
The Biological Traumatization of Crops Due to the Enzyme Stage of Enzyme-Mycotic Seed Depletion

 ,
, 
 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Sites
2.2. Plant Materials
2.3. Climatic Conditions and Soil Characteristics
2.4. Methodology
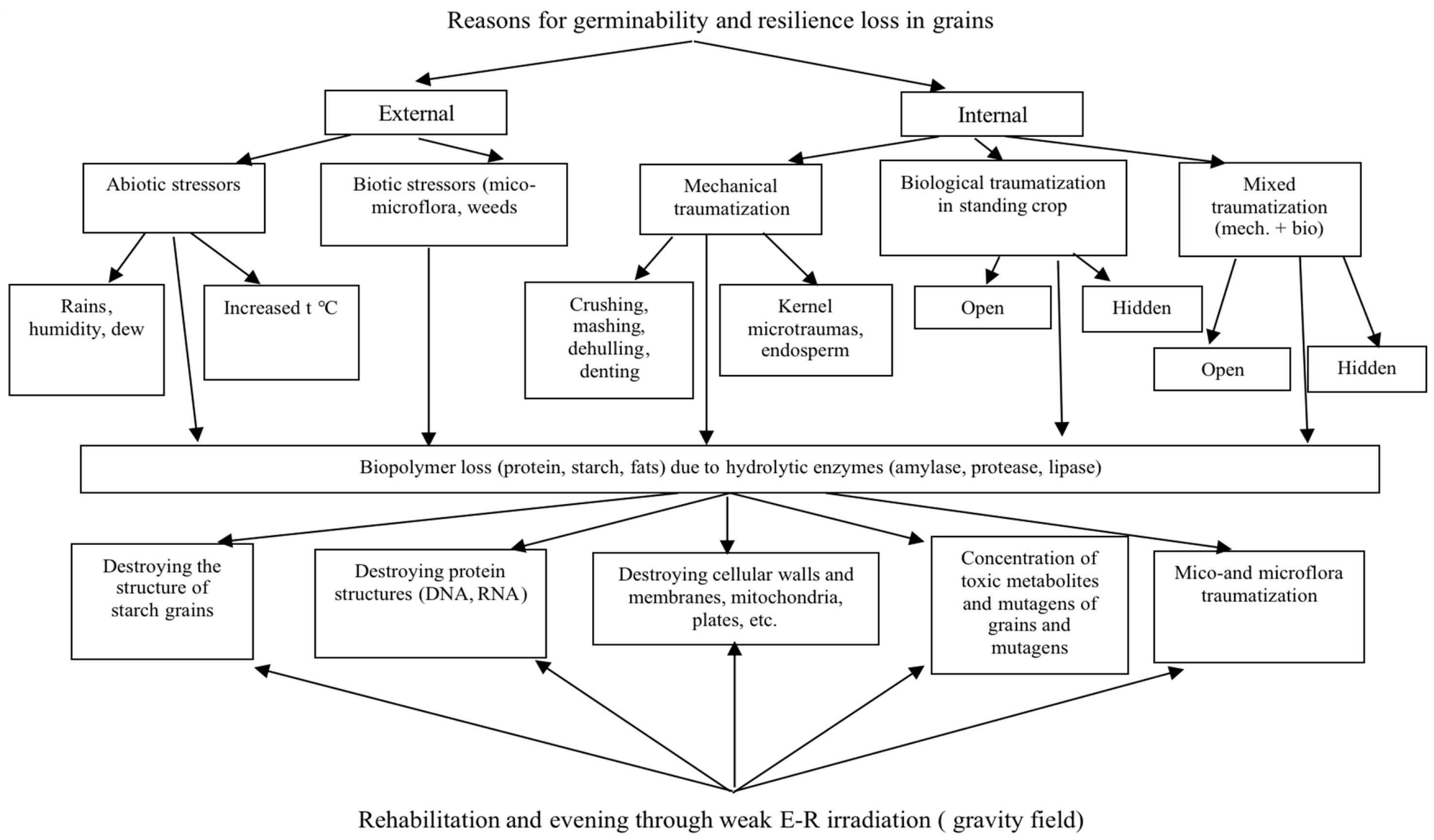
3. Results and Discussion
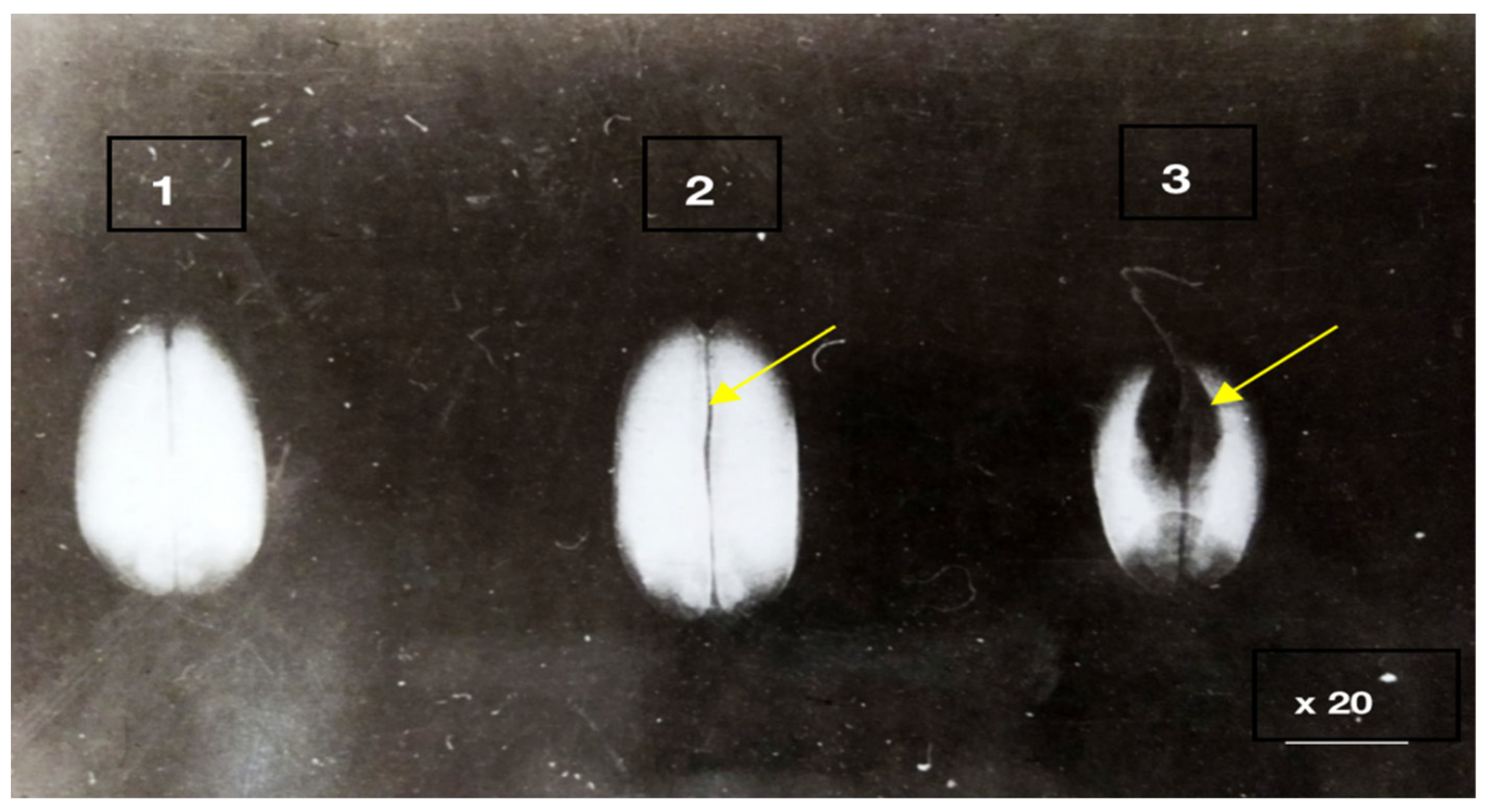
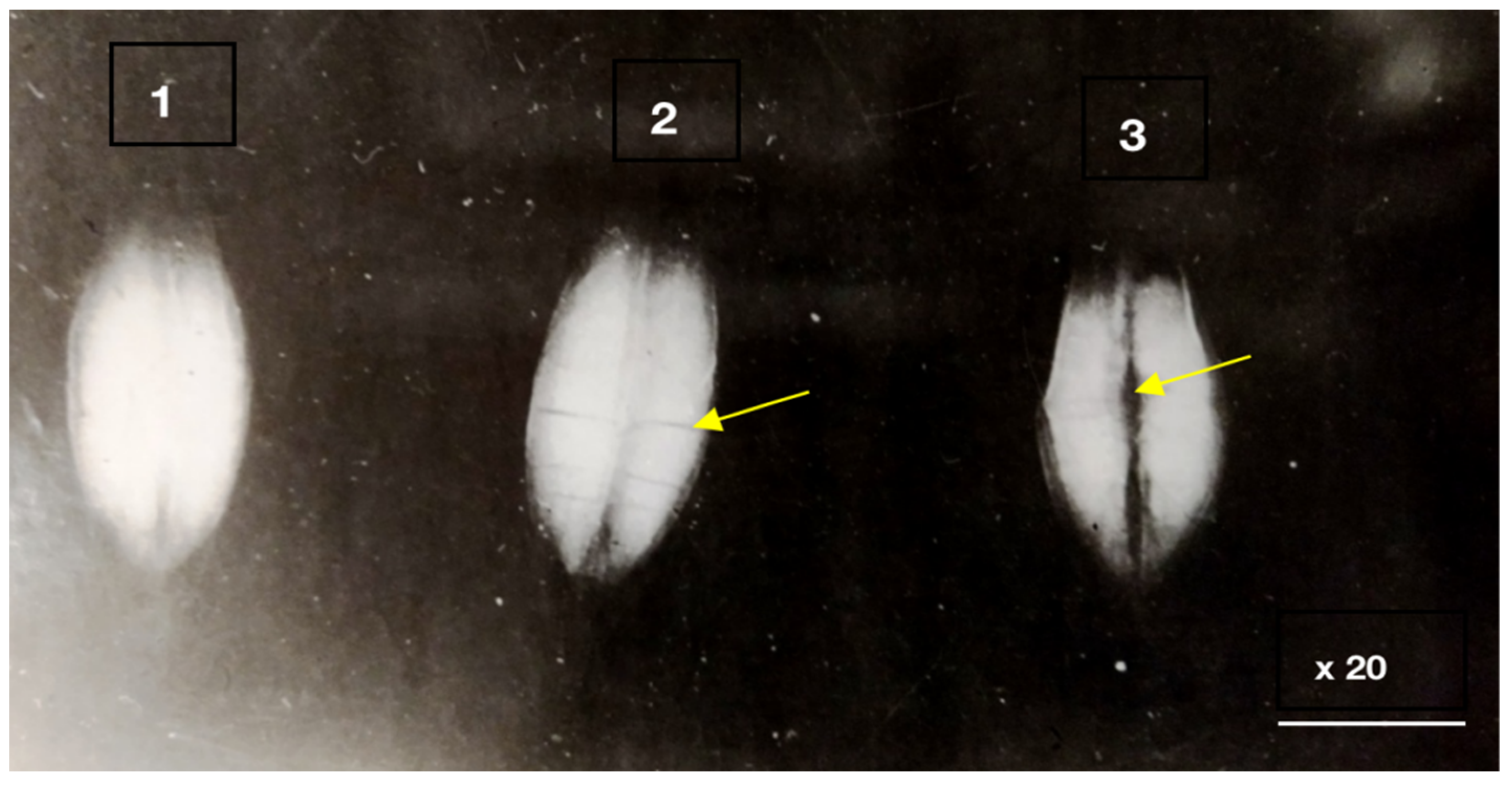
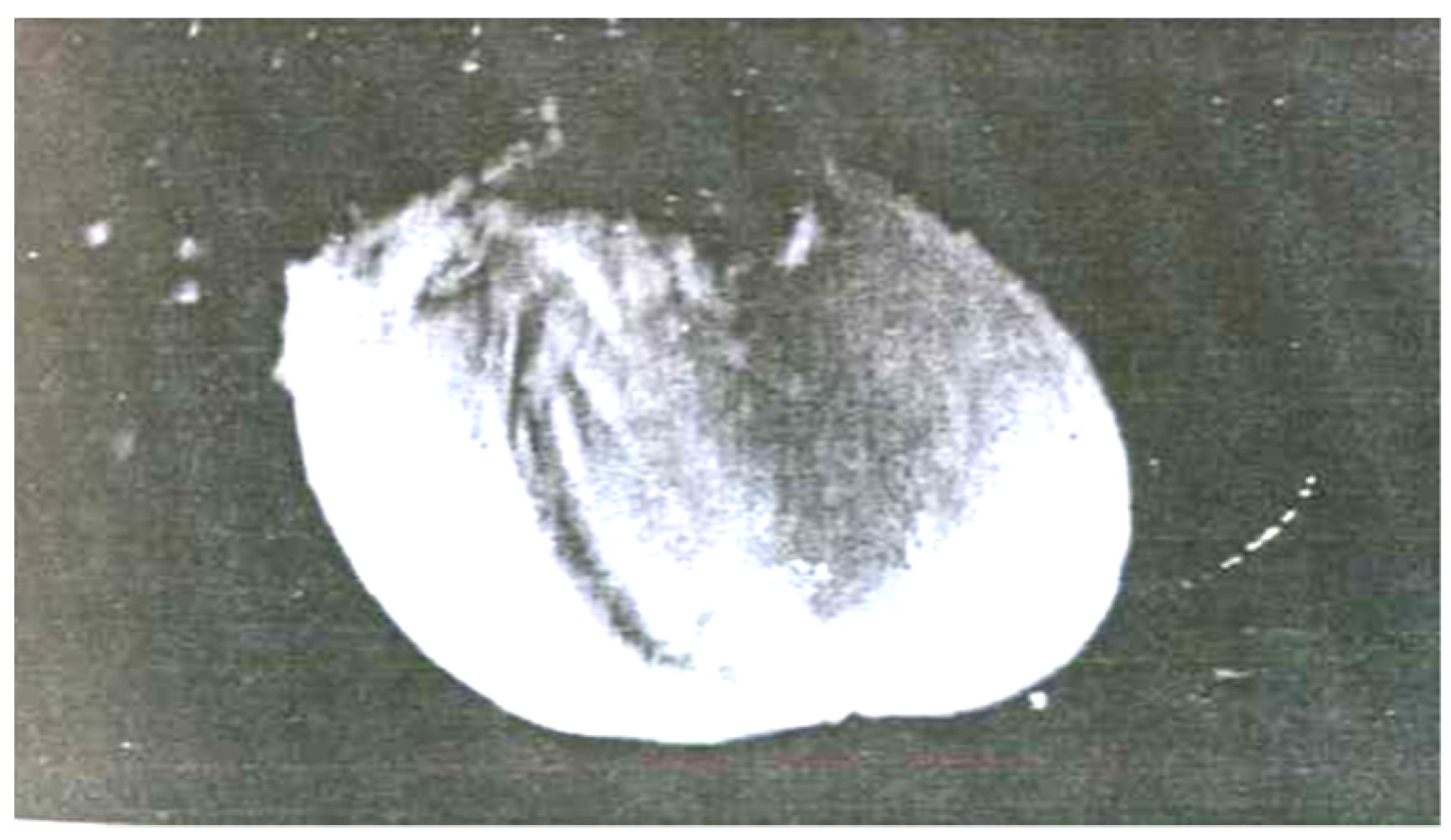
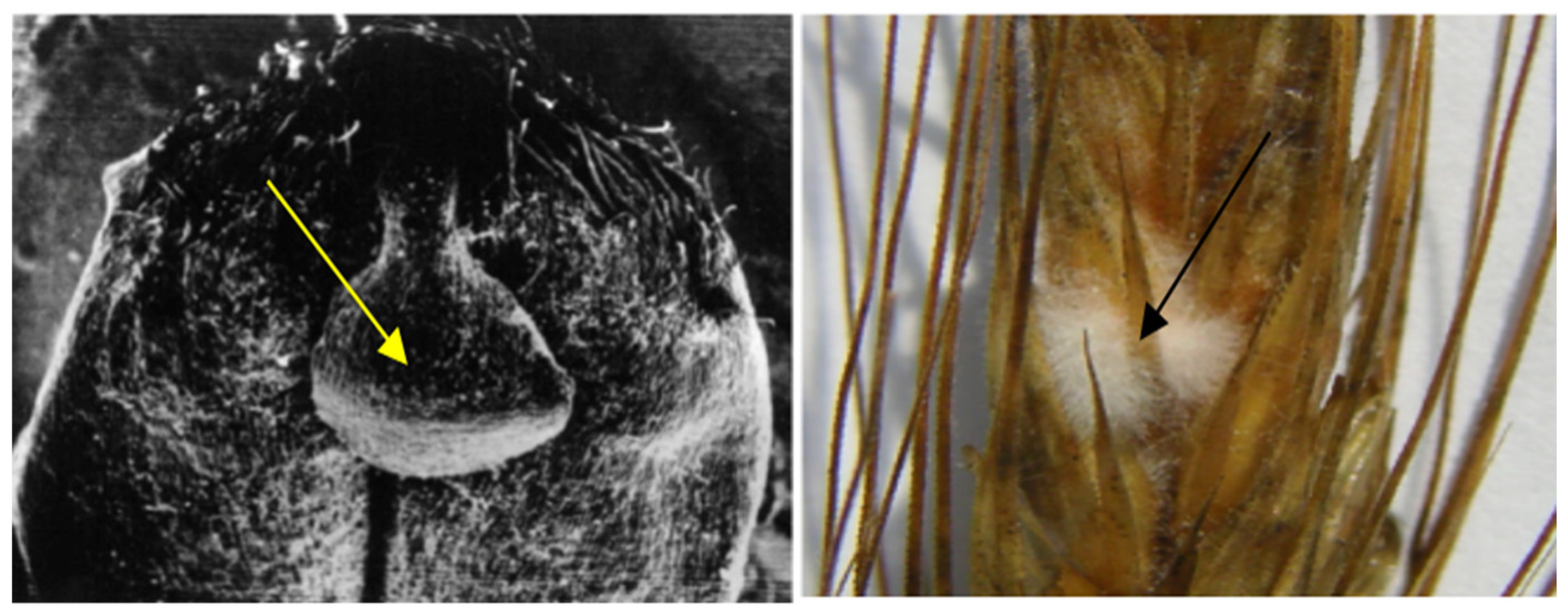
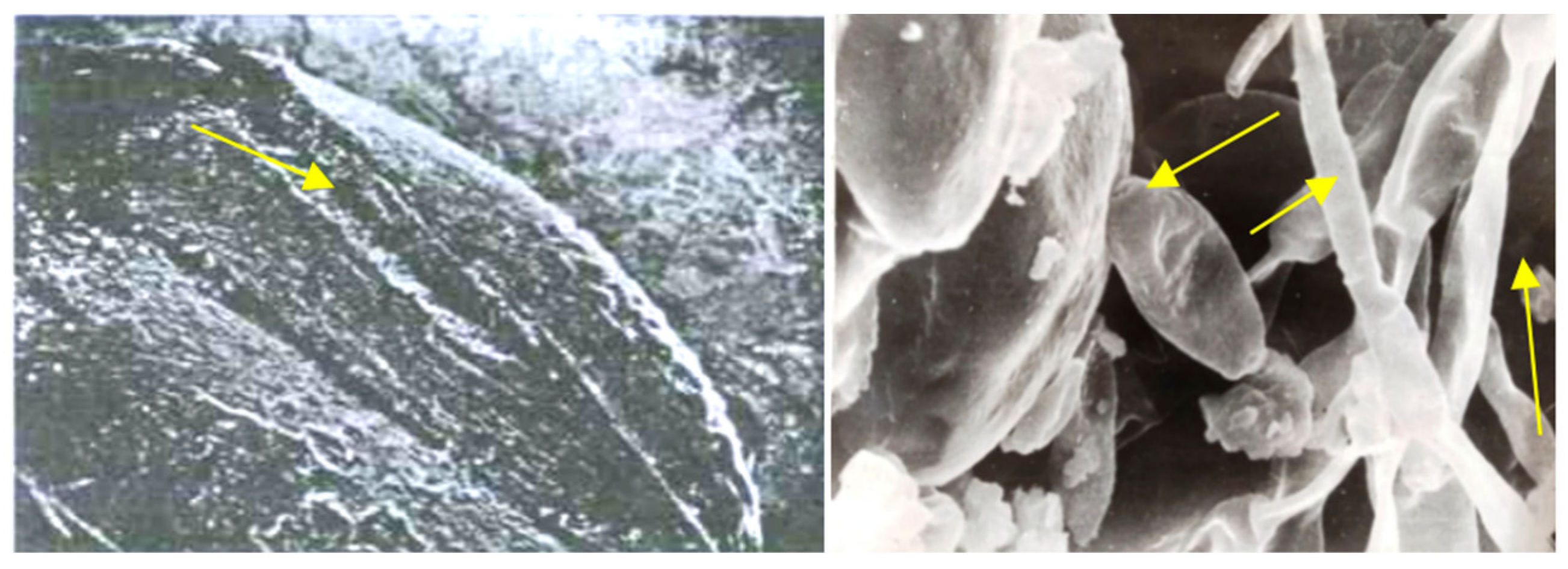
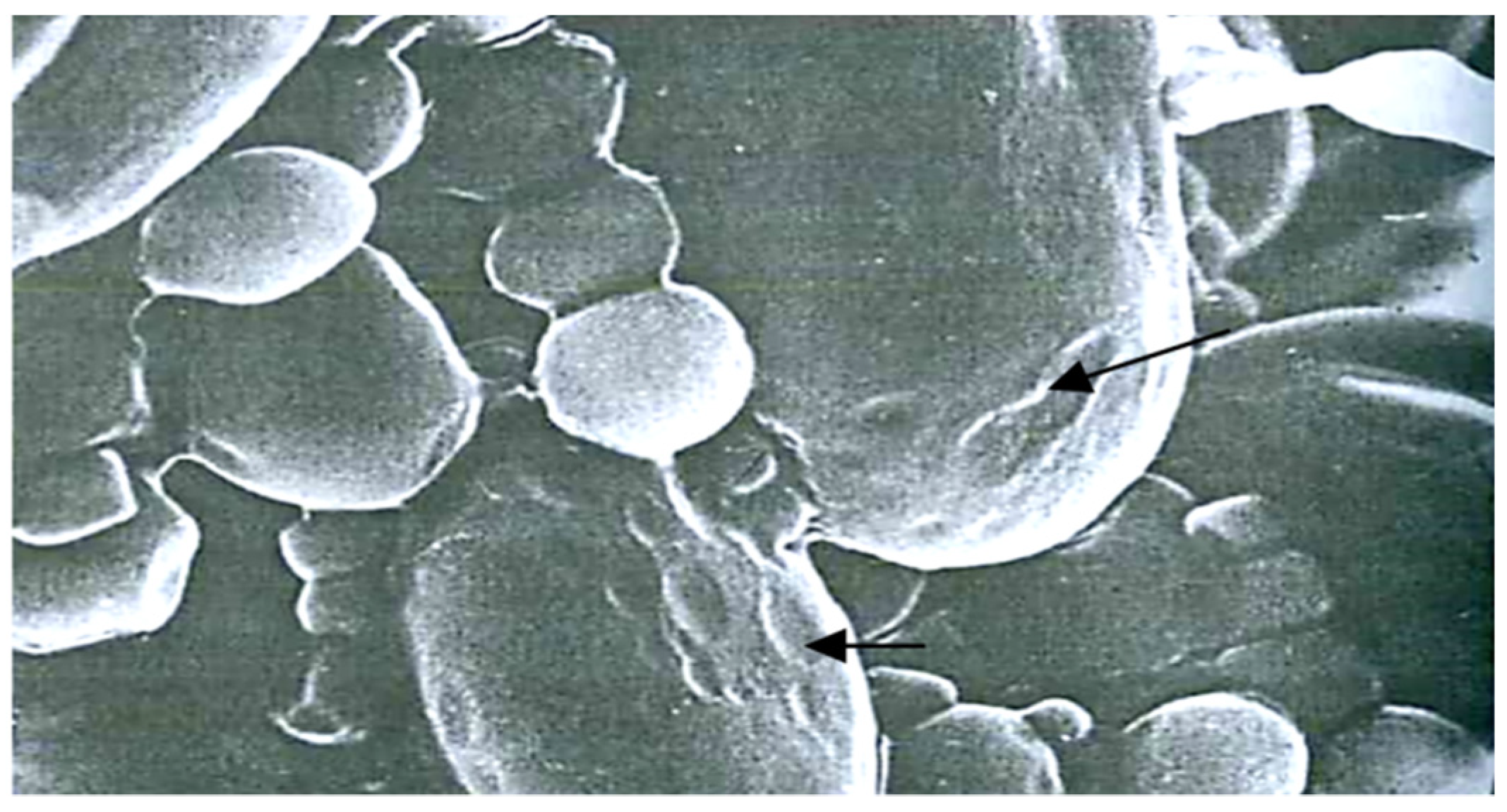
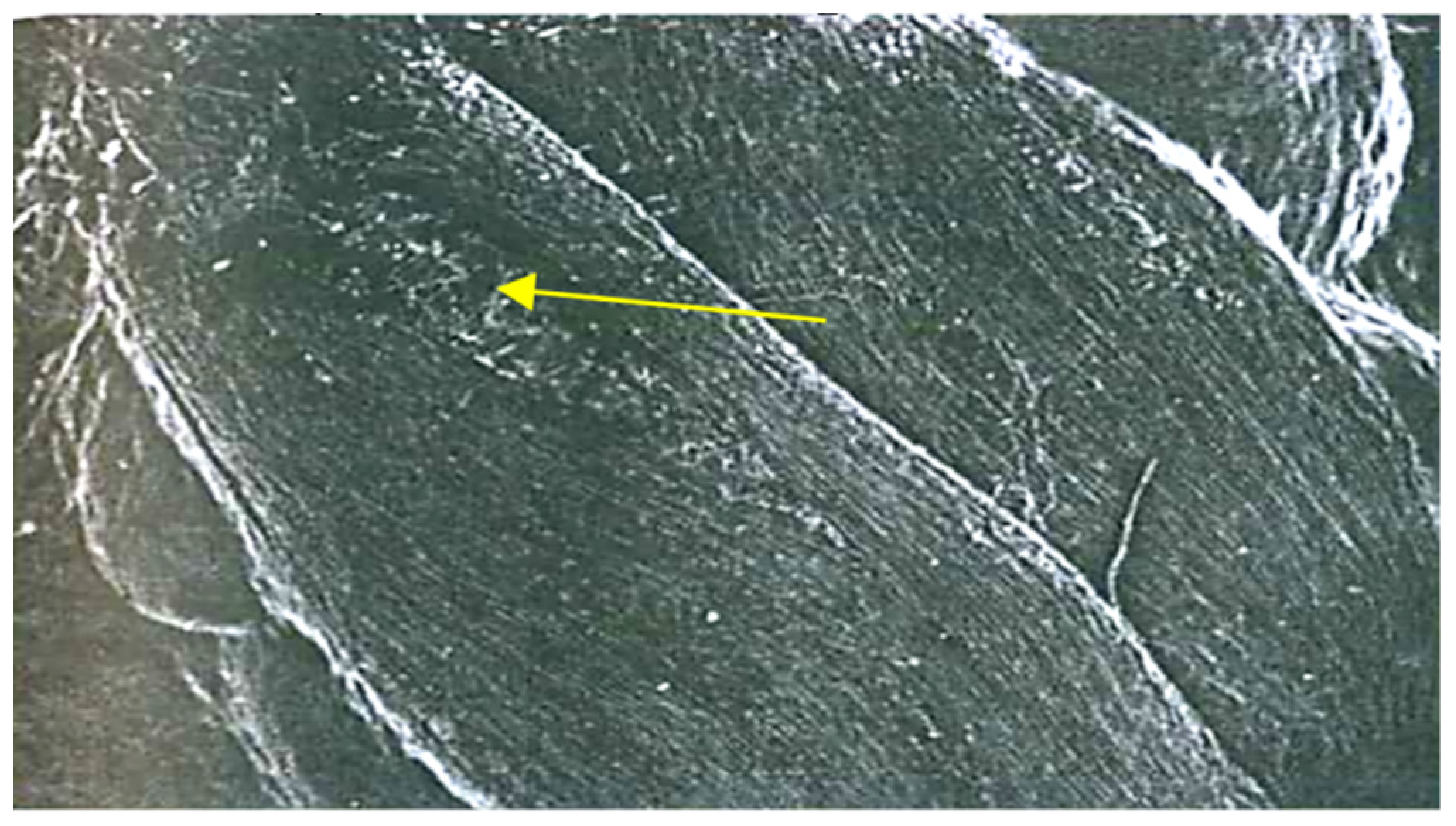
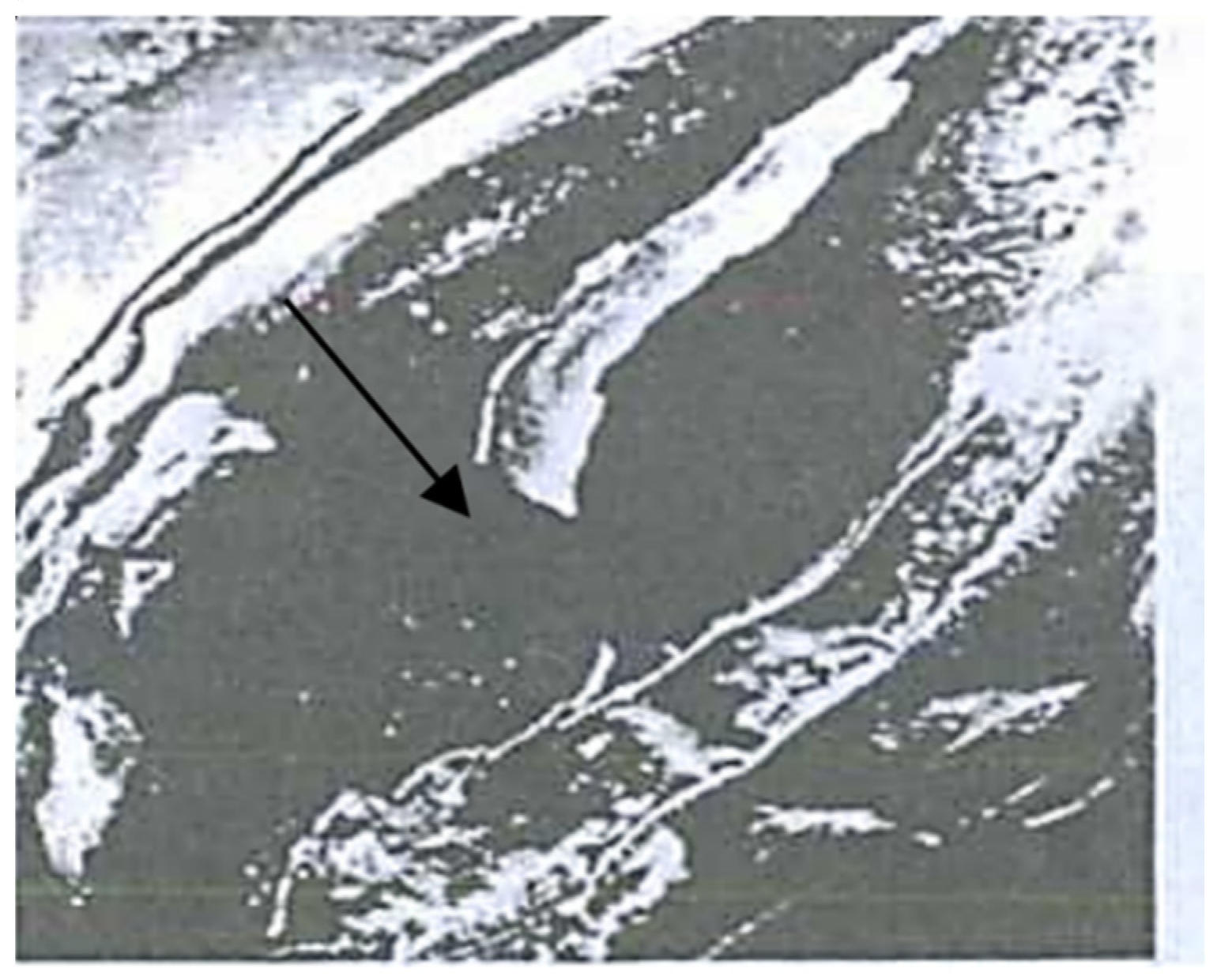
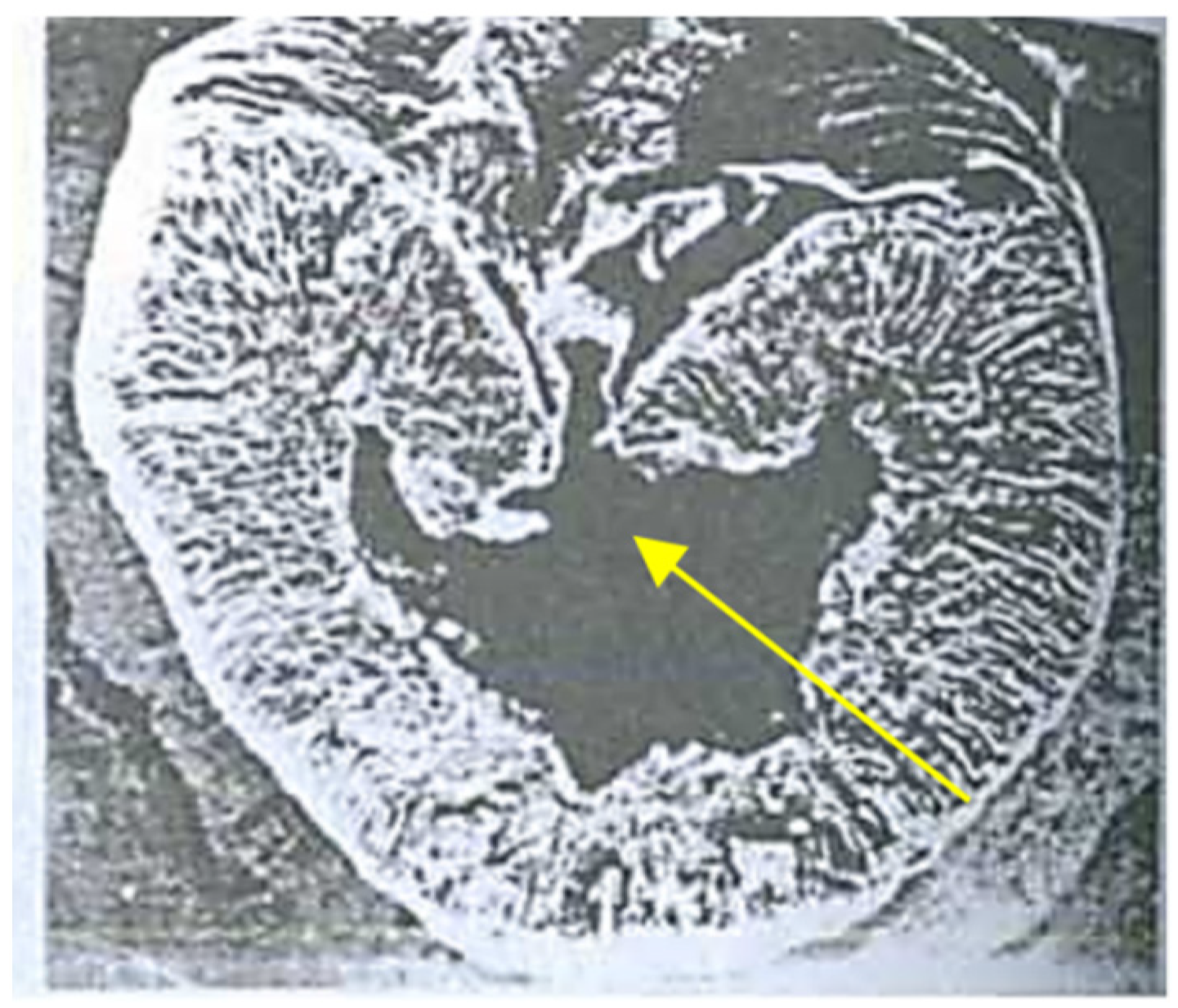
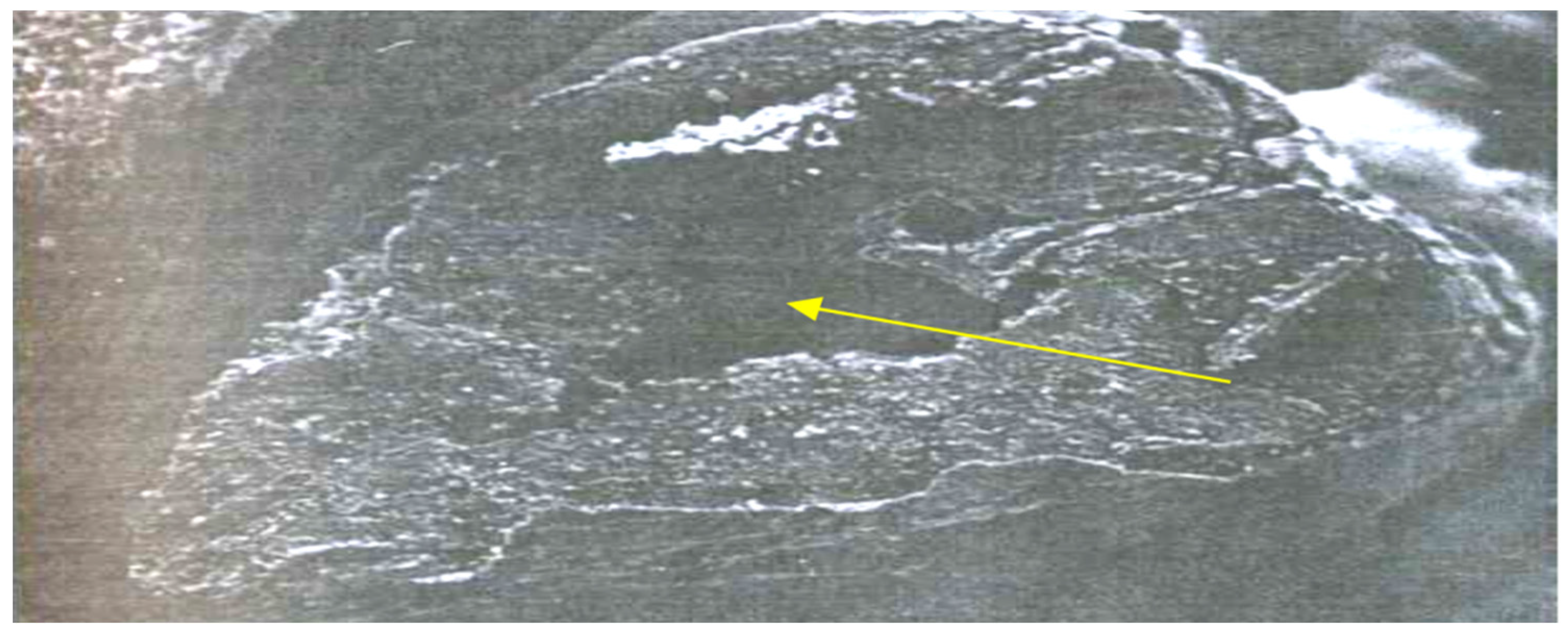
3.1. Hidden Biological Traumatization during Blooming
3.2. Open and Hidden Biological Traumatization during Grain Formation and the Milk/Wax Ripeness Phases
3.3. Open and Hidden Biological Traumatization during the Full Ripeness Phase
