
Transcriptomic Analysis of Fish Hosts Responses to Nervous Necrosis Virus




Abstract
:1. Introduction
2. Nervous Necrosis Virus
3. Results
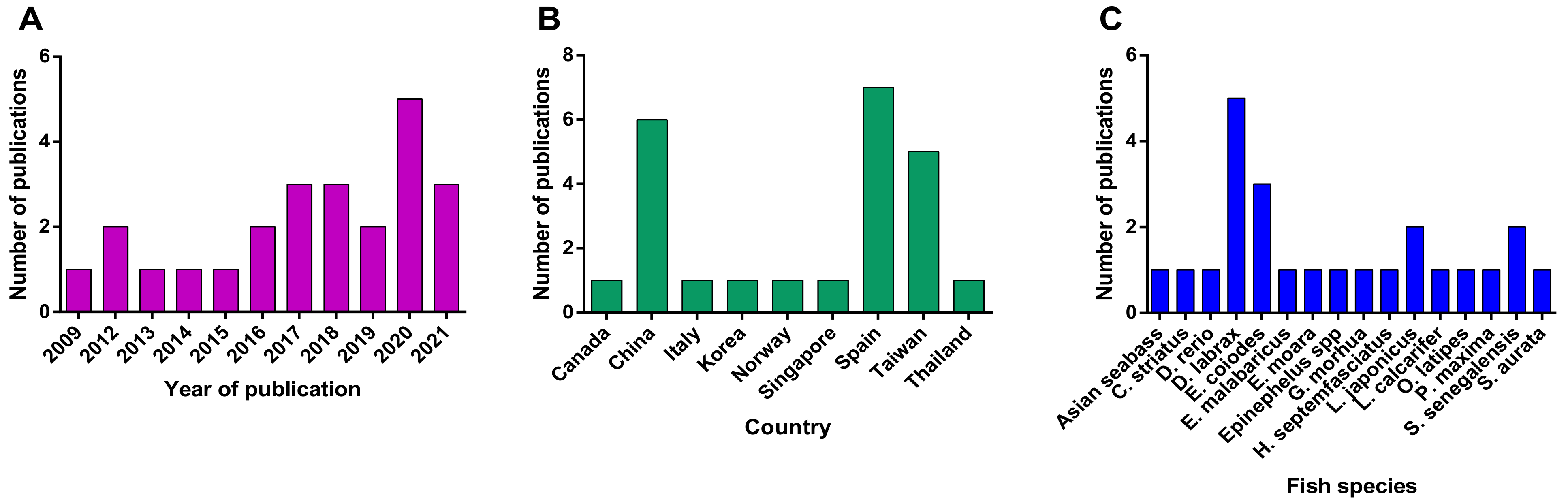
3.1. Search Strategy and Outcomes Analysis
3.2. Transcriptome Analysis Platform Information
3.3. Bioinformatic Methods for Transciptomic Data Analysis
4. Discussion
4.1. Kidney
4.2. Spleen
4.3. Liver
4.4. Brain
4.5. Larvae
4.6. Cell Lines
4.7. Pathway Analysis
5. Materials and Methods
5.1. Search Strategy
5.2. Study Selection Process and Eligibility Criteria
5.3. Data Extraction
5.4. Gene Name Standardization
5.5. Phylogenetic Analysis
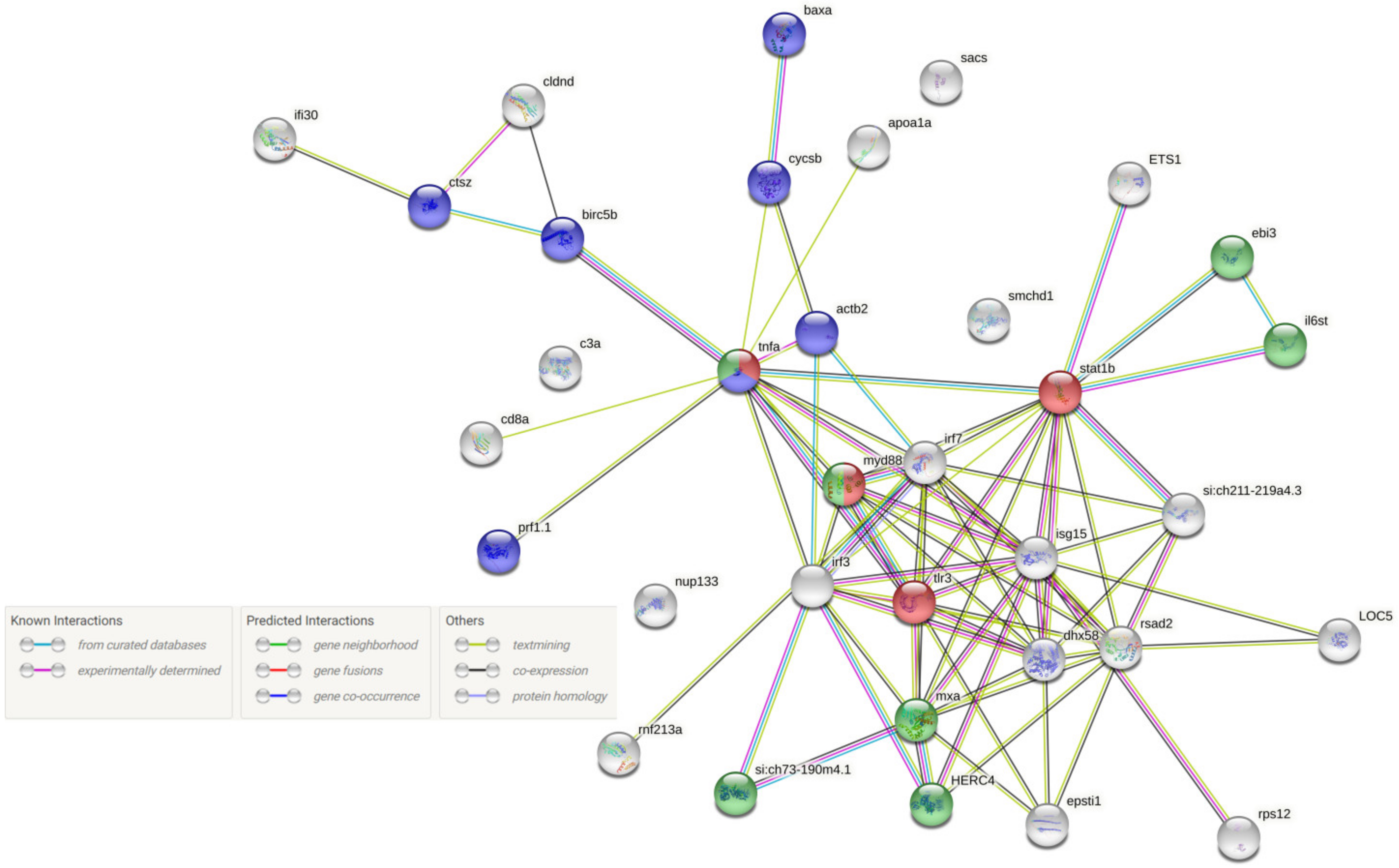
5.6. Construction of the PPI Network
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmed, N.; Thompson, S. The blue dimensions of aquaculture: A global synthesis. Sci. Total Environ. 2019, 652, 851–861. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2020. In Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Natnan, M.E.; Low, C.F.; Chong, C.M.; Bunawan, H.; Baharum, S.N. Integration of Omics Tools for Understanding the Fish Immune Response Due to Microbial Challenge. Front. Mar. Sci. 2021, 8, 668771. [Google Scholar] [CrossRef]
- Dadar, M.; Dhama, K.; Vakharia, V.N.; Hoseinifar, S.H.; Karthik, K.; Tiwari, R.; Khandia, R.; Munjal, A.; Salgado-Miranda, C.; Joshi, S.K. Advances in Aquaculture Vaccines Against Fish Pathogens: Global Status and Current Trends. Rev. Fish. Sci. Aquac. 2017, 25, 184–217. [Google Scholar] [CrossRef]
- Moreno, P.; Gemez-Mata, J.; Garcia-Rosado, E.; Bejar, J.; Labella, A.M.; Souto, S.; Alonso, M.C. Differential immunogene expression profile of European sea bass (Dicentrarchus labrax, L.) in response to highly and low virulent NNV. Fish Shellfish Immunol. 2020, 106, 56–70. [Google Scholar] [CrossRef]
- Ye, H.; Lin, Q.; Luo, H. Applications of transcriptomics and proteomics in understanding fish immunity. Fish Shellfish Immunol. 2018, 77, 319–327. [Google Scholar] [CrossRef]
- Sudhagar, A.; Kumar, G.; El-Matbouli, M. Transcriptome Analysis Based on RNA-Seq in Understanding Pathogenic Mechanisms of Diseases and the Immune System of Fish: A Comprehensive Review. Int. J. Mol. Sci. 2018, 19, 245. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.W.; Tso, C.H.; Periyasamy, T. Studies on Nervous Necrosis Virus Pathogenesis by Transcriptomic Approaches. In Marine OMICS: Principles and Applications; Kim, S.K., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 297–306. [Google Scholar]
- Maekawa, S.; Wang, P.C.; Chen, S.C. Comparative Study of Immune Reaction against Bacterial Infection from Transcriptome Analysis. Front. Immunol. 2019, 10, 153. [Google Scholar] [CrossRef]
- Chandhini, S.; Kumar, V.J.R. Transcriptomics in aquaculture: Current status and applications. Rev. Aquacult. 2019, 11, 1379–1397. [Google Scholar] [CrossRef]
- Munday, B.L.; Kwang, J.; Moody, N. Betanodavirus infections of teleost fish: A review. J. Fish Dis. 2002, 25, 127–142. [Google Scholar] [CrossRef]
- Bandín, I.; Souto, S. Betanodavirus and VER Disease: A 30-year Research Review. Pathogens 2020, 9, 106. [Google Scholar] [CrossRef] [Green Version]
- Doan, Q.K.; Vandeputte, M.; Chatain, B.; Morin, T.; Allal, F. Viral encephalopathy and retinopathy in aquaculture: A review. J. Fish Dis. 2016, 40, 717–742. [Google Scholar] [CrossRef] [Green Version]
- Das, P.K.; Salinas, I. Fish nasal immunity: From mucosal vaccines to neuroimmunology. Fish Shellfish Immunol. 2020, 104, 165–171. [Google Scholar] [CrossRef]
- Toledo-Ibarra, G.; Rojas-Mayorquín, A.E.; Girón-Pérez, M.I. Influence of the cholinergic system on the immune response of teleost fishes: Potential model in biomedical research. Clin. Dev. Immunol. 2013, 2013, 536534. [Google Scholar] [CrossRef]
