
Salmonella Enteritidis Subunit Vaccine Candidate Based on SseB Protein Co-Delivered with Simvastatin as Adjuvant

 and
and
Abstract
:1. Introduction
2. Results
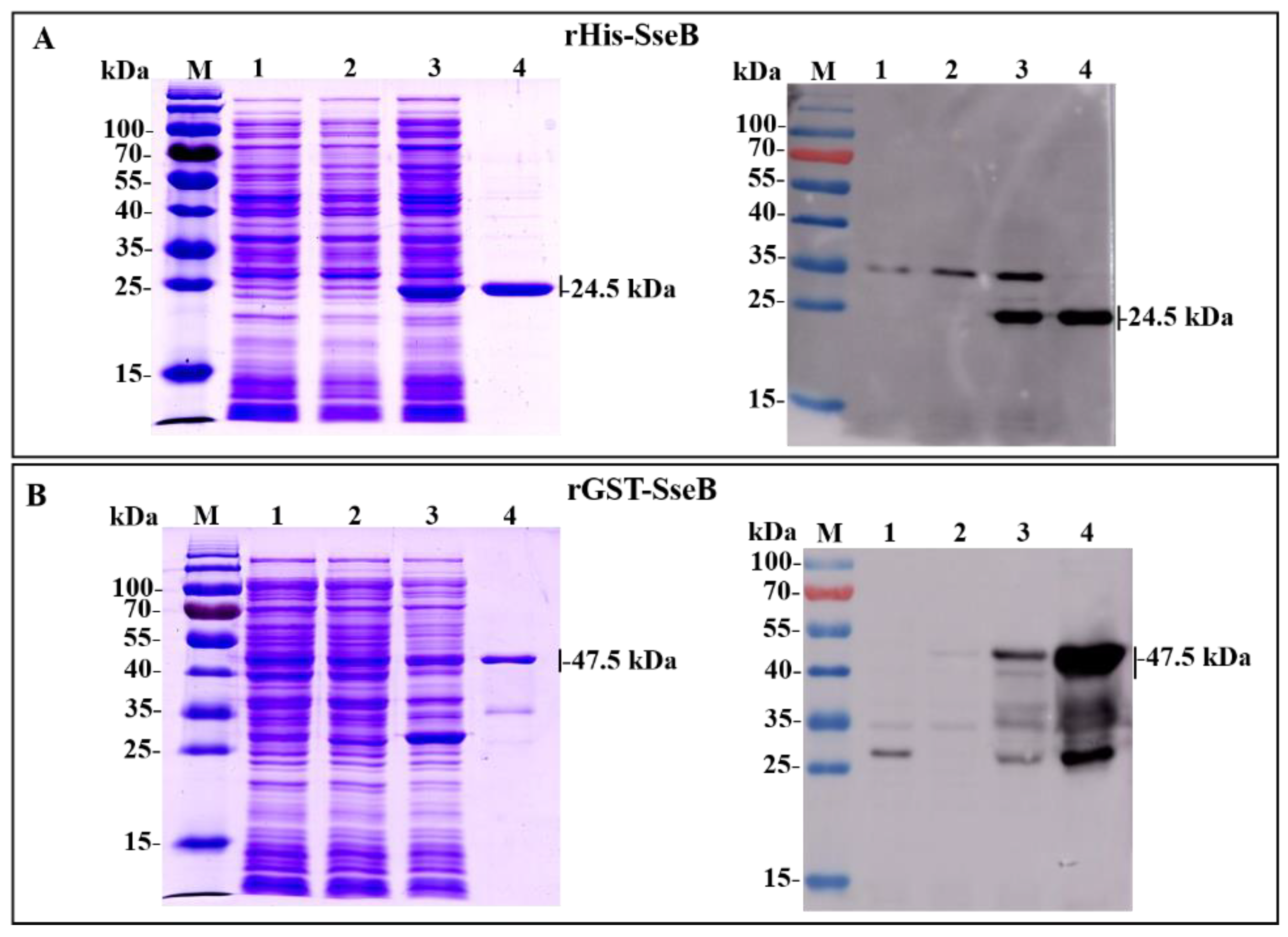
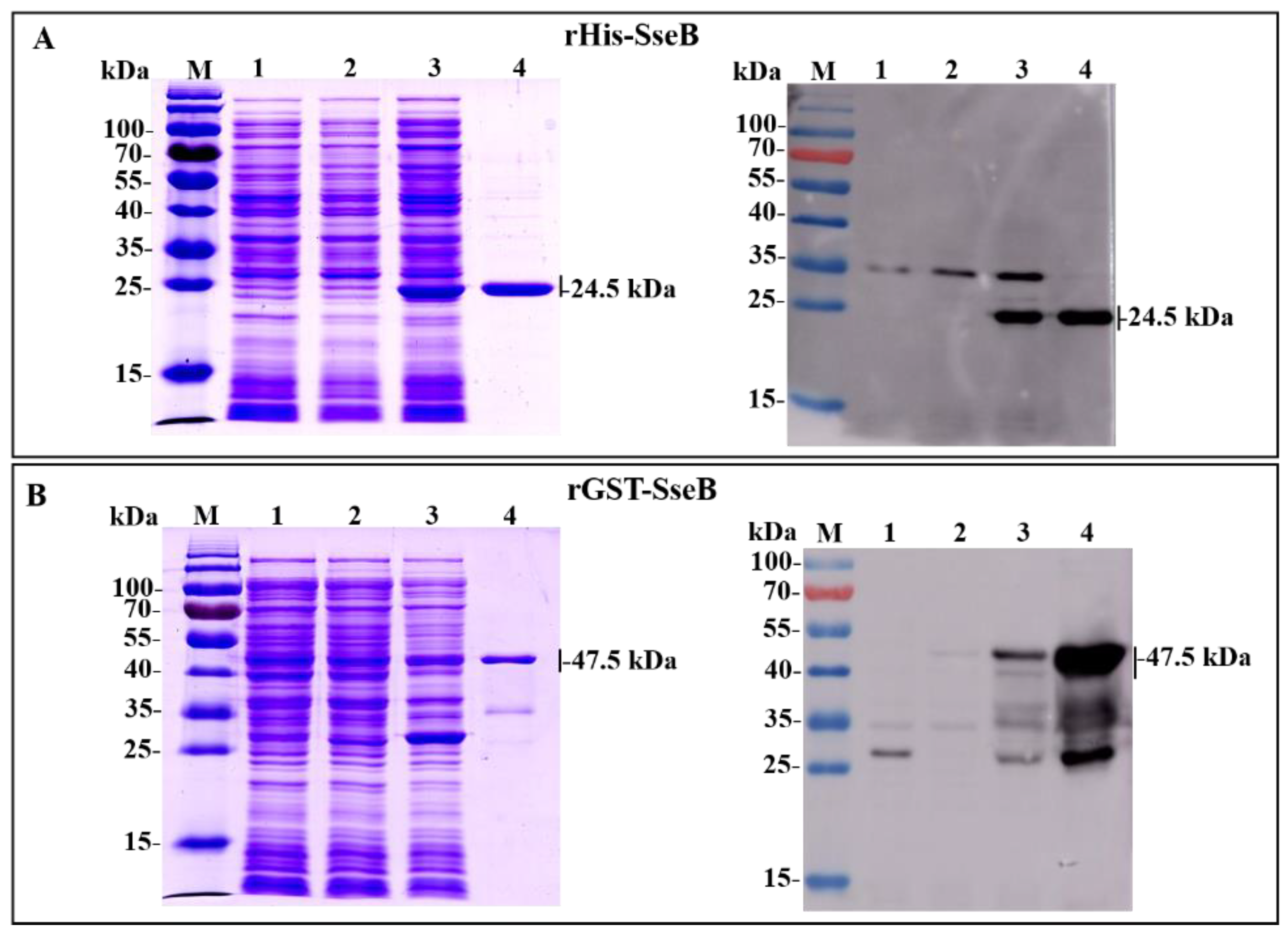
2.1. Expression, Purification, and Identification of Recombinant Proteins rHis-SseB and rGST-SseB
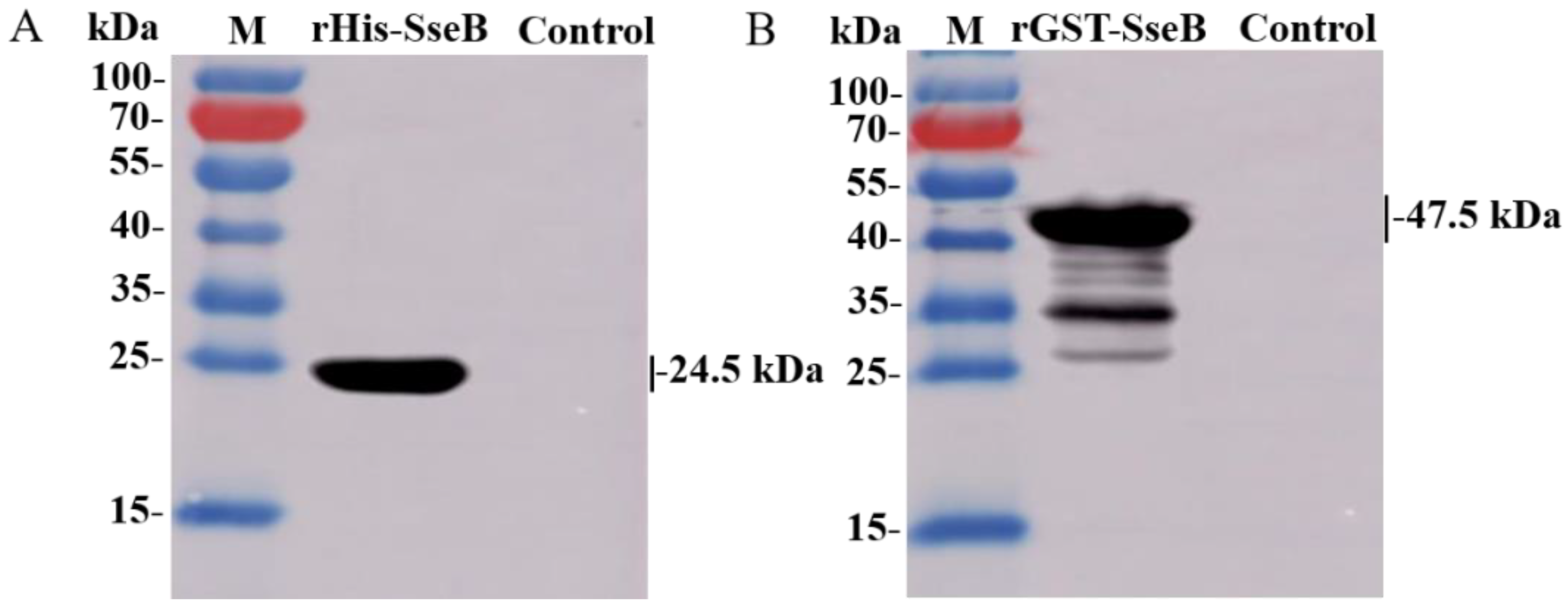
2.2. Immunoreactivity of Recombinant Proteins rHis-SseB and rGST-SseB
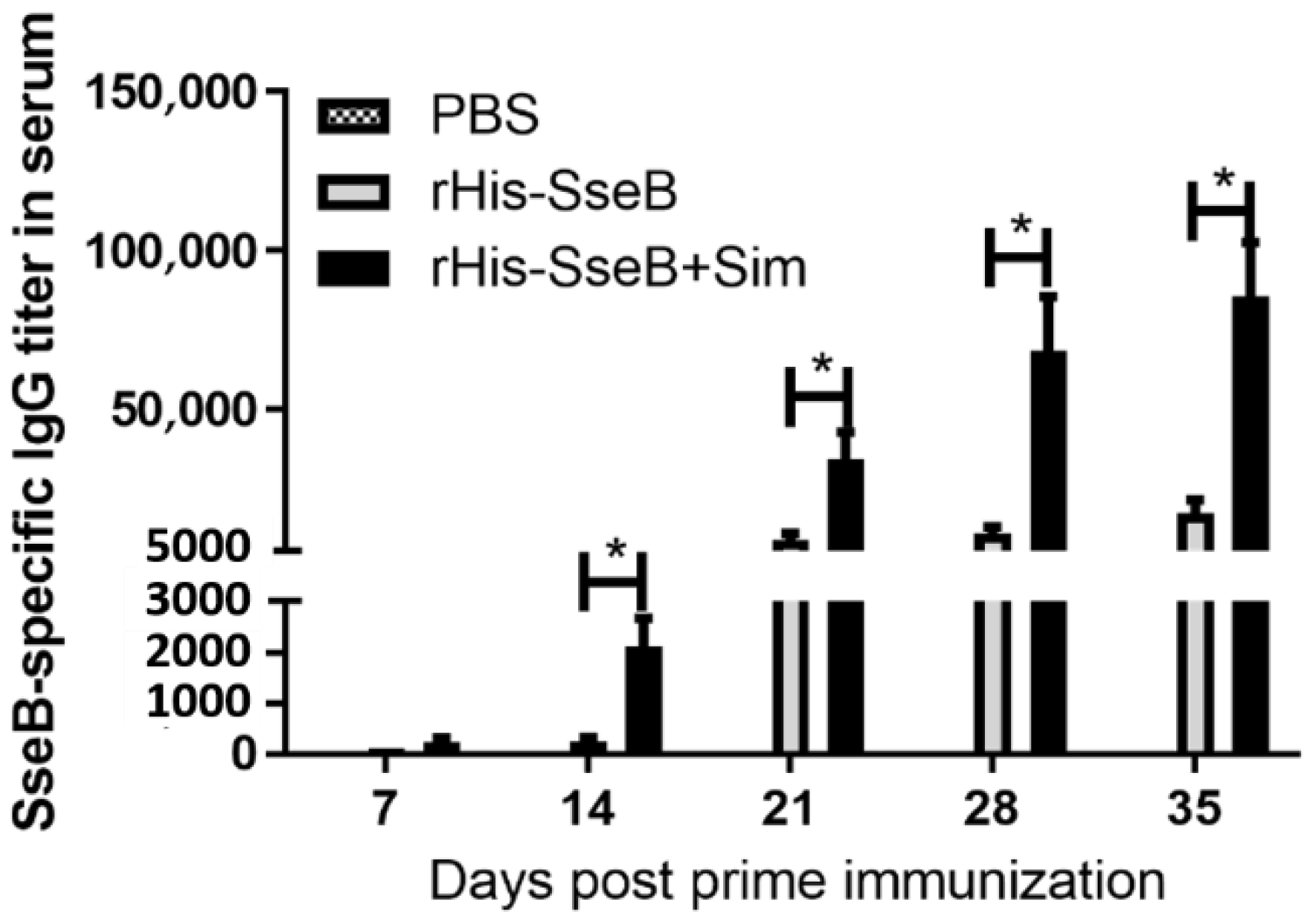
2.3. Simvastatin-Enhanced Antibodies Induced by rHis-SseB in Serum
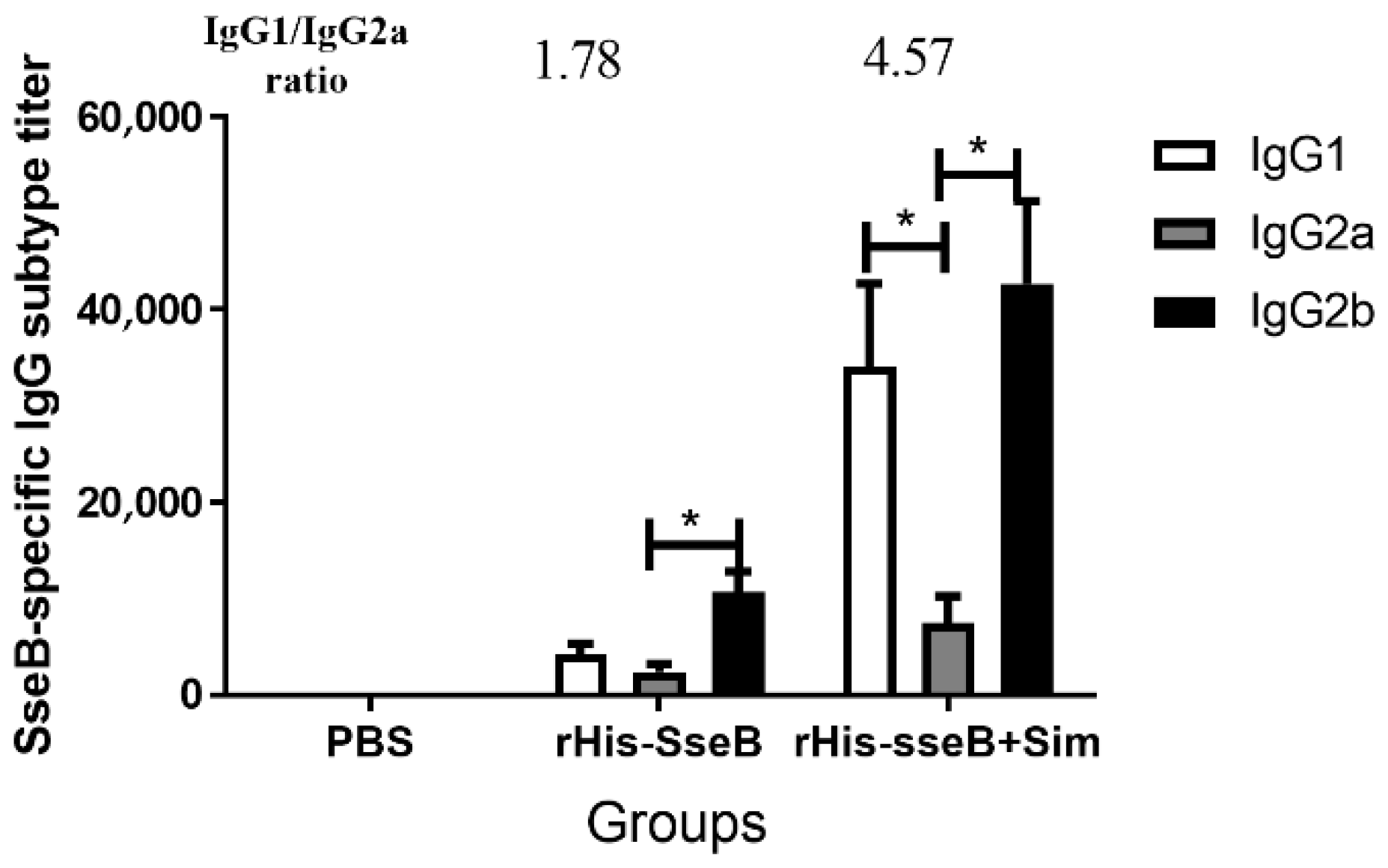
2.4. IgG Subtype Induced by rHis-SseB in Serum
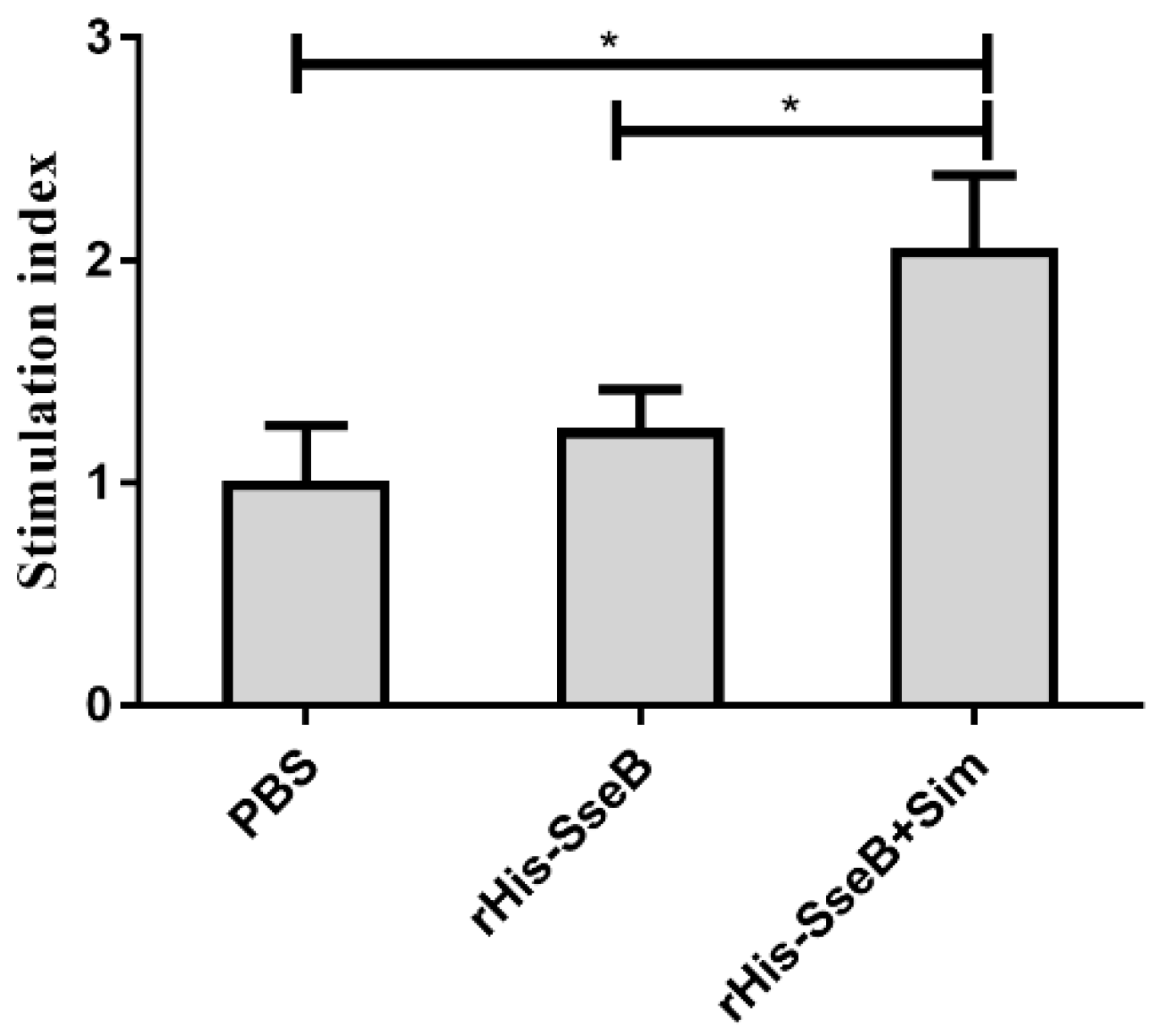
2.5. Simvastatin-Enhanced Cellular Immune Responses Induced by rHis-SseB
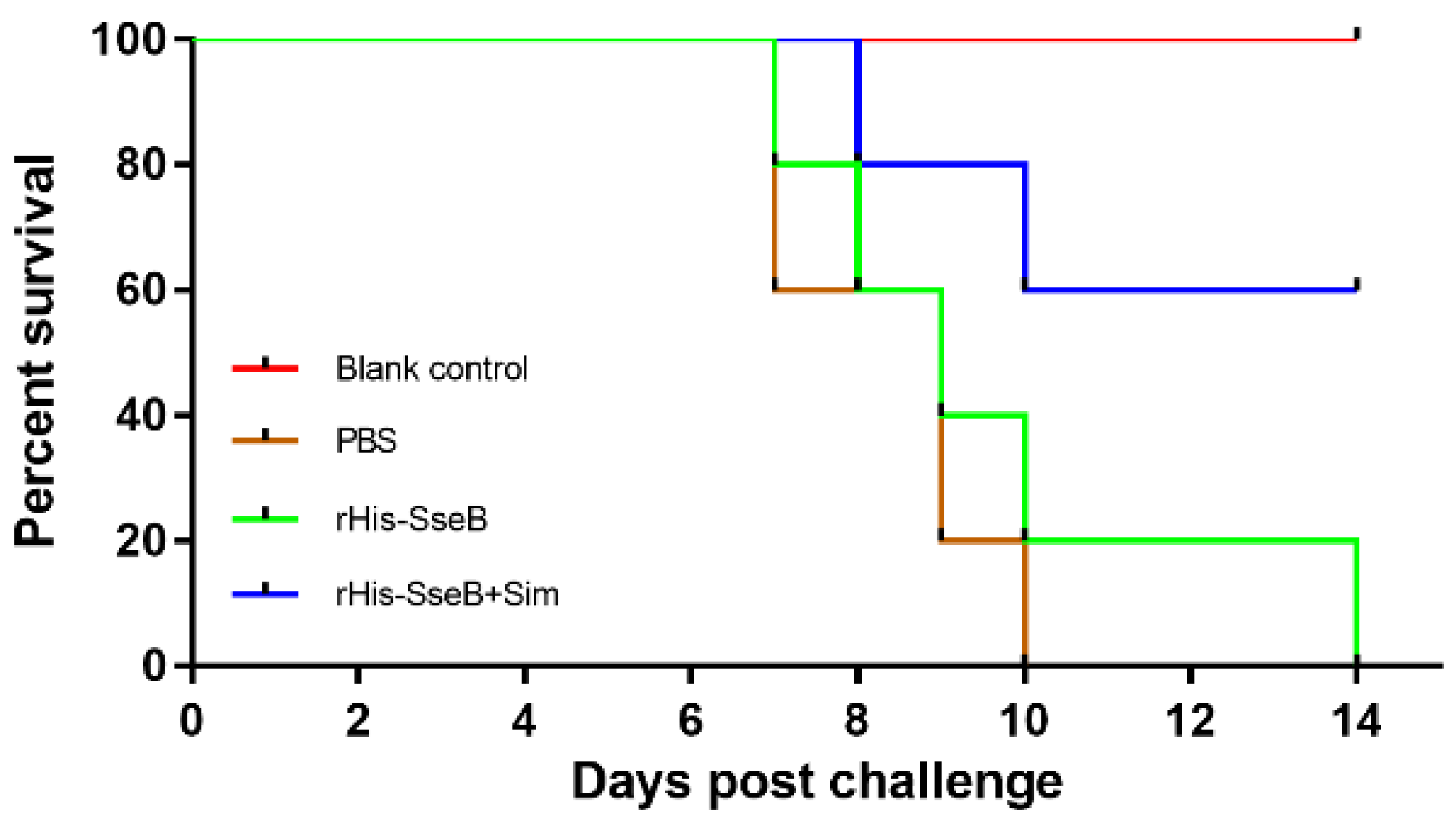
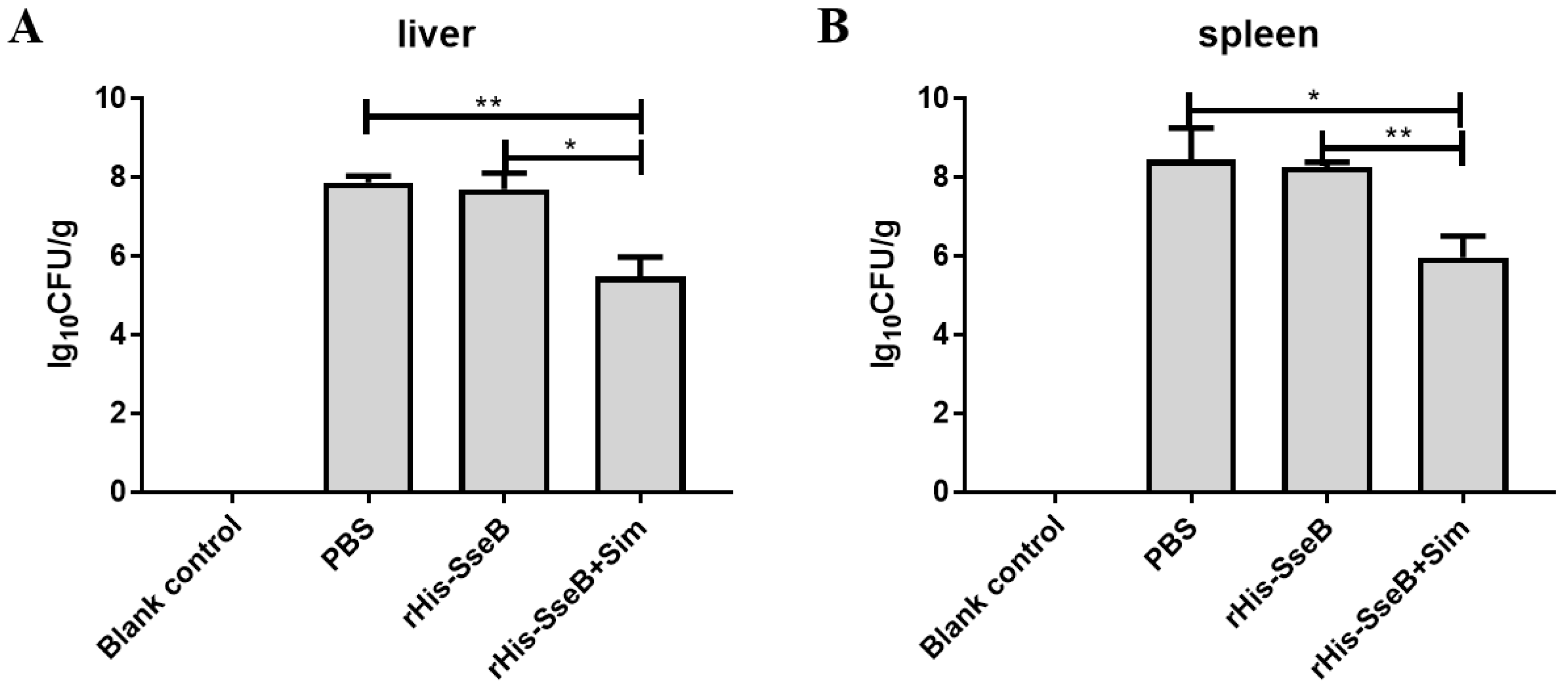
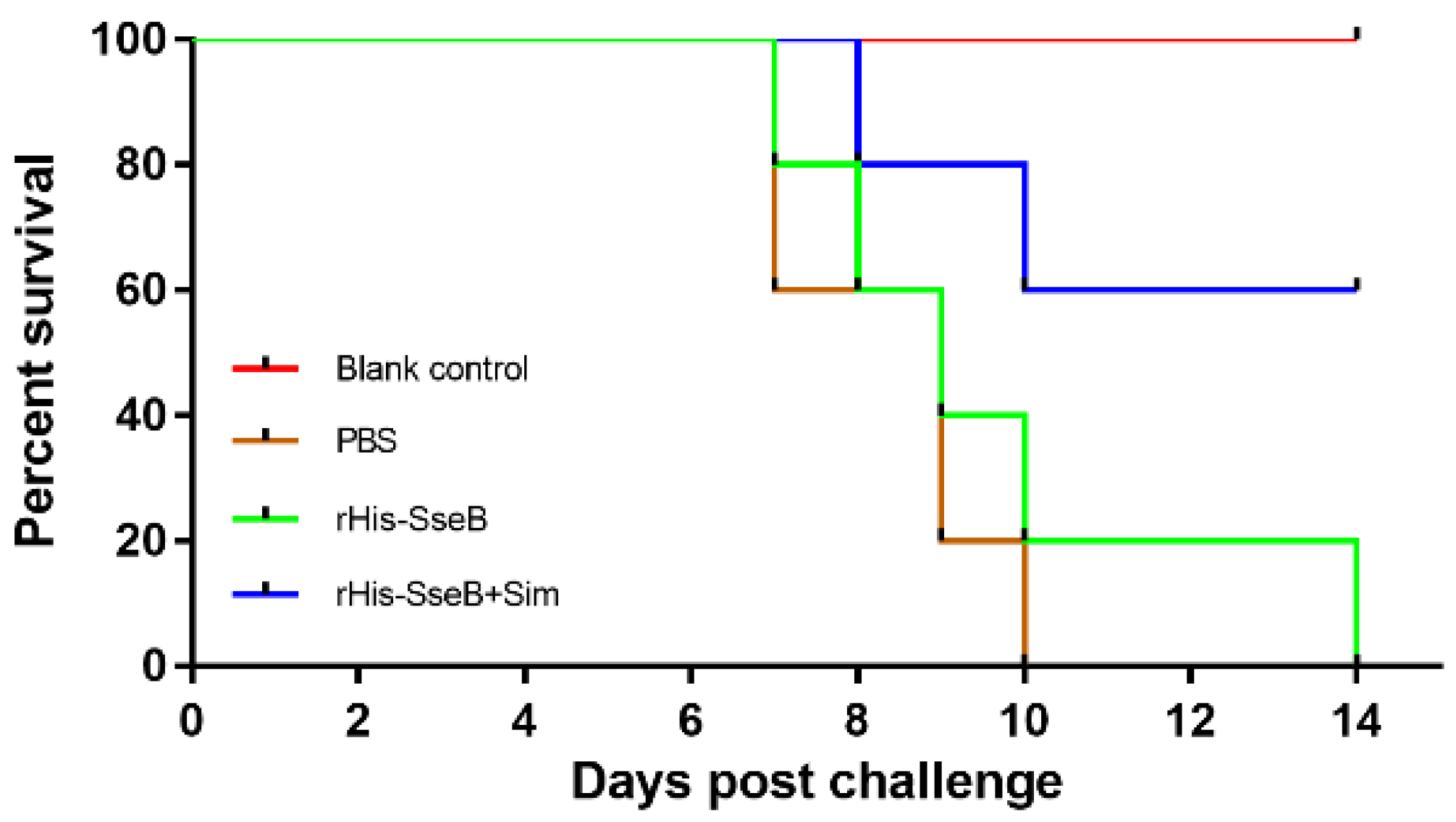
2.6. Candidate Vaccine Protected Mice against Oral Challenge with Wild-type S. Enteritidis
3. Discussion
4. Materials and Methods
4.1. Mice and Ethics Statement
4.2. Construction of Recombinant Expression Plasmid
4.3. Expression and Purification of Recombinant Proteins rHis-SseB and rGST-SseB
4.4. Western Blotting
4.5. Mouse Vaccination
4.6. Detection of Anti-SseB Antibodies by Indirect ELISA
4.7. Lymphocyte Proliferation Assay
4.8. Immune Protection Assessment
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, X.; Biswas, S.; Gu, G.; Elbediwi, M.; Li, Y.; Yue, M. Characterization of multidrug resistance patterns of emerging Salmonella enterica serovar Rissen along the food chain in China. Antibiotics 2020, 9, 660. [Google Scholar] [CrossRef]