4. Conclusion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Polityko, P.; Rebouh, N.Y.; Kucher, D.; Vvedenskiy, V.; Kapranov, V.; Atmachian, G.; Behzad, A.; Urazova, E.; Khomenets, N.; Pakina, E.; et al. Productivity and grain quality of three spring wheat (Triticum aestivum L.) cultivars under three cultivation technologies. Eurasian J. Biosci. 2020, 14, 2089–2095. [Google Scholar]
- Latati, M.; Rebouh, N.Y.; Aouiche, A.; Laouar, M. Modeling the functional role of the microorganisms in the daily exchanges of carbon and nitrogen in intercropping system under mediterranean conditions. Agron. Res. 2019, 17, 559–573. [Google Scholar] [CrossRef]
- Rebouh, N.Y.; Latati, M.; Polityko, P.; Maisam, Z.; Kavhiza, N.J.; Garmasch, N.; Pakina, E.; Lyshko, M.; Engeribo, A.; Orujov, E.; et al. Improvement of weeds management system and fertilisers application in winter wheat (Triticum aestivum L.) cultivation technologies. Agriculture 2021, 67, 76–86. [Google Scholar] [CrossRef]
- FAO. FAO Cereal Supply and Demand Brief; Food and Agriculture Organization of the United Nations: Washington, DC, USA, 2021. [Google Scholar]
- Lcamo, J.; Dronin, N.; Endejan, M.; Golubev, G.; Kirilenko, A. A new assessment of climate change impacts on food productionshortfalls and water availability in Russia. Glob. Environ. Chang. 2007, 17, 429–444. [Google Scholar]