- Gonzalez-Silvera, D.; Guardiola, F.A.; Espinosa, C.; Chaves-Pozo, E.; Esteban, M.Á.; Cuesta, A. Recombinant nodavirus vaccine produced in bacteria and administered without purification elicits humoral immunity and protects European sea bass against infection. Fish Shellfish Immunol. 2019, 88, 458–463. [Google Scholar] [CrossRef]
- Navaneeth Krishnan, A.; Bhuvaneswari, T.; Ezhil Praveena, P.; Jithendran, K.P. Paper-based archiving of biological samples from fish for detecting betanodavirus. Arch. Virol. 2016, 161, 2019–2024. [Google Scholar] [CrossRef]
- Costa, J.Z.; Thompson, K.D. Understanding the interaction between Betanodavirus and its host for the development of prophylactic measures for viral encephalopathy and retinopathy. Fish Shellfish Immunol. 2016, 53, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Nagai, T.; Nishizawa, T. Sequence of the non-structural protein gene encoded by RNA1 of striped jack nervous necrosis virus. J. Gen. Virol. 1999, 80, 3019–3022. [Google Scholar] [CrossRef]
- Nishizawa, T.; Mori, K.; Nakai, T.; Furusawa, I.; Muroga, K. Polymerase chain reaction (PCR) amplification of RNA of striped jack nervous necrosis virus (SJNNV). Dis. Aquat. Organ. 1994, 18, 103–107. [Google Scholar] [CrossRef]
- Iwamoto, T.; Mise, K.; Takeda, A.; Okinaka, Y.; Mori, K.I.; Arimoto, M.; Okuno, T.; Nakai, T. Characterization of Striped jack nervous necrosis virus subgenomic RNA3 and biological activities of its encoded protein B2. J. Gen. Virol. 2005, 86, 2807–2816. [Google Scholar] [CrossRef]
- Chen, L.J.; Su, Y.C.; Hong, J.R. Betanodavirus non-structural protein B1: A novel anti-necrotic death factor that modulates cell death in early replication cycle in fish cells. Virology 2009, 385, 444–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishizawa, T.; Furuhashi, M.; Nagai, T.; Nakai, T.; Muroga, K. Genomic classification of fish nodaviruses by molecular phylogenetic analysis of the coat protein gene. Appl. Environ. Microbiol. 1997, 63, 1633–1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olveira, J.G.; Souto, S.; Dopazo, C.P.; Thiéry, R.; Barja, J.L.; Bandín, I. Comparative analysis of both genomic segments of betanodaviruses isolated from epizootic outbreaks in farmed fish species provides evidence for genetic reassortment. J. Gen. Virol. 2009, 90, 2940–2951. [Google Scholar] [CrossRef] [PubMed]
- Toffan, A.; Pascoli, F.; Pretto, T.; Panzarin, V.; Abbadi, M.; Buratin, A.; Quartesan, R.; Gijón, D.; Padrós, F. Viral nervous necrosis in gilthead sea bream (Sparus aurata) caused by reassortant betanodavirus RGNNV/SJNNV: An emerging threat for Mediterranean aquaculture. Sci. Rep. 2017, 7, 46755. [Google Scholar] [CrossRef] [PubMed]
- Thiéry, R.; Cozien, J.; de Boisséson, C.; Kerbart-Boscher, S.; Névarez, L. Genomic classification of new betanodavirus isolates by phylogenetic analysis of the coat protein gene suggests a low host-fish species specificity. J. Gen. Virol. 2004, 85, 3079–3087. [Google Scholar] [CrossRef] [PubMed]
- Panzarin, V.; Fusaro, A.; Monne, I.; Cappellozza, E.; Patarnello, P.; Bovo, G.; Capua, I.; Holmes, E.C.; Cattoli, G. Molecular epidemiology and evolutionary dynamics of betanodavirus in southern Europe. Infect. Genet. Evol. 2012, 12, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Park, K.C.; Osborne, J.A.; Montes, A.; Dios, S.; Nerland, A.H.; Novoa, B.; Figueras, A.; Brown, L.L.; Johnson, S.C. Immunological responses of turbot (Psetta maxima) to nodavirus infection or polyriboinosinic polyribocytidylic acid (pIC) stimulation, using expressed sequence tags (ESTs) analysis and cDNA microarrays. Fish Shellfish Immunol. 2009, 26, 91–108. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.W.; Ngou, F.H.; Chao, Y.M.; Lai, Y.S.; Chen, N.Y.; Lee, F.Y.; Chiou, P.P. Transcriptome characterization and gene expression of Epinephelus spp. in endoplasmic reticulum stress-related pathway during betanodavirus infection in vitro. BMC Genom. 2012, 13, 651. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.S.; Chen, C.W.; Lin, C.H.; Tzeng, C.S.; Chang, C.Y. Differential expression profiling of orange-spotted grouper larvae, Epinephelus coioides (Hamilton), that survived a betanodavirus outbreak. J. Fish Dis. 2012, 35, 215–225. [Google Scholar] [CrossRef]