- Kuang, X.; Hao, H.; Dai, M.; Wang, Y.; Ahmad, I.; Liu, Z.; Zonghui, Y. Serotypes and antimicrobial susceptibility of Salmonella spp. isolated from farm animals in China. Front. Microbiol. 2015, 6, 602. [Google Scholar] [CrossRef] [Green Version]
- Ehuwa, O.; Jaiswal, A.K.; Jaiswal, S. Salmonella, food safety and food handling practices. Foods 2021, 10, 907. [Google Scholar] [CrossRef] [PubMed]
- Kirk, M.D.; Pires, S.M.; Black, R.E.; Caipo, M.; Crump, J.A.; Devleesschauwer, B.; Dopfer, D.; Fazil, A.; Fischer-Walker, C.L.; Hald, T.; et al. World health organization estimates of the global and regional disease burden of 22 foodborne bacterial, protozoal, and viral diseases, 2010: A data synthesis. PLoS Med. 2015, 12, e1001921. [Google Scholar]
- Cai, Y.Q.; Tao, J.; Jiao, Y.; Fei, X.; Zhou, L.; Wang, Y.; Zheng, H.J.; Pan, Z.M.; Jiao, X.N. Phenotypic characteristics and genotypic correlation between Salmonella isolates from a slaughterhouse and retail markets in Yangzhou, China. Int. J. Food Microbiol. 2016, 222, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Fei, X.; He, X.; Guo, R.X.; Yin, C.; Geng, H.P.; Wu, K.Y.; Yin, K.Q.; Geng, S.Z.; Pan, Z.M.; Li, Q.C.; et al. Analysis of prevalence and CRISPR typing reveals persistent antimicrobial-resistant Salmonella infection across chicken breeder farm production stages. Food Control 2017, 77, 102–109. [Google Scholar] [CrossRef]