- Temirbekova, S.K.; Kulikov, I.M.; Afanasyeva, Y.V.; Beloshapkina, O.O.; Kalashnikova, E.A.; Kirakosyan, R.N.; Dokukin, P.A.; Kucher, D.E.; Latati, M.; Rebouh, N.Y. The Evaluation ofWinter Wheat Adaptation to Climate Change in the Central Non-Black Region of Russia: Study of the Gene Pool Resistance of Wheat from the N.I. Vavilov Institute of Plant Industry (VIR) World Collection to Abiotic Stress Factors. Plants 2021, 10, 2337. [Google Scholar] [CrossRef] [PubMed]
- Zargar, M.; Rebouh, N.; Pakina, E.; Gadzhikurbanov, A.; Lyashko, M.; Ortskhanov, B. Impact of climate change on cerealproduction in the highlands of eastern Algeria. Res. Crop. 2017, 18, 575–582. [Google Scholar] [CrossRef]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef]
- Poudel, P.B.; Poudel, M.R. Heat stress effects and tolerance in wheat: A review. J. Biol. Today’s World 2020, 9, 1–6. [Google Scholar]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savary, S.; Ficke, A.; Aubertot, J.N. Clayton Hollier. Crop losses due to diseases and their implications for global food production losses and food security. Food Sec. 2012, 4, 519–537. [Google Scholar] [CrossRef]
