
Clinical Progression of Theileria haneyi in Splenectomized Horses Reveals Decreased Virulence Compared to Theileria equi


Abstract
1. Introduction
2. Results
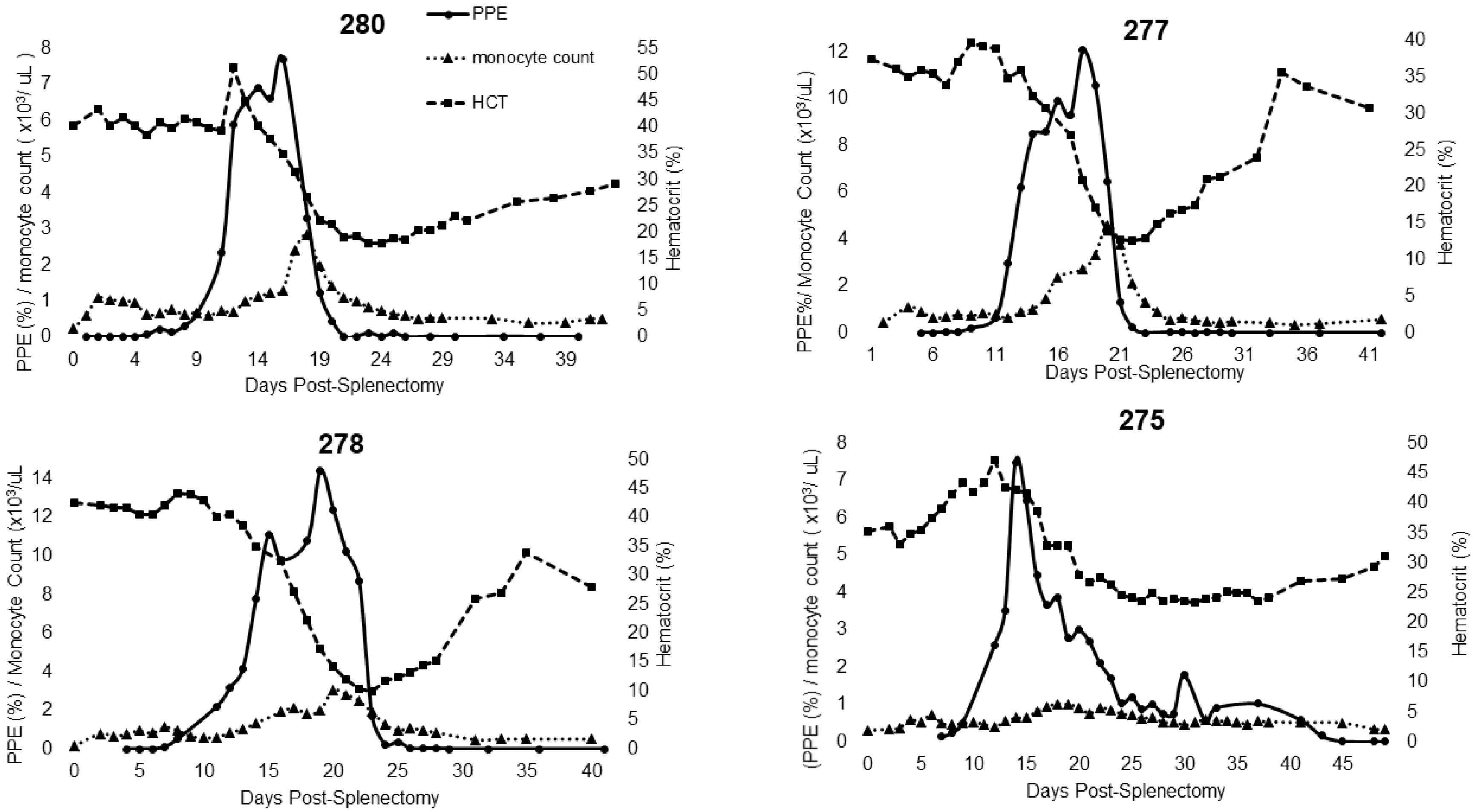
2.1. Characterization of Experimental T. haneyi Infection of Splenectomized Horses
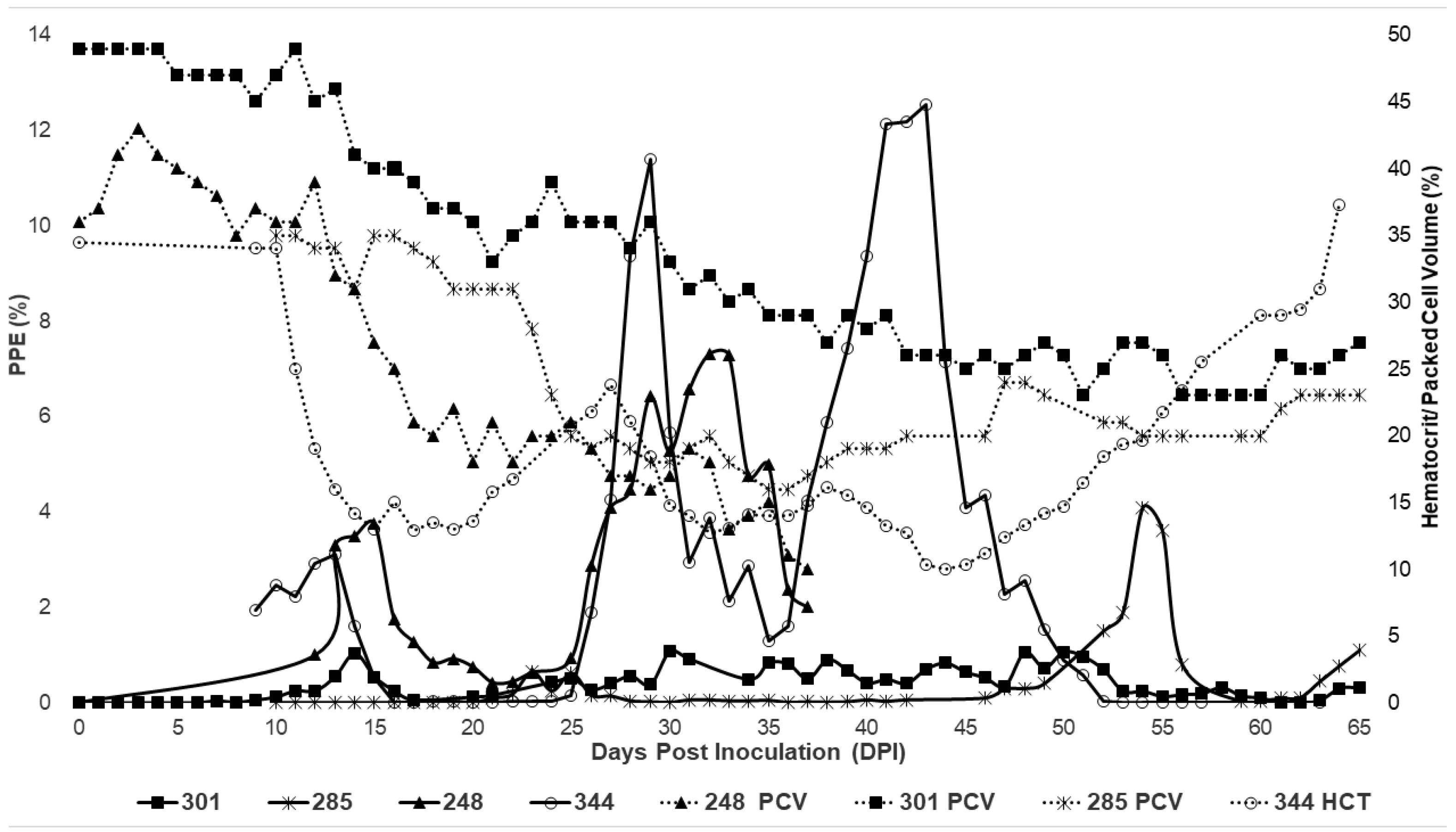
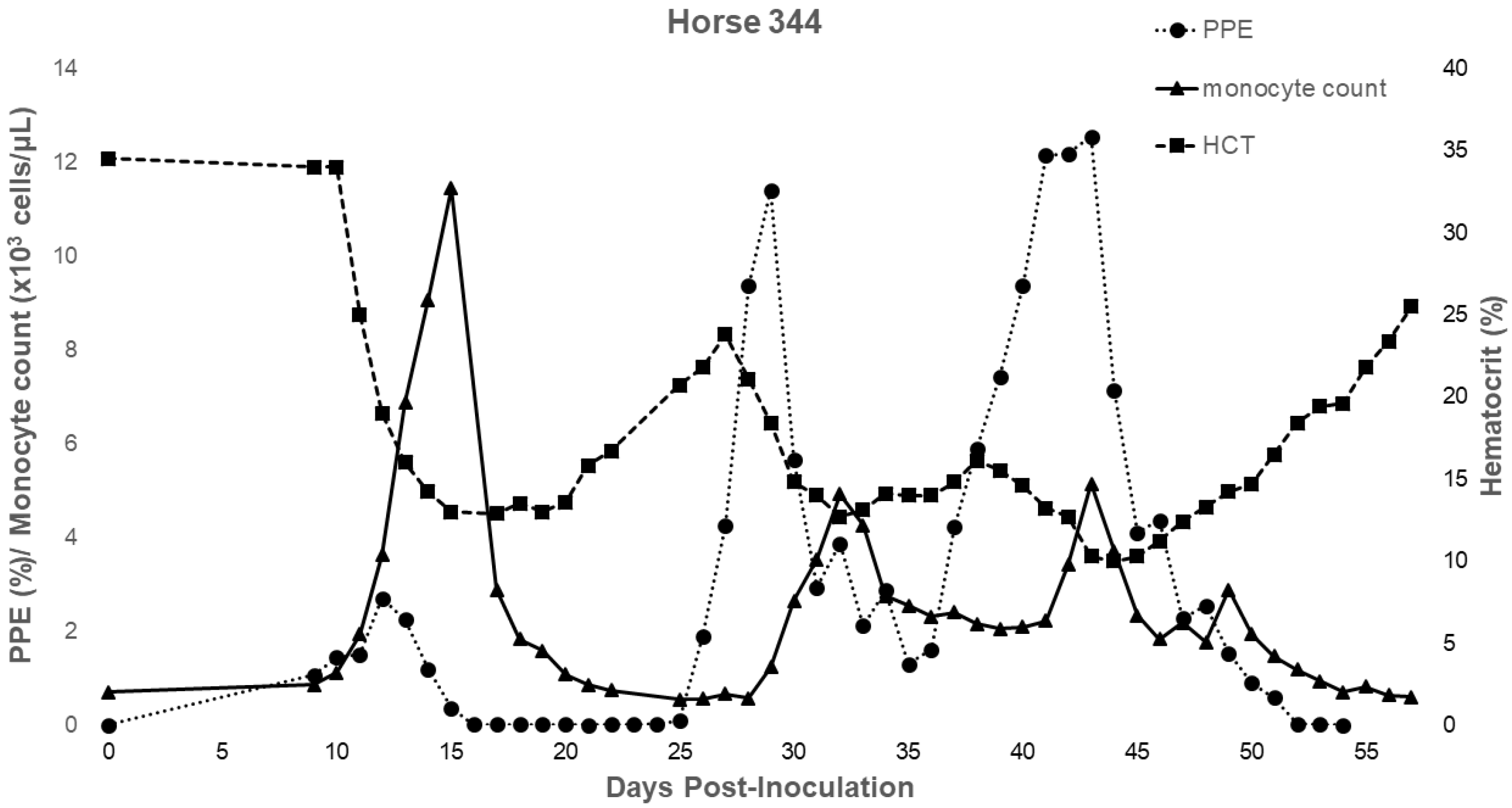
2.2. Characterization of Experimental T. haneyi Infection of Spleen-Intact Horses
2.3. Characterization of Complete Blood Count Parameters in Experimentally Infected Horses
2.4. Monitoring of T. haneyi Parasitemia via nPCR and Blood Smear Cytology
3. Discussion
4. Materials and Methods
4.1. Equine Infection Studies
4.2. Blood Collection
4.3. Light Microscopy
4.4. T. haneyi Nested PCR
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knowles, D.P.; Kappmeyer, L.S.; Haney, D.; Herndon, D.R.; Fry, L.M.; Munro, J.B.; Sears, K.; Ueti, M.W.; Wise, L.N.; Silva, M.; et al. Discovery of a novel species, Theileria haneyi n. sp., infective to equids, highlights exceptional genomic diversity within the genus Theileria: Implications for apicomplexan parasite surveillance. Int. J. Parasitol. 2018, 48, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Wise, L.N.; Kappmeyer, L.S.; Mealey, R.H.; Knowles, D.P. Review of equine piroplasmosis. J. Vet. Intern. Med. 2013, 27, 1334–1346. [Google Scholar] [CrossRef] [PubMed]
- Mshelia, P.W.; Kappmeyer, L.; Johnson, W.C.; Kudi, C.A.; Oluyinka, O.O.; Balogun, E.O.; Richard, E.E.; Onoja, E.; Sears, K.P.; Ueti, M.W. Molecular detection of Theileria species and Babesia caballi from horses in Nigeria. Parasitol. Res. 2020, 119, 2955–2963. [Google Scholar] [CrossRef] [PubMed]
- Coultous, R.M.; McDonald, M.; Raftery, A.G.; Shiels, B.R.; Sutton, D.G.M.; Weir, W. Analysis of Theileria equi diversity in The Gambia using a novel genotyping method. Transbound. Emerg. Dis. 2019, 67, 1213–1221. [Google Scholar] [CrossRef]
- Bhoora, R.V.; Collins, N.E.; Schnittger, L.; Troskie, C.; Marumo, R.; Labuschagne, K.; Smith, R.M.; Dalton, D.L.; Mbizeni, S. Molecular genotyping and epidemiology of equine piroplasmids in South Africa. Ticks Tick Borne Dis. 2019, 11, 101358. [Google Scholar] [CrossRef]