- Klemm, E.J.; Gkrania-Klotsas, E.; Hadfield, J.; Forbester, J.L.; Harris, S.R.; Hale, C.; Heath, J.N.; Wileman, T.; Clare, S.; Kane, L.; et al. Emergence of host-adapted Salmonella Enteritidis through rapid evolution in an immunocompromised host. Nat. Microbiol. 2016, 1, 15023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patrick, M.E.; Adcock, P.M.; Gomez, T.M.; Altekruse, S.F.; Holland, B.H.; Tauxe, R.V.; Swerdlow, D.L. Salmonella enteritidis infections, United States, 1985–1999. Emerg. Infect. Dis. 2004, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S. Pathogenicity of Salmonella enteritidis in poultry. Int. J. Food Microbiol. 1994, 21, 89–105. [Google Scholar] [CrossRef]
- Lin, Z.J.; Tang, P.P.; Jiao, Y.; Kang, X.L.; Li, Q.C.; Xu, X.L.; Sun, J.; Pan, Z.M.; Jiao, X.A. Immunogenicity and protective efficacy of a Salmonella Enteritidis sptP mutant as a live attenuated vaccine candidate. BMC Vet. Res. 2017, 13, 194. [Google Scholar] [CrossRef] [PubMed]
- Babu, U.; Scott, M.; Myers, M.J.; Okamura, M.; Gaines, D.; Yancy, H.F.; Lillehoj, H.; Heckert, R.A.; Raybourne, R.B. Effects of live attenuated and killed Salmonella vaccine on T-lymphocyte mediated immunity in laying hens. Vet. Immunol. Immunopathol. 2003, 91, 39–44. [Google Scholar] [CrossRef]
- Liu, H.F.; Chen, L.P.; Wang, X.M.; Si, W.; Wang, H.N.; Wang, C.L.; Liu, S.G.; Li, G.X. Decrease of colonization in the chicks’ cecum and internal organs of Salmonella enterica serovar Pullorum by deletion of cpdB by Red system. Microb. Pathog. 2015, 80, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Penha, R.A.C.; de Paiva, J.B.; Arguello, Y.M.S.; da Silva, M.D.; Gardin, Y.; Resende, F.; Berchieri, A.; Sesti, L. Efficacy of several vaccination programmes in commercial layer and broiler breeder hens against experimental challenge with Salmonella enterica serovar Enteritidis. Avian Pathol. 2009, 38, 367–375. [Google Scholar]