- Rebouh, N.Y.; Polityko, P.; Latati, M.; Pakina, E.; Kapranov, V.; Imbia, A.; Norezzine, A.; Gadzhikurbanov, A.; Vvedenskiy, V.; Iguerouada, M. Influence of three pest management treatments against aphid, sitobion avenae in winter wheat (Triticum aestivum l.) under moscow area conditions. Res. Crop. 2019, 20, 381–388. [Google Scholar]
- Goswami, R.S.; Kistler, T.I.C. Headingfor disaster: Fusarium graminearum oncereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Rebouh, N.Y.; Polityko, P.M.; Pakina, E.; Plushikov, V.G.; Norezzine, A.; Gadzhikurbanov, A.; Iguer-Ouada, M. Impact of three integrated crop protection treatments on the varieties of winter wheat (Triticum aestivum L.) in Moscow area, Russia. Res. Crop. 2019, 20, 161–168. [Google Scholar] [CrossRef]
- Temirbekova, S.; Jafarov, I.; Kulikov, I.; Afanaseva, Y.; Kalashnikova, E. Gene Pool of Winter Wheat from theWorld Collection of N.I. Vavilov Institute of Plant Industry (VIR) for Biotic Stress Resistance. Pathogens 2021, 10, 514. [Google Scholar] [CrossRef] [PubMed]