- Krasnov, A.; Kileng, Ø.; Skugor, S.; Jørgensen, S.M.; Afanasyev, S.; Timmerhaus, G.; Sommer, A.I.; Jensen, I. Genomic analysis of the host response to nervous necrosis virus in Atlantic cod (Gadus morhua) brain. Mol. Immunol. 2013, 54, 443–452. [Google Scholar] [CrossRef]
- Wang, Y.D.; Rajanbabu, V.; Chen, J.Y. Transcriptome analysis of medaka following epinecidin-1 and TH1-5 treatment of NNV infection. Fish Shellfish Immunol. 2015, 42, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Liu, W.; Wu, S.Y.; Chiou, P.P.; Li, Y.H.; Chen, Y.C.; Lin, G.H.; Lu, M.W.; Wu, J.L. RIG-I specifically mediates group II type I IFN activation in nervous necrosis virus infected zebrafish cells. Fish Shellfish Immunol. 2015, 43, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wang, L.; Kwang, J.; Yue, G.H.; Wong, S.M. Transcriptome analysis of genes responding to NNV infection in Asian seabass epithelial cells. Fish Shellfish Immunol. 2016, 54, 342–352. [Google Scholar] [CrossRef]
- Chen, W.; Yi, L.; Feng, S.; Liu, X.; Asim, M.; Zhou, Y.; Lan, J.; Jiang, S.; Tu, J.; Lin, L. Transcriptomic profiles of striped snakehead fish cells (SSN-1) infected with red-spotted grouper nervous necrosis virus (RGNNV) with an emphasis on apoptosis pathway. Fish Shellfish Immunol. 2017, 60, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Pozo, E.; Valero, Y.; Esteve-Codina, A.; Gómez-Garrido, J.; Dabad, M.; Alioto, T.; Meseguer, J.; Esteban, M.Á.; Cuesta, A. Innate Cell-Mediated Cytotoxic Activity of European Sea Bass Leucocytes Against Nodavirus-Infected Cells: A Functional and RNA-seq Study. Sci. Rep. 2017, 7, 15396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.O.; Kim, J.O.; Kim, W.S.; Oh, M.J. Characterization of the Transcriptome and Gene Expression of Brain Tissue in Sevenband Grouper (Hyporthodus septemfasciatus) in Response to NNV Infection. Genes 2017, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Yi, L.; Feng, S.; Zhao, L.; Li, J.; Zhou, M.; Liang, R.; Gu, N.; Wu, Z.; Tu, J.; et al. Characterization of microRNAs in orange-spotted grouper (Epinephelus coioides) fin cells upon red-spotted grouper nervous necrosis virus infection. Fish Shellfish Immunol. 2017, 63, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Jia, P.; Liu, W.; Yi, M.; Jia, K. Comparative transcriptome analysis reveals the role of p53 signalling pathway during red-spotted grouper nervous necrosis virus infection in Lateolabrax japonicus brain cells. J. Fish Dis. 2019, 42, 585–595. [Google Scholar] [CrossRef]
- Labella, A.M.; Garcia-Rosado, E.; Bandín, I.; Dopazo, C.P.; Castro, D.; Alonso, M.C.; Borrego, J.J. Transcriptomic Profiles of Senegalese Sole Infected with Nervous Necrosis Virus Reassortants Presenting Different Degree of Virulence. Front. Immunol. 2018, 9, 1626. [Google Scholar] [CrossRef]
- Tso, C.H.; Lu, M.W. Transcriptome profiling analysis of grouper during nervous necrosis virus persistent infection. Fish Shellfish Immunol. 2018, 76, 224–232. [Google Scholar] [CrossRef]
- Chaves-Pozo, E.; Bandín, I.; Olveira, J.G.; Esteve-Codina, A.; Gómez-Garrido, J.; Dabad, M.; Alioto, T.; Ángeles Esteban, M.; Cuesta, A. European sea bass brain DLB-1 cell line is susceptible to nodavirus: A transcriptomic study. Fish Shellfish Immunol. 2019, 86, 14–24. [Google Scholar] [CrossRef]
- Wang, L.; Tian, Y.; Cheng, M.; Li, Z.; Li, S.; Wu, Y.; Zhang, J.; Ma, W.; Li, W.; Pang, Z.; et al. Transcriptome comparative analysis of immune tissues from asymptomatic and diseased Epinephelus moara naturally infected with nervous necrosis virus. Fish Shellfish Immunol. 2019, 93, 99–107. [Google Scholar] [CrossRef]