- Zobba, R.; Ardu, M.; Niccolini, S.; Chessa, B.; Manna, L.; Cocco, R.; Pinna Parpaglia, M.L. Clinical and Laboratory Findings in Equine Piroplasmosis. J. Equine Vet. Sci. 2008, 28, 301–308. [Google Scholar] [CrossRef]
- Hailat, N.Q.; Lafi, S.Q.; Al-Darraji, A.M.; Al-Ani, F.K. Equine babesiosis associated with strenuous exercise: Clinical and pathological studies in Jordan. Vet. Parasitol. 1997, 69, 1–8. [Google Scholar] [CrossRef]
- Sears, K.P.; Kappmeyer, L.S.; Wise, L.N.; Silva, M.; Ueti, M.W.; White, S.; Reif, K.E.; Knowles, D.P. Infection dynamics of Theileria equi and Theileria haneyi, a newly discovered apicomplexan of the horse. Vet. Parasitol. 2019, 271, 68–75. [Google Scholar] [CrossRef]
- Wise, L.N.; Kappmeyer, L.S.; Silva, M.G.; White, S.N.; Grause, J.F.; Knowles, D.P. Verification of post-chemotherapeutic clearance of Theileria equi through concordance of nested PCR and immunoblot. Ticks Tick Borne Dis. 2018, 9, 135–140. [Google Scholar] [CrossRef]
- Sears, K.; Knowles, D.; Dinkel, K.; Mshelia, P.W.; Onzere, C.; Silva, M.; Fry, L. Imidocarb Dipropionate Lacks Efficacy against Theileria haneyi and Fails to Consistently Clear Theileria equi in Horses Co-Infected with T. haneyi. Pathogens 2020, 9, 1035. [Google Scholar] [CrossRef]
- Ogier, J.C.; Pagès, S.; Bisch, G.; Chiapello, H.; Médigue, C.; Rouy, Z.; Teyssier, C.; Vincent, S.; Tailliez, P.; Givaudan, A.; et al. Attenuated virulence and genomic reductive evolution in the entomopathogenic bacterial symbiont species, Xenorhabdus poinarii. Genome Biol. Evol. 2014, 6, 1495–1513. [Google Scholar] [CrossRef] [PubMed]
- Diop, A.; Raoult, D.; Fournier, P.E. Rickettsial genomics and the paradigm of genome reduction associated with increased virulence. Microbes Infect. 2018, 20, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A. The Pathogenic Potential of a Microbe. mSphere 2017, 2, e15–e17. [Google Scholar] [CrossRef]