- Lee, J.H. Protection against Salmonella typhimurium, Salmonella gallinarum, and Salmonella enteritidis infection in layer chickens conferred by a live attenuated Salmonella typhimurium strain. Immune Netw. 2015, 15, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Gast, R.K. Serotype-specific and serotype-independent strategies for preharvest control of food-borne Salmonella in poultry. Avian Dis. 2007, 51, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Barrow, P.A. Salmonella infections: Immune and non-immune protection with vaccines. Avian Pathol. 2007, 36, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Desin, T.S.; Koster, W.; Potter, A.A. Salmonella vaccines in poultry: Past, present and future. Expert Rev. Vaccines 2013, 12, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Meenakshi, M.; Bakshi, C.S.; Butchaiah, G.; Bansal, M.P.; Siddiqui, M.Z.; Singh, V.P. Adjuvanted outer membrane protein vaccine protects poultry against infection with Salmonella enteritidis. Vet. Res. Commun. 1999, 23, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Toyota-Hanatani, Y.; Kyoumoto, Y.; Baba, E.; Ekawa, T.; Ohta, H.; Tani, H.; Sasai, K. Importance of subunit vaccine antigen of major FliC antigenic site of Salmonella enteritidis II: A challenge trial. Vaccine 2009, 27, 1680–1684. [Google Scholar] [CrossRef]