- Lama, R.; Pereiro, P.; Valenzuela-Muñoz, V.; Gallardo-Escárate, C.; Tort, L.; Figueras, A.; Novoa, B. RNA-Seq analysis of European sea bass (Dicentrarchus labrax L.) infected with nodavirus reveals powerful modulation of the stress response. Vet. Res. 2020, 51, 64. [Google Scholar] [CrossRef]
- Pereiro, P.; Lama, R.; Moreira, R.; Valenzuela-Muñoz, V.; Gallardo-Escárate, C.; Novoa, B.; Figueras, A. Potential Involvement of lncRNAs in the Modulation of the Transcriptome Response to Nodavirus Challenge in European Sea Bass (Dicentrarchus labrax L.). Biology 2020, 9, 165. [Google Scholar] [CrossRef]
- Angsujinda, K.; Plaimas, K.; Smith, D.R.; Kettratad, J.; Assavalapsakul, W. Transcriptomic analysis of red-spotted grouper nervous necrosis virus infected Asian seabass Lates calcarifer (Bloch, 1790). Aquac. Rep. 2020, 18, 100517. [Google Scholar] [CrossRef]
- Ge, H.; Lin, K.; Zhou, C.; Lin, Q.; Zhang, Z.; Wu, J.; Zheng, L.; Yang, Q.; Wu, S.; Chen, W.; et al. A multi-omic analysis of orange-spotted grouper larvae infected with nervous necrosis virus identifies increased adhesion molecules and collagen synthesis in the persistent state. Fish Shellfish Immunol. 2020, 98, 595–604. [Google Scholar] [CrossRef]
- Gémez-Mata, J.; Labella, A.M.; Bandín, I.; Borrego, J.J.; García-Rosado, E. Immunogene expression analysis in betanodavirus infected-Senegalese sole using an OpenArray® platform. Gene 2021, 774, 145430. [Google Scholar] [CrossRef]
- Peruzza, L.; Pascoli, F.; Dalla Rovere, G.; Franch, R.; Ferraresso, S.; Babbucci, M.; Biasini, L.; Abbadi, M.; Panzarin, V.; Toffan, A.; et al. Transcriptome analysis reveals a complex response to the RGNNV/SJNNV reassortant Nervous Necrosis Virus strain in sea bream larvae. Fish Shellfish Immunol. 2021, 114, 282–292. [Google Scholar] [CrossRef]
- Jia, P.; Pan, H.; Cui, K.; Jia, K.; Yi, M. MicroRNA expression profiling of sea perch brain cells reveals the roles of microRNAs in autophagy induced by RGNNV infection. J. Fish Dis. 2021, 44, 1305–1314. [Google Scholar] [CrossRef]
- Caruffo, M.; Mandakovic, D.; Cabrera, P.; Pacheco, I.; Montt, L.; Chávez-Báez, I.; Mejías, M.; Vera-Tamargo, F.; Perez-Valenzuela, J.; Carrasco-Labra, A.; et al. Insights into gene expression responses to infections in teleosts using microarray data: A systematic review. Rev. Aquacult. 2021, 13, 18–42. [Google Scholar] [CrossRef]
- Tortorella, D.; Gewurz, B.E.; Furman, M.H.; Schust, D.J.; Ploegh, H.L. Viral subversion of the immune system. Annu. Rev. Immunol. 2000, 18, 861–926. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langevin, C.; Aleksejeva, E.; Passoni, G.; Palha, N.; Levraud, J.P.; Boudinot, P. The antiviral innate immune response in fish: Evolution and conservation of the IFN system. J. Mol. Biol. 2013, 425, 4904–4920. [Google Scholar] [CrossRef]
- Zhou, Z.; Lin, Z.; Pang, X.; Shan, P.; Wang, J. MicroRNA regulation of Toll-like receptor signaling pathways in teleost fish. Fish Shellfish Immunol. 2018, 75, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Rauta, P.R.; Samanta, M.; Dash, H.R.; Nayak, B.; Das, S. Toll-like receptors (TLRs) in aquatic animals: Signaling pathways, expressions and immune responses. Immunol. Lett. 2014, 158, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Munang’andu, H.M.; Evensen, Ø. A Review of Intra- and Extracellular Antigen Delivery Systems for Virus Vaccines of Finfish. J. Immunol. Res. 2015, 60859. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Zhu, Z.; Ge, H.; Zheng, L.; Huang, Z.; Wu, S. Immunity to nervous necrosis virus infections of orange-spotted grouper (Epinephelus coioides) by vaccination with virus-like particles. Fish Shellfish Immunol. 2016, 56, 136–143. [Google Scholar] [CrossRef]