- Temirbekova, S.K.; Kulikov, I.M.; Afanasieva, Y.V.; Zuev, E.V.; Beloshapkina, O.O.; Dementiev, A.V. Source material of a winter wheat for a stress-resistant varieties selection. Vestnik Russ. Agric. Sci. 2021, 16–24. (In Russian) [Google Scholar] [CrossRef]
- Rebouh, N.Y.; Latati, M.; Polityko, P.; Kucher, D.; Hezla, L.; Norezzine, A.; Iguer-Ouada, M. Influence of three cultivation technologies to control Fusarium spp. in winter wheat (Triticum aestivum L.) production under Moscow conditions. Res. Crop. 2020, 21, 17–25. [Google Scholar] [CrossRef]
- Temirbekova, S.K.; Kulikov, I.M.; Afanasieva, Y.V.; Okhotnikova, T.V. Initional material for breeding stress-resistant winter wheat cultivars. Vestnik Russ. Agric. Sci. 2021, 10, 8–14. (In Russian) [Google Scholar] [CrossRef]
- Li, H.J.; Murray, T.D.; McIntosh, R.A.; Zhou, Y. Wheat breeding in northern China: Achievements and technical advances. Crop. J. 2019, 7, 718–729. [Google Scholar] [CrossRef]
- Xiang, C.; Huang, J. The role of exotic wheat germplasms in wheat breeding and their impact on wheat yield and production inChina. China Econ. Rev. 2020, 62, 101239. [Google Scholar] [CrossRef]