- Lover, A.A.; Coker, R.J. Do mixed infections matter? Assessing virulence of mixed-clone infections in experimental human and murine malaria. Infect. Genet. Evol. 2015, 36, 82–91. [Google Scholar] [CrossRef]
- Mackinnon, M.J.; Read, A.F. Virulence in malaria: An evolutionary viewpoint. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 965–986. [Google Scholar] [CrossRef]
- Schmidt, L.H.; Fradkin, R.; Sesler, C.; Squires, W.; Zeyen, P. Attenuation of the virulence of the M strain of Plasmodium cynomolgi during prolonged multiplication in splenectomized rhesus monkeys. Am. J. Trop. Med. Hyg. 1987, 37, 460–490. [Google Scholar] [CrossRef] [PubMed]
- Ueti, M.W.; Mealey, R.H.; Kappmeyer, L.S.; White, S.N.; Kumpula-McWhirter, N.; Pelzel, A.M.; Grause, J.F.; Bunn, T.O.; Schwartz, A.; Traub-Dargatz, J.L.; et al. Re-Emergence of the apicomplexan Theileria equi in the United States: Elimination of persistent infection and transmission risk. PLoS ONE 2012, 7, e44713. [Google Scholar] [CrossRef]
- Kuttler, K.L.; Zaugg, J.L.; Gipson, C.A. Imidocarb and parvaquone in the treatment of piroplasmosis (Babesia equi) in equids. Am. J. Vet. Res. 1987, 48, 1613–1616. [Google Scholar]
- Zaugg, J.L.; Lane, V.M. Evaluations of buparvaquone as a treatment for equine babesiosis (Babesia equi). Am. J. Vet. Res. 1989, 50, 782–785. [Google Scholar]
- Bach, O.; Baier, M.; Pullwitt, A.; Fosiko, N.; Chagaluka, G.; Kalima, M.; Pfister, W.; Straube, E.; Molyneux, M. Falciparum malaria after splenectomy: A prospective controlled study of 33 previously splenectomized Malawian adults. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 861–867. [Google Scholar] [CrossRef]
- Buffet, P.A.; Safeukui, I.; Deplaine, G.; Brousse, V.; Prendki, V.; Thellier, M.; Turner, G.D.; Mercereau-Puijalon, O. The pathogenesis of Plasmodium falciparum malaria in humans: Insights from splenic physiology. Blood 2011, 117, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Ambawat, H.K.; Malhotra, D.V.; Kumar, S.; Dhar, S. Erythrocyte associated haemato-biochemical changes in Babesia equi infection experimentally produced in donkeys. Vet. Parasitol. 1999, 85, 319–324. [Google Scholar] [CrossRef]