- Souto, S.; Olveira, J.G.; Vázquez-Salgado, L.; Dopazo, C.P.; Bandín, I. Betanodavirus infection in primary neuron cultures from sole. Vet. Res. 2018, 49, 86. [Google Scholar] [CrossRef] [Green Version]
- Chiang, Y.H.; Wu, Y.C.; Chi, S.C. Interleukin-1β secreted from betanodavirus-infected microglia caused the death of neurons in giant grouper brains. Dev. Comp. Immunol. 2017, 70, 19–26. [Google Scholar] [CrossRef]
- Forrester, J.V.; Xu, H.; Lambe, T.; Cornall, R. Immune privilege or privileged immunity? Mucosal. Immunol. 2008, 1, 372–781. [Google Scholar] [CrossRef]
- Forrester, J.V.; McMenamin, P.G.; Dando, S.J. CNS infection and immune privilege. Nat. Rev. Neurosci. 2018, 19, 655–671. [Google Scholar] [CrossRef]
- Li, Y.; Li, C.; Xue, P.; Zhong, B.; Mao, A.P.; Ran, Y.; Chen, H.; Wang, Y.Y.; Yang, F.; Shu, H.B. ISG56 is a negative-feedback regulator of virus-triggered signaling and cellular antiviral response. Proc. Natl. Acad. Sci. USA 2009, 106, 7945–7950. [Google Scholar] [CrossRef] [Green Version]
- Mankouri, J.; Fragkoudis, R.; Richards, K.H.; Wetherill, L.F.; Harris, M.; Kohl, A.; Elliott, R.M.; Macdonald, A. Optineurin negatively regulates the induction of IFNbeta in response to RNA virus infection. PLoS Pathog. 2010, 6, e1000778. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Liao, J.; Meyerdierks, A.; Feng, L.; Naumovski, L.; Bottger, E.C.; Omary, M.B. Interferon-alpha induces nmi-IFP35 heterodimeric complex formation that is affected by the phosphorylation of IFP35. J. Biol. Chem. 2000, 275, 21364–21371. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Naumovski, L. Intracellular redistribution of interferon-inducible proteins Nmi and IFP 35 in apoptotic cells. J. Interferon Cytokine Res. 2002, 22, 237–243. [Google Scholar] [CrossRef]
- Das, A.; Dinh, P.X.; Panda, D.; Pattnaik, A.K. Interferon-inducible protein IFI35 negatively regulates RIG-I antiviral signaling and supports vesicular stomatitis virus replication. J. Virol. 2014, 88, 3103–3113. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Qin, Z.; Babu, V.S.; Zhao, L.; Li, J.; Zhang, X.; Lin, L. Transcriptomic profiles of striped snakehead cells (SSN-1) infected with snakehead vesiculovirus (SHVV) identifying IFI35 as a positive factor for SHVV replication. Fish Shellfish Immunol. 2019, 86, 46–52. [Google Scholar] [CrossRef]
- Banha, J.; Marques, L.; Oliveira, R.; Martins, M.d.F.; Paixão, E.; Pereira, D.; Malhó, R.; Penque, D.; Costa, L. Ceruloplasmin expression by human peripheral blood lymphocytes: A new link between immunity and iron metabolism. Free Radic. Biol. Med. 2008, 44, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Burger, D.; Dayer, J.M. High-density lipoprotein-associated apolipoprotein A-I: The missing link between infection and chronic inflammation? Autoimmun. Rev. 2002, 1, 111–117. [Google Scholar] [CrossRef]
- Souto, S.; Mérour, E.; Biacchesi, S.; Brémont, M.; Olveira, J.G.; Bandín, I. In vitro and in vivo characterization of molecular determinants of virulence in reassortant betanodavirus. J. Gen. Virol. 2015, 96, 1287–1296. [Google Scholar] [CrossRef] [Green Version]
- Harris, M.A.; Clark, J.; Ireland, A.; Lomax, J.; Ashburner, M.; Foulger, R.; Eilbeck, K.; Lewis, S.; Marshall, B.; Mungall, C.; et al. Gene Ontology Consortium. The Gene Ontology (GO) database and informatics resource. Nucleic Acids Res. 2004, 32, D258–D261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaus, J.P.; Eisenhauer, P.; Russo, J.; Mason, A.B.; Do, D.; King, B.; Taatjes, D.; Cornillez-Ty, C.; Boyson, J.E.; Thali, M.; et al. The intracellular cargo receptor ERGIC-53 is required for the production of infectious arenavirus, coronavirus, and filovirus particles. Cell Host Microbe 2013, 14, 522–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapani, J.A.; Smyth, M.J. Functional significance of the perforin/granzyme cell death pathway. Nat. Rev. Immunol. 2002, 2, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Gorissen, M.; Flik, G. The Endocrinology of the Stress Response in Fish. In Biology of Stress in Fish-Fish Physiology; Tort, L., Farrell, A.P., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 35. [Google Scholar] [CrossRef]