- De Buck, J.; Van Immerseel, F.; Haesebrouck, F.; Ducatelle, R. Protection of laying hens against Salmonella Enteritidis by immunization with type 1 fimbriae. Vet. Microbiol. 2005, 105, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Wisner, A.L.; Desin, T.S.; Lam, P.K.; Berberov, E.; Mickael, C.S.; Townsend, H.G.; Potter, A.A.; Koster, W. Immunization of chickens with Salmonella enterica subspecies enterica serovar Enteritidis pathogenicity island-2 proteins. Vet. Microbiol. 2011, 153, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.J.; Jones, C.; Blohmke, C.J.; Darton, T.C.; Goudet, A.; Sergeant, R.; Maillere, B.; Pollard, A.J.; Altmann, D.M.; Boyton, R.J. The serodominant secreted effector protein of Salmonella, SseB, is a strong CD4 antigen containing an immunodominant epitope presented by diverse HLA class II alleles. Immunology 2014, 143, 438–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutwiri, G.; Gerdts, V.; van Drunen Littel-van den Hurk, S.; Auray, G.; Eng, N.; Garlapati, S.; Babiuk, L.A.; Potter, A. Combination adjuvants: The next generation of adjuvants? Expert Rev. Vaccines 2011, 10, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Xie, Y.; Yu, Z.; Xiao, H.; Jiang, G.; Zhou, X.; Yang, Y.; Li, X.; Zhao, M.; Li, L.; et al. The mevalonate pathway is a druggable target for vaccine adjuvant discovery. Cell 2018, 175, 1059–1073. [Google Scholar] [CrossRef] [Green Version]
- Havelaar, A.H.; Kirk, M.D.; Torgerson, P.R.; Gibb, H.J.; Hald, T.; Lake, R.J.; Praet, N.; Bellinger, D.C.; De Silva, N.R.; Gargouri, N.; et al. World health organization global estimates and regional comparisons of the burden of foodborne disease in 2010. PLoS Med. 2015, 12, e1001923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, Y.; Xia, Z.; Zhou, X.; Guo, Y.; Guo, R.; Kang, X.; Wu, K.; Sun, J.; Xu, X.; Jiao, X.; et