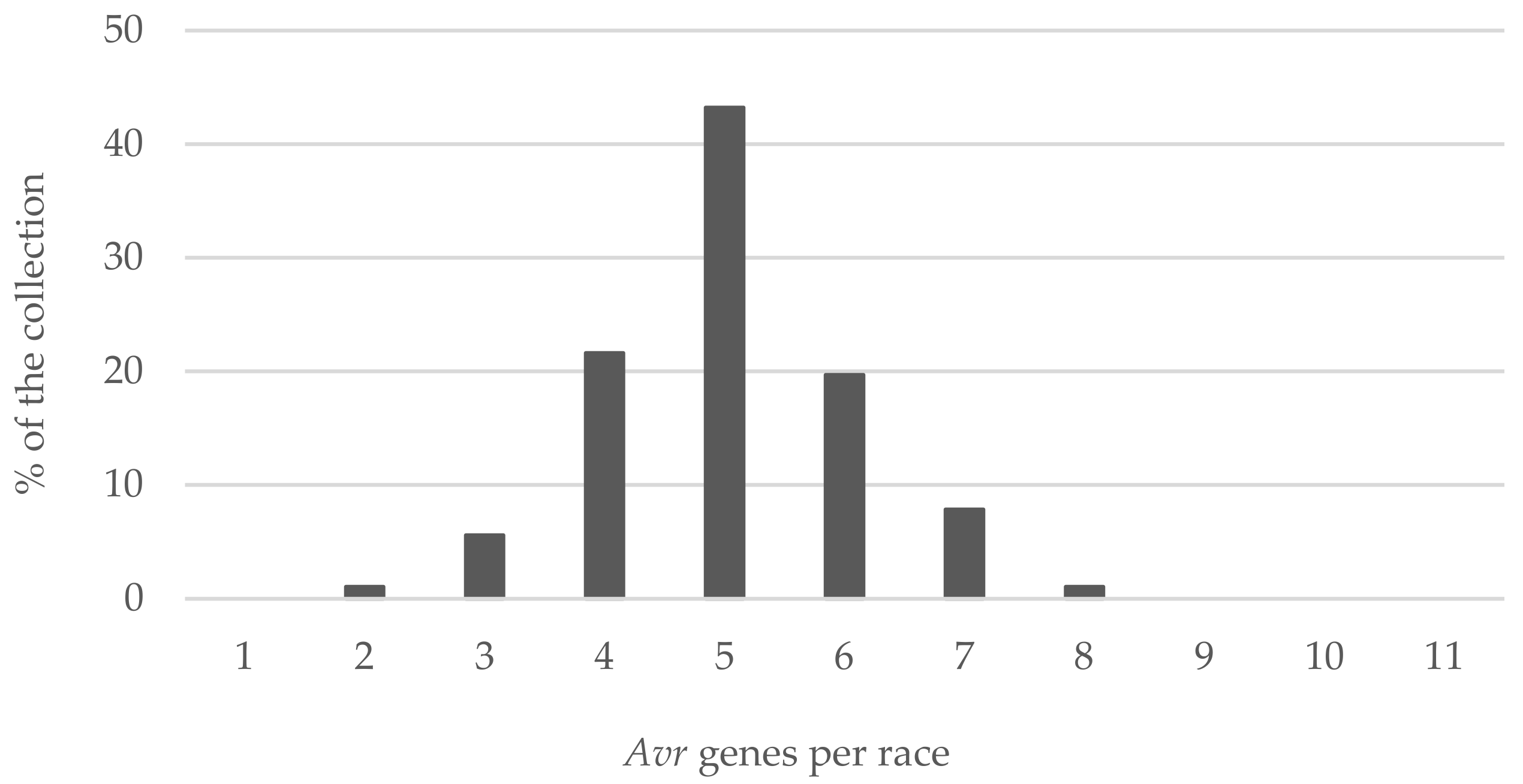
The race structure in our study was described based on the phenotypic analysis of eight
Avr genes and the PCR assays for
Avrlm6 and
Avrlm11. Assuming each of the 10 tested
Avr genes has a minimum of 2 alleles, the theoretical number of combinations would be 1024 (i.e., 2
10). This shows the theoretical potential of the pathogen to break resistance. In our study, we were able to determine 52 races, the majority of which had a complexity of five
Avr genes. Race complexity depicts the range of the available effective major
R genes for setting management strategies for resistant cultivar rotation in a region [
45]. In general, we found, according to our isolate collection, that the major
R genes that provide sufficient resistance in German oilseed rape fields are
LepR1, LepR2, Rlm6, Rlm11 and
Rlm7, while 61.3% of the isolates can equally infect cultivars harboring
Rlm1, Rlm2 and
Rlm4. This is consistent with our previous investigation in Germany [
46]. Combining quantitative resistance with race-specific resistance can be important to expand the durability of major
R genes. It was demonstrated that
Rlm6 in a cultivar with a quantitative resistance background preserved its efficacy two years longer than in a cultivar with a susceptible background [
41].