- Dai, Z.; Li, J.; Hu, C.; Wang, F.; Wang, B.; Shi, X.; Hou, Q.; Huang, W.; Lin, G. Transcriptome data analysis of grass carp (Ctenopharyngodon idella) infected by reovirus provides insights into two immune-related genes. Fish Shellfish Immunol. 2017, 64, 68–77. [Google Scholar] [CrossRef]
- Liu, P.; Wang, L.; Ye, B.Q.; Huang, S.; Wong, S.M.; Yue, G.H. Characterization of a novel disease resistance gene rtp3 and its association with VNN disease resistance in Asian seabass. Fish Shellfish Immunol. 2017, 61, 61–67. [Google Scholar] [CrossRef]
- Wang, W.; Huang, Y.; Yu, Y.; Yang, Y.; Xu, M.; Chen, X.; Ni, S.; Qin, Q.; Huang, X. Fish TRIM39 regulates cell cycle progression and exerts its antiviral function against iridovirus and nodavirus. Fish Shellfish Immunol. 2016, 50, 1–10. [Google Scholar] [CrossRef]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Jiang, S.; Wu, W.; Yu, F.; Chang, W.; Li, P.; Wang, K. Non-coding RNAs Function as Immune Regulators in Teleost Fish. Front. Immunol. 2018, 9, 2801. [Google Scholar] [CrossRef]
- Pietrangelo, A.; Ridgway, N.D. Bridging the molecular and biological functions of the oxysterol-binding protein family. Cell. Mol. Life Sci. 2018, 75, 3079–3098. [Google Scholar] [CrossRef]
- Cai, J.; Zou, Z.; Wei, S.; Zheng, Q.; Xu, Y.; Lu, Y.; Wu, Z.; Qin, Q.; Jian, J. Identification of Beclin-1 from orange-spotted grouper (Epinephelus coioides) involved in viral infection. Fish Shellfish Immunol. 2019, 94, 336–345. [Google Scholar] [CrossRef]
- Serada, K.; Moritomo, T.; Teshirogi, K.; Itou, T.; Shibashi, T.; Inoue, Y.; Nakanishi, T. Comparison of respiratory burst activity of inflammatory neutrophils in ayu (Plecoglossus altivelis) and carp (Cyprinus carpio). Fish Shellfish Immunol. 2005, 19, 363–373. [Google Scholar] [CrossRef]
- Cha, I.S.; Kwon, J.; Mun, J.Y.; Park, S.B.; Jang, H.B.; Nho, S.W.; del Castillo, C.S.; Hikima, J.; Aoki, T.; Jung, T.S. Cathepsins in the kidney of olive flounder, Paralichthys olivaceus, and their responses to bacterial infection. Dev. Comp. Immunol. 2012, 38, 538–544. [Google Scholar] [CrossRef]
- Gorski, S.A.; Lawrence, M.G.; Hinkelman, A.; Spano, M.M.; Steinke, J.W.; Borish, L.; Teague, W.G.; Braciale, T.J. Expression of IL-5 receptor alpha by murine and human lung neutrophils. PLoS ONE 2019, 14, e0221113. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.W.; Su, Y.C.; Her, G.M.; Ken, C.F.; Hong, J.R. Betanodavirus induces oxidative stress-mediated cell death that prevented by anti-oxidants and zfcatalase in fish cells. PLoS ONE 2011, 6, e25853. [Google Scholar] [CrossRef]
- Star, B.; Nederbragt, A.J.; Jentoft, S.; Grimholt, U.; Malmstrøm, M.; Gregers, T.F.; Rounge, T.B.; Paulsen, J.; Solbakken, M.H.; Sharma, A.; et al. The genome sequence of Atlantic cod reveals a unique immune system. Nature 2011, 477, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Koonin, E.V.; Aravind, L. The NACHT family—A new group of predicted NTPases implicated in apoptosis and MHC transcription activation. Trends Biochem. Sci. 2000, 25, 223–224. [Google Scholar] [CrossRef]
- Martinon, F.; Tschopp, J. NLRs join TLRs as innate sensors of pathogens. Trends Immunol. 2005, 26, 447–454. [Google Scholar] [CrossRef]
- Rhodes, D.A.; de Bono, B.; Trowsdale, J. Relationship between SPRY and B30.2 protein domains. Evolution of a component of immune defence? Immunology 2005, 116, 411–417. [Google Scholar] [CrossRef]
- Davidson, S.; Maini, M.K.; Wack, A. Disease-promoting effects of type I interferons in viral, bacterial, and coinfections. J. Interferon Cytokine Res. 2015, 35, 252–264. [Google Scholar] [CrossRef]