- Guimarães, A.M.; Lima, J.D.; Tafuri, W.L.; Ribeiro, M.F.B.; Sciavicco, C.J.S.; Botelho, A.C.C. Clinical and histopathological aspects of splenectomized foals infected by Babesia equi. J. Equine Vet. Sci. 1997, 17, 211–216. [Google Scholar] [CrossRef]
- Singh, B.; Banerjee, D.P.; Gautam, O.P.; Gupta, R.K.P. Clinicopathological changes in splenectomised donkeys infected with Babesia equi. Indian J. Parasitol. 1980, 4, 77–80. [Google Scholar]
- Knowles, D.P., Jr.; Kappmeyer, L.S.; Perryman, L.E. Specific immune responses are required to control parasitemia in Babesia equi infection. Infect. Immun. 1994, 62, 1909–1913. [Google Scholar] [CrossRef]
- White, N.J. Malaria parasite clearance. Malar. J. 2017, 16, 88. [Google Scholar] [CrossRef]
- Kuttler, K.L.; Gipson, C.A.; Goff, W.L.; Johnson, L.W. Experimental Babesia equi infection in mature horses. Am. J. Vet. Res. 1986, 47, 1668–1670. [Google Scholar]
- Cunha, C.W.; McGuire, T.C.; Kappmeyer, L.S.; Hines, S.A.; Lopez, A.M.; Dellagostin, O.A.; Knowles, D.P. Development of specific immunoglobulin Ga (IgGa) and IgGb antibodies correlates with control of parasitemia in Babesia equi Infection. Clin. Vaccine Immunol. 2006, 13, 297–300. [Google Scholar] [CrossRef]
- Teuscher, F.; Gatton, M.L.; Chen, N.; Peters, J.; Kyle, D.E.; Cheng, Q. Artemisinin-Induced Dormancy in Plasmodium falciparum: Duration, Recovery Rates, and Implications in Treatment Failure. J. Infect. Dis. 2010, 202, 1362–1368. [Google Scholar] [CrossRef]
- Bediako, Y.; Ngoi, J.M.; Nyangweso, G.; Wambua, J.; Opiyo, M.; Nduati, E.W.; Bejon, P.; Marsh, K.; Ndungu, F.M. The effect of declining exposure on T cell-mediated immunity to Plasmodium falciparum–an epidemiological “natural experiment”. BMC Med. 2016, 14, 143. [Google Scholar] [CrossRef]
- Nahrendorf, W.; Ivens, A.; Spence, P.J. Inducible mechanisms of disease tolerance provide an alternative strategy of acquired immunity to malaria. eLife 2021, 10, e63838. [Google Scholar] [CrossRef] [PubMed]