- Li, H.; Timothy, D.; Murray, T.D.; McIntosh, R.A.; Zhou, Y. Breeding new cultivars for sustainable wheat production. Crop. J. 2019, 7, 715–717. [Google Scholar] [CrossRef]
- Halford, N.G. New insights on the effects of heat stress on crops. J. Exp. Bot. 2009, 60, 4215–4216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Ashkar, I.; Ibrahim, A.; Ghazy, A.; Attia, K.; Al-Ghamdi, A.A.; Al-Dosary, M.A. Assessing the correlations and selection criteriabetween different traits in wheat salt-tolerant genotypes. Saudi J. Biol. Sci. 2021, 28, 5414–5427. [Google Scholar] [CrossRef]
- Gradchaninova, O.D.; Filatenko, A.A.; Rudenko, M.I. Methodical Guidelines for Studying the World Collection of Wheat; VIR Publ.: Leningrad, Russia, 1987; p. 28. [Google Scholar]
- Merezhko, A.F.; Udachin, R.A.; Zuev, E.V.; Filotenko, A.A.; Serbin, A.A.; Liapunova, O.A.; Kosov, V.I.; Kurkiev, U.K.; Okhotnikova, T.V.; Nazrubekov, N.A.; et al. Replenishment, Preservation in a Viable State, and Studying of a World Collection of Wheat, Goatgrass, and Triticale. Methodical Guidelines; VIR Publ.: St. Petersburg, Russia, 1999; p. 81. [Google Scholar]