- Chen, Y.M.; Wang, T.Y.; Chen, T.Y. Immunity to betanodavirus infections of marine fish. Dev. Comp. Immunol. 2014, 43, 174–183. [Google Scholar] [CrossRef]
- Yan, X.; Zhao, X.; Huo, R.; Xu, T. IRF3 and IRF8 Regulate NF-κB Signaling by Targeting MyD88 in Teleost Fish. Front. Immunol. 2020, 11, 606. [Google Scholar] [CrossRef]
- Chawla-Sarkar, M.; Lindner, D.J.; Liu, Y.F.; Williams, B.R.; Sen, G.C.; Silverman, R.H.; Borden, E.C. Apoptosis and interferons: Role of interferon-stimulated genes as mediators of apoptosis. Apoptosis 2003, 8, 237–249. [Google Scholar] [CrossRef]
- Ritchie, K.J.; Zhang, D.E. ISG15: The immunological kin of ubiquitin. Semin. Cell Dev. Biol. 2004, 15, 237–246. [Google Scholar] [CrossRef]
- Weiss, J.M.; Nath, A.; Major, E.O.; Berman, J.W. HIV-1 Tat induces monocyte chemoattractant protein-1-mediated monocyte transmigration across a model of the human blood-brain barrier and up-regulates CCR5 expression on human monocytes. J. Immunol. 1999, 163, 2953–2959. [Google Scholar]
- Gerard, C.; Rollins, B.J. Chemokines and disease. Nat. Immunol. 2001, 2, 108–115. [Google Scholar] [CrossRef]
- Kurt-Jones, E.A.; Chan, M.; Zhou, S.; Wang, J.; Reed, G.; Bronson, R.; Arnold, M.M.; Knipe, D.M.; Finberg, R.W. Herpes simplex virus 1 interaction with Toll-like receptor 2 contributes to lethal encephalitis. Proc. Natl. Acad. Sci. USA 2004, 101, 1315–1320. [Google Scholar] [CrossRef] [Green Version]
- Culley, F.J.; Pennycook, A.M.; Tregoning, J.S.; Hussell, T.; Openshaw, P.J. Differential chemokine expression following respiratory virus infection reflects Th1- or Th2-biased immunopathology. J. Virol. 2006, 80, 4521–4527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, Y.T.; Liao, F.; Hsiao, C.H.; Kao, C.L.; Chen, Y.C.; Wu-Hsieh, B.A. Modeling the early events of severe acute respiratory syndrome coronavirus infection in vitro. J. Virol. 2006, 80, 2684–2693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaudreault, E.; Fiola, S.; Olivier, M.; Gosselin, J. Epstein-Barr virus induces MCP-1 secretion by human monocytes via TLR2. J. Virol. 2007, 81, 8016–8024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, M.K.; Kumawat, K.L.; Basu, A. Japanese encephalitis virus differentially modulates the induction of multiple pro-inflammatory mediators in human astrocytoma and astroglioma cell-lines. Cell Biol. Int. 2008, 32, 1506–1513. [Google Scholar] [CrossRef]
- Kanoke, A.; Fujimura, M.; Niizuma, K.; Fujimura, T.; Kakizaki, A.; Ito, A.; Sakata, H.; Sato-Maeda, M.; Kure, S.; Tominaga, T. Temporal profile of magnetic resonance angiography and decreased ratio of regulatory T cells after immunological adjuvant administration to mice lacking RNF213, a susceptibility gene for moyamoya disease. Brain Res. 2016, 1642, 1–9. [Google Scholar] [CrossRef]
- Barton, B.A.; Iwama, G.K. Physiological changes in fish from stress in aquaculture with emphasis on the response and effects of corticosteroids. Ann. Rev. Fish Dis. 1991, 1, 13–26. [Google Scholar] [CrossRef]
- Galińska-Rakoczy, A.; Engel, P.; Xu, C.; Jung, H.; Craig, R.; Tobacman, L.S.; Lehman, W. Structural basis for the regulation of muscle contraction by troponin and tropomyosin. J. Mol. Biol. 2008, 379, 929–935. [Google Scholar] [CrossRef] [Green Version]
- Stoupa, A.; Adam, F.; Kariyawasam, D.; Strassel, C.; Gawade, S.; Szinnai, G.; Kauskot, A.; Lasne, D.; Janke, C.; Natarajan, K.; et al. TUBB1 mutations cause thyroid dysgenesis associated with abnormal platelet physiology. EMBO Mol. Med. 2018, 10, e9569. [Google Scholar] [CrossRef]