al. Signature-tagged mutagenesis screening revealed the role of lipopolysaccharide biosynthesis gene rfbH in smooth-to-rough transition in Salmonella Enteritidis. Microbiol. Res. 2018, 212–213, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Saenz, L.; Guzman, M.; Vidal, S.; Caruffo, M.; Siel, D.; Zayas, C.; Paredes, R.; Valenzuela, C.; Hidalgo, H.; Perez, O.; et al. Efficacy of multivalent, cochleate-based vaccine against Salmonella Infantis, S. Enteritidis and S. Typhimurium in laying hens. Vaccines 2022, 10, 226. [Google Scholar] [CrossRef] [PubMed]
- Van Immerseel, F.; Methner, U.; Rychlik, I.; Nagy, B.; Velge, P.; Martin, G.; Foster, N.; Ducatelle, R.; Barrow, P.A. Vaccination and early protection against non-host-specific Salmonella serotypes in poultry: Exploitation of innate immunity and microbial activity. Epidemiol. Infect. 2005, 133, 959–978. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Whalen, B.J.; Tirabassi, R.S.; Selin, L.K.; Levchenko, T.S.; Torchilin, V.P.; Kislauskis, E.H.; Guberski, D.L. A DNA vaccine prime followed by a liposome-encapsulated protein boost confers enhanced mucosal immune responses and protection. J. Immunol. 2008, 180, 6159–6167. [Google Scholar] [CrossRef]
- Mastroeni, P.; Villarreal-Ramos, B.; Demarco de Hormaeche, R.; Hormaeche, C.E. Delayed (footpad) hypersensitivity and Arthus reactivity using protein-rich antigens and LPS in mice immunized with live attenuated aroA Salmonella vaccines. Microb. Pathog. 