- Triticum, L. The International COMECON List of Descriptors for the Genus; Institute of Plant Industry: Leningrad, Russia, 1989; p. 43. [Google Scholar]
- Savin, V.N.; Arkhipov, M.V.; Badenko, A.L.; Ioffe, Y.u.K.; Grun, L.B. X-ray to detect internal damage and their impact on the yield quality of seeds. S.-X. Sci. 1981, 10, 99–104. [Google Scholar]
- Musaev, F.B.; Beletskiy, S.L. History and Prospects for the Application of X-ray Diffraction Analysis in Seed Breeding and Seed Study. J. Russ. Univ. Radioelectron. 2021, 24, 6–15. [Google Scholar] [CrossRef]
- Gromova, B.B.-O.; Temirbekova, S.K.; Buys, J. Evaluation of the resistance of cereals against infection by Fusarium of the ears during the enzymatic stadium of enzyme-mycotic seed exhaustion (EMES). In Proceedings of the BRIGTON Conference, Pests-Diseases, British Crop Protection COUNCIL, Farnham, British, 18–21 November 1996. [Google Scholar]
- GOST 12038-84; Seeds of Agricultural Crops. Methods of Analysis. Publishing House of Standards, SB. GOSTs: Moscow, Russia, 2004; p. 47c.