Pathogens with a high evolutionary potential are expected to overcome genetic resistance more rapidly [
5]. It is believed that there are two important factors that enhance the evolutionary potential of
L. maculans: first, the mixed reproduction systems, and second, the dispersal mechanisms. The sexual reproduction is a milestone for producing variations in the population, while the wind dispersal of spores spread the evolving races effectively over several kilometers [
54,
63]. However, in seasons when the environmental conditions do not favor sexual reproduction, asexual reproduction would be dominant, and thus, the ratio of idiomorphs might change. For example, in some Canadian regions, samples collected in 2010 and 2015 showed that the mating type
MAT1-2 was significantly more dominant than
MAT1-1 [
64]. In our study, we showed that the idiomorphs of
L. maculans in Germany did not significantly deviate from the ratio 1:1. This is consistent with the ratio observed in France in isolates collected in 2000–2003 [
65] and in Canada in samples from 2011–2014 [
64], which indicates similar lifecycles of this pathogen in different regions in the world and a central role of the sexual outcrossing in forming the population of the pathogen.
Overall, we comprehensively compared the effectiveness of major
R genes in different countries. We found similarities in the
R gene efficacies throughout Germany’s neighboring countries, but increasingly significant differences the further those countries were from our study region. This may be related to the fact that the pathogen mainly spreads through wind-borne ascospores that can fly up to several kilometers and stay viable for six weeks [
1]. We therefore recommend the consideration of geographical factors for the successful management of blackleg disease.




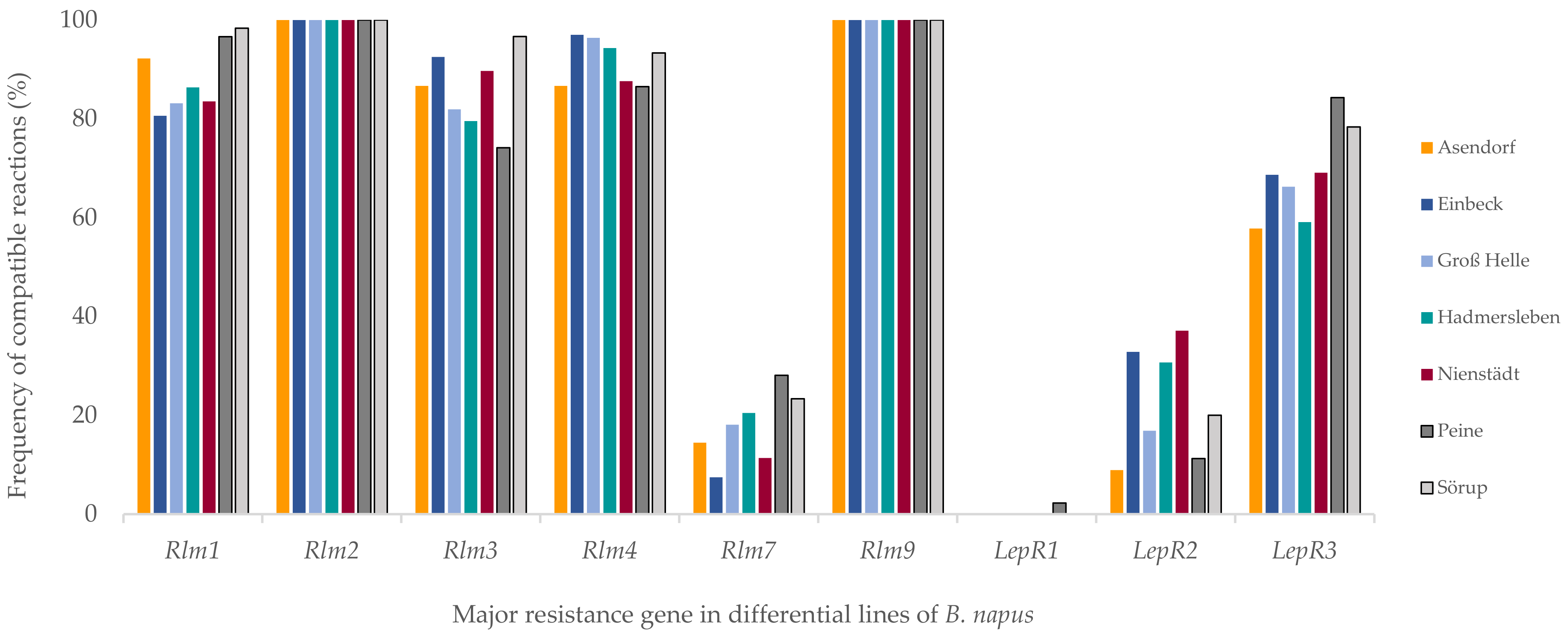



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}