1993, 14, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wolfenden, A.; Mandal, R.K.; Faulkner, O.; Hargis, B.; Kwon, Y.M.; Bielke, L. Evaluation of recombinant Salmonella vaccines to provide cross-serovar and cross-serogroup protection. Poult. Sci. 2017, 96, 4352–4360. [Google Scholar] [CrossRef] [PubMed]
- Rollenhagen, C.; Sorensen, M.; Rizos, K.; Hurvitz, R.; Bumann, D. Antigen selection based on expression levels during infection facilitates vaccine development for an intracellular pathogen. Proc. Natl. Acad. Sci. USA 2004, 101, 8739–8744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.J.; Liang, L.; Juarez, S.; Nanton, M.R.; Gondwe, E.N.; Msefula, C.L.; Kayala, M.A.; Necchi, F.; Heath, J.N.; Hart, P.; et al. Identification of a common immune signature in murine and human systemic Salmonellosis. Proc. Natl. Acad. Sci. USA 2012, 109, 4998–5003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.C.; Ren, J.W.; Xian, H.H.; Yin, C.; Yuan, Y.; Li, Y.; Ji, R.Y.; Chu, C.; Qiao, Z.; Jiao, X.A. rOmpF and OMVs as efficient subunit vaccines against Salmonella enterica serovar Enteritidis infections in poultry farms. Vaccine 2020, 38, 7094–7099. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Wang, J.; Jiao, Y.; Tang, P.; Song, L.; Xiong, D.; Yin, Y.; Pan, Z.; Jiao, X. Expression of recombinant Newcastle disease virus F protein in Pichia pastoris and its immunogenicity using flagellin as the adjuvant. Protein Expr. Purif. 2016, 128, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Sudworth, A.; Segers, F.M.; Yilmaz, B.; Guslund, N.C.; Macpherson, A.J.; Dissen, E.; Qiao, S.W.; Inngjerdingen, M. Innate lymphoid cell characterization in the rat and their correlation to gut commensal microbes. Eur. J. Immunol. 2022. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequences (5′→3′) | Restricted Site | Usage |