- Vavilov, N.I. Selected Works in Five Volumes. In The Law of Homologous Series in Hereditary Variability; Nauka: Leningrad, Russia, 1965; Volume 5, pp. 179–220. [Google Scholar]
- Temirbekova, S.K. On the Problem of Enzyme-Mycotic Depletion of Seeds (Grain Exhaustion) in the Plant Industry, 2nd ed.; Russian Academy of Agricultural Sciences: Moscow, Russia, 2000; p. 306. [Google Scholar]
- Temirbekova, S.K.; Cheremisova, T.D.; Kulikov, I.M.; Afanasieva, Y.V.; Zuev, E.V.; Potapova, E.S. VIRWorld Collection as Key to N.I. Vavilov’s Ideal Variety Resistant to the Abiotic and Biotic Stresses of Russia’s Central Region; FGBNU FNC Sadovodstva: Moscow, Russia, 2020; p. 108. [Google Scholar]
- Roberts, E.H. Viability of Seeds; Charman and Hall Ltd.: London, UK, 1972. [Google Scholar]
- Vavilov, N.I. Selected Works in Five Volumes. In Immunity Problems of Cultivated Plants; Nauka: Leningrad, Russia, 1964; Volume 4, p. 518. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| № Registration VIR | Cultivars | Origins | Winter Hardiness, Point | Weight per 1000 grains, g | Yield, g/m2 |
|---|---|---|---|---|---|
| Dry matter losses up to 15%, stability of group I: stable | |||||
| 45335 | Ibis | Germany | 5 | 34.9 | 260 |
| 49916 | Zarya | Russia | 7 | 39.6 | 350 |
| - | Gelderseris | Netherlands | 8 | 40.4 | 425 |
| 64027 | Bassard | Germany | 8 | 41.3 | 400 |
| 64160 | Moskovskaya 39 | Russia | 7 | 45.7 | 430 |
| 58526 | Ivanovskaya 16 | Ivanovo region | 9 | 41.0 | 367 |
| Losses of dry matter up to 25%, stability of group II: relatively stable | |||||
| 65760 | Moskovskaya 56 | Russia | 7 | 48.0 | 400 |
| 65757 | Nemchnovskaya 24 | Russia | 5 | 41.7 | 450 |
| 64013 | Bersy | Netherlands | 7 | 39.0 | 290 |
| 64030 | Zentos | Switzerland | 5 | 37.5 | 280 |
| 62052 | Obelisk | Germany | 5 | 38.2 | 300 |
| 64034 | Orestis | Germany | 5 | 37.4 | 310 |
| 34230 | Varmalands | Sweden | 7 | 39.3 | 302 |
| Losses of dry matter up to 35%, stability of group III: highly tolerant | |||||
| 43920 | Mironovskaya 808 | Ukraine | 9 | 40.7 | 380 |
| 54610 | Yantarnaya 50 | Russia | 7 | 38.9 | 310 |
| 63560 | Kazanskaya 285 | Russia | 7 | 39.8 | 320 |
| 63041 | Umanka | Russia | 7 | 38.7 | 300 |
| 63404 | Nika Kuban | Russia | 7 | 39.1 | 340 |
| Dry matter losses of more than 35%, stability of group IV: unstable (susceptible) | |||||
| 54508 | Polokarlik 3 | Ukraine | 5 | 34.1 | 220 |
| 57582 | Fakta | Germany | 5 | 33.4 | 210 |
| 64028 | Faktor | Germany | 5 | 35.6 | 180 |
| 63016 | lives | Finland | 5 | 36.1 | 200 |
| 63273 | Expert | Austria | 4 | 35.4 | 185 |
| 64051 | Nike | Poland | 5 | 36.5 | 170 |
| 63119 | Zadorinka | Irkutsk region | 5 | 38.1 | 240 |
| LSD05 | 0.8 | 90 | |||
| Cultivars | Germination, % | Survivability %, 2018 г. | Infestation Rate, % | |||||
|---|---|---|---|---|---|---|---|---|
| 2013 г. | 2018 г. | |||||||
| 2013 | 2018 | Difference | Altemaria altematа | Fusarium spp. | Altemaria altemata | Fusarium spp. | ||
| Resistance to EMSD | ||||||||
| Ibis | 92.0 | 64.0 | 28.0 | 67.0 | 7.0 | 3.0 | 25.0 | 10.0 |
| Zarya | 94.0 | 60.0 | 34.0 | 71.0 | 12.0 | 4.0 | 28.0 | 12.0 |
| Gelderseris | 90.0 | 61.0 | 29.0 | 69.0 | 10.0 | 5.0 | 30.0 | 10.0 |
| Bussard | 95.0 | 58.0 | 37.0 | 70.0 | 8.0 | 3.0 | 20.0 | 7.0 |
| Ivanovskaya 16 | 90.0 | 62.0 | 28.0 | 73.0 | 15.0 | 8.0 | 31.0 | 10.0 |
| Moskovskaya 39 | 92.0 | 67.0 | 25.0 | 72.0 | 12.0 | 5.0 | 15.0 | 10.0 |
| Relatively resistant to EMSD | ||||||||
| Moskovskaya 56 cт. | 95.0 | 70.0 | 25.0 | 75.0 | 7.0 | 2.0 | 21.0 | 12.0 |
| Bersy | 90.0 | 57.0 | 33.0 | 64.0 | 12.0 | 5.0 | 28.0 | 10.0 |
| Zentos | 90.0 | 55.0 | 35.0 | 60.0 | 8.0 | 4.0 | 51.0 | 10.0 |
| Obelisk | 89.0 | 59.0 | 30.0 | 67.0 | 10.0 | 7.0 | 49.0 | 15.0 |
| Orestis | 85.0 | 50.0 | 35.0 | 62.0 | 15.0 | 10.0 | 52.0 | 15.0 |
| Varmalands | 87.0 | 48.0 | 39.0 | 55.0 | 10.0 | 5.0 | 46.0 | 15.0 |
| Highly tolerant to EMSD | ||||||||
| Mironovskaya 808 sт. | 85.0 | 52.0 | 33.0 | 61.0 | 12.0 | 8.0 | 50.0 | 22.0 |
| Nika Kuban | 90.0 | 43.0 | 47.0 | 53.0 | 10.0 | 12.0 | 45.0 | 18.0 |
| Kazanskaya 285 | 80.0 | 45.0 | 35.0 | 55.0 | 15.0 | 10.0 | 57.0 | 25.0 |
| Umanka | 85.0 | 47.0 | 38.0 | 54.0 | 9.0 | 15.0 | 40.0 | 30.0 |
| Not resistant to EMSD | ||||||||
| Polokarilik 3 | 87.0 | 35.0 | 52.0 | 37.0 | 12.0 | 25.0 | 52.0 | 100 |
| Fakta | 85.0 | 34.0 | 51.0 | 33.0 | 20.0 | 28.0 | 56.0 | 100 |
| Faktor | 87.0 | 30.0 | 57.0 | 34.0 | 20.0 | 18.0 | 60.0 | 100 |
| lives | 80.0 | 36.0 | 44.0 | 39.0 | 10.0 | 27.0 | 85.0 | 90.0 |
| Expert | 85.0 | 33.0 | 52.0 | 41.0 | 30.0 | 25.0 | 80.0 | 100 |
| Nike | 80.0 | 31.0 | 49.0 | 35.0 | 25.0 | 18.0 | 90.0 | 100 |
| Zadorinka | 65.0 | 17.0 | 48.0 | 15.0 | 20.0 | 35.0 | 60.0 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Temirbekova, S.K.; Kulikov, I.M.; Afanasyeva, Y.V.; Ashirbekov, M.Z.; Beloshapkina, O.O.; Kalashnikova, E.A.; Sardarova, I.; Begeulov, M.S.; Kucher, D.E.; Ionova, N.E.; et al. The Biological Traumatization of Crops Due to the Enzyme Stage of Enzyme-Mycotic Seed Depletion. Pathogens 2022, 11, 376. https://doi.org/10.3390/pathogens11030376
Temirbekova SK, Kulikov IM, Afanasyeva YV, Ashirbekov MZ, Beloshapkina OO, Kalashnikova EA, Sardarova I, Begeulov MS, Kucher DE, Ionova NE, et al. The Biological Traumatization of Crops Due to the Enzyme Stage of Enzyme-Mycotic Seed Depletion. Pathogens. 2022; 11(3):376. https://doi.org/10.3390/pathogens11030376
Chicago/Turabian StyleTemirbekova, Sulukhan K., Ivan M. Kulikov, Yuliya V. Afanasyeva, Mukhtar Z. Ashirbekov, Olga O. Beloshapkina, Elena A. Kalashnikova, Irina Sardarova, Marat Sh. Begeulov, Dmitry E. Kucher, Natalia E. Ionova, and et al. 2022. "The Biological Traumatization of Crops Due to the Enzyme Stage of Enzyme-Mycotic Seed Depletion" Pathogens 11, no. 3: 376. https://doi.org/10.3390/pathogens11030376
APA StyleTemirbekova, S. K., Kulikov, I. M., Afanasyeva, Y. V., Ashirbekov, M. Z., Beloshapkina, O. O., Kalashnikova, E. A., Sardarova, I., Begeulov, M. S., Kucher, D. E., Ionova, N. E., & Rebouh, N. Y. (2022). The Biological Traumatization of Crops Due to the Enzyme Stage of Enzyme-Mycotic Seed Depletion. Pathogens, 11(3), 376. https://doi.org/10.3390/pathogens11030376