- Fry, L.M.; Schneider, D.A.; Frevert, C.W.; Nelson, D.D.; Morrison, W.I.; Knowles, D.P. East Coast Fever Caused by Theileria parva Is Characterized by Macrophage Activation Associated with Vasculitis and Respiratory Failure. PLoS ONE 2016, 11, e0156004. [Google Scholar] [CrossRef]
- Kho, S.; Barber, B.E.; Johar, E.; Andries, B.; Poespoprodjo, J.R.; Kenangalem, E.; Piera, K.A.; Ehmann, A.; Price, R.N.; William, T.; et al. Platelets kill circulating parasites of all major Plasmodium species in human malaria. Blood 2018, 132, 1332–1344. [Google Scholar] [CrossRef] [PubMed]
- Nafie, T.H.; El-Allawy, T.; Mottelib, A.A.; EL-Amrousi, S. Effect of induced equine piroplasmosis on some blood constituents. Assiut Vet. Med. J. 1982, 9, 123–128. [Google Scholar]
- Rudolph, W.; Correa, J.; Zurita, L.; Manley, W. Equine piroplasmosis: Leukocytic response to Babesia equi (Laveran, 1901) infection in Chile. Br. Vet. J. 1975, 131, 601–609. [Google Scholar] [CrossRef]
- Hviid, L.; Kemp, K. What is the cause of lymphopenia in malaria? Infect. Immun. 2000, 68, 6087–6089. [Google Scholar] [CrossRef]
- Aitken, E.H.; Alemu, A.; Rogerson, S.J. Neutrophils and Malaria. Front. Immunol. 2018, 9, 3005. [Google Scholar] [CrossRef]
- Juneja, S.; Januszewicz, E.; Wolf, M.; Cooper, I. Post-Splenectomy lymphocytosis. Clin. Lab. Haematol. 1995, 17, 335–337. [Google Scholar]
- Dürig, M.; Landmann, R.M.; Harder, F. Lymphocyte subsets in human peripheral blood after splenectomy and autotransplantation of splenic tissue. J. Lab. Clin. Med. 1984, 104, 110–115. [Google Scholar]
- Sponaas, A.M.; Freitas do Rosario, A.P.; Voisine, C.; Mastelic, B.; Thompson, J.; Koernig, S.; Jarra, W.; Renia, L.; Mauduit, M.; Potocnik, A.J.; et al. Migrating monocytes recruited to the spleen play an important role in control of blood stage malaria. Blood 2009, 114, 5522–5531. [Google Scholar] [CrossRef]
- Court, R.A.; Jackson, L.A.; Lee, R.P. Elevated anti-parasitic activity in peripheral blood monocytes and neutrophils of cattle infected with Babesia bovis. Int. J. Parasitol. 2001, 31, 29–37. [Google Scholar] [CrossRef]
- Klei, T.R.; Meinderts, S.M.; van den Berg, T.K.; van Bruggen, R. From the Cradle to the Grave: The Role of Macrophages in Erythropoiesis and Erythrophagocytosis. Front. Immunol. 2017, 8, 73. [Google Scholar] [CrossRef] [PubMed]