|---|---|---|---|
| his-SseB-F | AAGGTAGGCATATGGAGCTCGGTACCATGTCTTCAGGAAACATCTT | Kpn I | Construction of pCold-SseB |
| his-SseB-R | GACTGCAGGTCGACAAGCTTGAATTCTCATGAGTACGTTTTCTG | EcoR I | |
| GST- SseB-F | TTCTGTTCCAGGGGCCCCTGGGATCCATGTCTTCAGGAAACATCTT | BamH I | Construction of pGEX-6p-1-SseB |
| GST- SseB-R | GGCCGCTCGAGTCGACCCGGGAATTCTCATGAGTACGTTTTCTGC | EcoR I |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, X.; Huang, T.; Shen, H.; Meng, C.; Jiao, X.; Pan, Z. Salmonella Enteritidis Subunit Vaccine Candidate Based on SseB Protein Co-Delivered with Simvastatin as Adjuvant. Pathogens 2022, 11, 443. https://doi.org/10.3390/pathogens11040443
Kang X, Huang T, Shen H, Meng C, Jiao X, Pan Z. Salmonella Enteritidis Subunit Vaccine Candidate Based on SseB Protein Co-Delivered with Simvastatin as Adjuvant. Pathogens. 2022; 11(4):443. https://doi.org/10.3390/pathogens11040443
Chicago/Turabian StyleKang, Xilong, Tingting Huang, Huanhuan Shen, Chuang Meng, Xinan Jiao, and Zhiming Pan. 2022. "Salmonella Enteritidis Subunit Vaccine Candidate Based on SseB Protein Co-Delivered with Simvastatin as Adjuvant" Pathogens 11, no. 4: 443. https://doi.org/10.3390/pathogens11040443
APA StyleKang, X., Huang, T., Shen, H., Meng, C., Jiao, X., & Pan, Z. (2022). Salmonella Enteritidis Subunit Vaccine Candidate Based on SseB Protein Co-Delivered with Simvastatin as Adjuvant. Pathogens, 11(4), 443. https://doi.org/10.3390/pathogens11040443