- Duez, J.; Holleran, J.P.; Ndour, P.A.; Pionneau, C.; Diakité, S.; Roussel, C.; Dussiot, M.; Amireault, P.; Avery, V.M.; Buffet, P.A. Mechanical clearance of red blood cells by the human spleen: Potential therapeutic applications of a biomimetic RBC filtration method. Transfus. Clin. Biol. 2015, 22, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.C.; Nelson, C.S.; Wilson, E.B.; Hou, B.; DeFranco, A.L.; DeRisi, J.L. Splenic red pulp macrophages produce type I interferons as early sentinels of malaria infection but are dispensable for control. PLoS ONE 2012, 7, e48126. [Google Scholar] [CrossRef] [PubMed]
- Stijlemans, B.; Cnops, J.; Naniima, P.; Vaast, A.; Bockstal, V.; De Baetselier, P.; Magez, S. Development of a pHrodo-based assay for the assessment of in vitro and in vivo erythrophagocytosis during experimental trypanosomosis. PLoS Negl. Trop. Dis. 2015, 9, e0003561. [Google Scholar] [CrossRef] [PubMed]
- Erhart, L.M.; Yingyuen, K.; Chuanak, N.; Buathong, N.; Laoboonchai, A.; Miller, R.S.; Meshnick, S.R.; Gasser, R.A., Jr.; Wongsrichanalai, C. Hematologic and clinical indices of malaria in a semi-immune population of western Thailand. Am. J. Trop. Med. Hyg. 2004, 70, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Gérardin, P.; Rogier, C.; Ka, A.S.; Jouvencel, P.; Brousse, V.; Imbert, P. Prognostic value of thrombocytopenia in African children with falciparum malaria. Am. J. Trop. Med. Hyg. 2002, 66, 686–691. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nwaneri, D.; Oladipo, O.; Ifebi, E.; Oviawe, O.; Asemota, O.; Ogboghodo, B.; Israel-Aina, Y.; Sadoh, A. Haematological Parameters and Spleen Rate of Asymptomatic and Malaria Negative Children in Edo South District, Nigeria. Ann. Glob. Health 2020, 86, 62. [Google Scholar] [CrossRef]
- Kini, R.G.; Chandrashekhar, J. Parasite and the Circulating Pool-Characterisation of Leukocyte Number and Morphology in Malaria. J. Clin. Diagn. Res. 2016, 10, Ec44–Ec48. [Google Scholar] [CrossRef]
- Dobbs, K.R.; Embury, P.; Vulule, J.; Odada, P.S.; Rosa, B.A.; Mitreva, M.; Kazura, J.W.; Dent, A.E. Monocyte dysregulation and systemic inflammation during pediatric falciparum malaria. JCI Insight 2017, 2, e95352. [Google Scholar] [CrossRef]
- Ortega-Pajares, A.; Rogerson, S.J. The Rough Guide to Monocytes in Malaria Infection. Front. Immunol. 2018, 9, 2888. [Google Scholar] [CrossRef] [PubMed]
- Bastos, R.G.; Sears, K.; Dinkel, K.D.; Knowles, D.P.; Fry, L.M. Changes in the Molecular and Functional Phenotype of Bovine Monocytes during Theileria parva Infection. Infect. Immun. 2019, 87, e00703-19. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Horse | Group | Inoculation Type | Stabilate Volume and PPE | Splenectomy Pre- or Post-Inoculation | Associated Study |
|---|---|---|---|---|---|
| 248 | 1 | Blood—120 mL | 2 mL/12% (donor) | Pre | [1] |
| 301 | 1 | Blood—120 mL | 2 mL/12% (donor) | Pre | [1] |
| 285 | 1 | Stabilate | 5 mL/8.8% | Pre | This study |
| 344 | 1 | Blood—150 mL | 4 mL/8.8% (donor) | Pre | This study |
| 275 | 2 | Stabilate | 2 mL/12% | Post | [8,10] |
| 277 | 2 | Stabilate | 2 mL/12% | Post | [8,10] |
| 278 | 2 | Stabilate | 2 mL/12% | Post | [8,10] |
| 280 | 2 | Stabilate | 2 mL/12% | Post | [8,10] |
| Horse | 248 | 285 | 301 | 344 |
|---|---|---|---|---|
| PPE peaks | 15 dpi—3.75% 32 dpi—7.32% | 23 dpi—0.65% 54 dpi—4.1% 69 dpi—2.34% | 14 dpi—1.03% 30 dpi—1.08% 50 dpi—1.06% | 13 dpi—3.10% 29 dpi—11.40% 43 dpi—12.55% |
| PCV nadirs | 20 dpi—18% 37 dpi—10% | 29 dpi—18% 54 dpi—20% 72 dpi—15% | 21 dpi—33% 38 dpi—27% 51 dpi—23% | 15—13% 32—12.7% 44—10% |
| Maximum% change in PCV # | −72.22% | −57.14% # | −53.06% | −71.01% |
| Anemia duration | 15–37 dpi | 23–87 dpi | 35–67 dpi | 11–62 |
| Survival | Euthanized 37 dpi | survived | survived | Survived |
| Hemogram Changes | 344 (dpi) | 275 (dps) | 277 (dps) | 278 (dps) | 280 (dps) |
|---|---|---|---|---|---|
| Monocytosis | 11–22, 30–57 | 15–28 | 14–24 | 13–28 | 15–26 |
| Lymphocytosis | 11–67 | None | 15–27, 29 | 19, 21–31 | 20–24,27 |
| Lymphopenia | None | 1–3 | None | None | None |
| Neutropenia | 30–39, 42–51 | 1–3 | None | None | 2 |
| Neutrophilia | 28 | 7, 9–10 | 3, 6–10, 19–25 | 6–29 | 9–11, 13–14, 20–21, 23–30 |
| Anemia duration | 11–62 | 20–42 | 17–32 | 18–34 | 18–43 |
| Anemia nadir | 15, 32, 44 | 31 | 22 | 24 | 24 |
| Maximum% change in HCT | −71.01% | −33.52% | −66.31% | −76.53% | −55.72% |
| Parasitemia peak(s) | 13, 29, 43 | 14 | 17 | 19 | 17 |
| Horse | Survival Acute Infection | Long-Term Survival | nPCR 6–12 Months Post-Infection/Splenectomy |
|---|---|---|---|
| 248 | N | N | Positive—37 dpi |
| 301 | Y | Euthanized; chronic renal failure | Positive—1402 dpi |
| 285 | Y | Euthanized; guttural pouch empyema and acute, severe laminitis | Positive—556 dpi |
| 344 | Y | Y | No longer detected; last positive 267 days post-inoculation; negative at 350 dpi and last negative at 1565 dpi |
| 275 | Y | Y | Positive; 536 days post-splenectomy |
| 277 | Y | Y | No longer detected after 106 dps (1/17/20), then neg on 3/3, 5/2, and 7/24, faint pos on 9/23, then neg 11/17, and 1/5/21 |
| 278 | Y | Y | No longer detected after 152 dps |
| 280 | Y | Y | No longer detected after 171 dps |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sears, K.P.; Knowles, D.P.; Fry, L.M. Clinical Progression of Theileria haneyi in Splenectomized Horses Reveals Decreased Virulence Compared to Theileria equi. Pathogens 2022, 11, 254. https://doi.org/10.3390/pathogens11020254
Sears KP, Knowles DP, Fry LM. Clinical Progression of Theileria haneyi in Splenectomized Horses Reveals Decreased Virulence Compared to Theileria equi. Pathogens. 2022; 11(2):254. https://doi.org/10.3390/pathogens11020254
Chicago/Turabian StyleSears, Kelly P., Donald P. Knowles, and Lindsay M. Fry. 2022. "Clinical Progression of Theileria haneyi in Splenectomized Horses Reveals Decreased Virulence Compared to Theileria equi" Pathogens 11, no. 2: 254. https://doi.org/10.3390/pathogens11020254
APA StyleSears, K. P., Knowles, D. P., & Fry, L. M. (2022). Clinical Progression of Theileria haneyi in Splenectomized Horses Reveals Decreased Virulence Compared to Theileria equi. Pathogens, 11(2), 254. https://doi.org/10.3390/pathogens11020254