- Iseka, F.M.; Goetz, B.T.; Mushtaq, I.; An, W.; Cypher, L.R.; Bielecki, T.A.; Tom, E.C.; Arya, P.; Bhattacharyya, S.; Storck, M.D.; et al. Role of the EHD Family of Endocytic Recycling Regulators for TCR Recycling and T Cell Function. J. Immunol. 2018, 200, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Ding, L.; Zi, Z.; Gao, S.; Wang, C.; Wang, Y.; Zhu, C.; Yuan, Z.; Wei, F.; Cai, Q. Viral-Mediated AURKB Cleavage Promotes Cell Segregation and Tumorigenesis. Cell Rep. 2019, 26, 3657–3671. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, P.; Nawadkar, R.; Ojha, H.; Kumar, J.; Sahu, A. Complement Evasion Strategies of Viruses: An Overview. Front. Microbiol. 2017, 8, 1117. [Google Scholar] [CrossRef] [Green Version]
- Moraes-Vieira, P.M.; Castoldi, A.; Aryal, P.; Wellenstein, K.; Peroni, O.D.; Kahn, B.B. Antigen Presentation and T-Cell Activation Are Critical for RBP4-Induced Insulin Resistance. Diabetes 2016, 65, 1317–1327. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.B.; Qiu, C.; Zhang, A.L.; Cai, L.; Lin, S.J.; Yao, Y.; Tang, Q.S.; Xu, M.; Hua, W.; Chu, Y.W.; et al. Glioma-associated antigen HEATR1 induces functional cytotoxic T lymphocytes in patients with glioma. J. Immunol. Res. 2014, 2014, 131494. [Google Scholar] [CrossRef]
- Matsumiya, T.; Xing, F.; Ebina, M.; Hayakari, R.; Imaizumi, T.; Yoshida, H.; Kikuchi, H.; Topham, M.K.; Satoh, K.; Stafforini, D.M. Novel role for molecular transporter importin 9 in posttranscriptional regulation of IFN-ε expression. J. Immunol. 2013, 191, 1907–1915. [Google Scholar] [CrossRef] [Green Version]
- Hammer, C.; Wanitchakool, P.; Sirianant, L.; Papiol, S.; Monnheimer, M.; Faria, D.; Ousingsawat, J.; Schramek, N.; Schmitt, C.; Margos, G.; et al. A Coding Variant of ANO10, Affecting Volume Regulation of Macrophages, Is Associated with Borrelia Seropositivity. Mol. Med. 2015, 21, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Bayrer, J.R.; Wang, H.; Nattiv, R.; Suzawa, M.; Escusa, H.S.; Fletterick, R.J.; Klein, O.D.; Moore, D.D.; Ingraham, H.A. LRH-1 mitigates intestinal inflammatory disease by maintaining epithelial homeostasis and cell survival. Nat. Commun. 2018, 9, 4055. [Google Scholar] [CrossRef] [PubMed]
- Labernadie, A.; Trepat, X. Sticking, steering, squeezing and shearing: Cell movements driven by heterotypic mechanical forces. Curr. Opin. Cell Biol. 2018, 54, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Mohan, H.; Krumbholz, M.; Sharma, R.; Eisele, S.; Junker, A.; Sixt, M.; Newcombe, J.; Wekerle, H.; Hohlfeld, R.; Lassmann, H.; et al. Extracellular matrix in multiple sclerosis lesions: Fibrillar collagens, biglycan and decorin are upregulated and associated with infiltrating immune cells. Brain Pathol. 2010, 20, 966–975. [Google Scholar] [CrossRef]
- Nishikimi, A.; Koyama, Y.I.; Ishihara, S.; Kobayashi, S.; Tometsuka, C.; Kusubata, M.; Kuwaba, K.; Hayashida, O.; Hattori, S.; Katagiri, K. Collagen-derived peptides modulate CD4+ T-cell differentiation and suppress allergic responses in mice. Immun. Inflamm. Dis. 2018, 6, 245–255. [Google Scholar] [CrossRef] [Green Version]
- Cook, K.C.; Moreno, J.A.; Jean Beltran, P.M.; Cristea, I.M. Peroxisome Plasticity at the Virus-Host Interface. Trends Microbiol. 2019, 27, 906–914. [Google Scholar] [CrossRef]
- Rivas, C.; Aaronson, S.A.; Munoz-Fontela, C. Dual Role of p53 in Innate Antiviral Immunity. Viruses 2010, 2, 298–313. [Google Scholar] [CrossRef] [Green Version]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. PLoS Med. 2021, 18, e1003583. [Google Scholar] [CrossRef]
- Yuan, Z.; Liu, S.; Zhou, T.; Tian, C.; Bao, L.; Dunham, R.; Liu, Z. Comparative genome analysis of 52 fish species suggests differential associations of repetitive elements with their living aquatic environments. BMC Genom. 2018, 19, 141. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Fish Species | Organ Sample/Cell Culture | Sampling Time | Analysis Method | Number of Up-Regulated DEGs | Number of Down-Regulated DEGs | Reference |
|---|---|---|---|---|---|---|
| Psetta maxima | Head kidney | 3 hpi; 6 hpi; 24 hpi; 3 dpi | Microarray | 94 | 12 | [29] |
| Epinephelus spp. | Grouper kidney cells | 6 hpi; 33 hpi | RNA-Seq | N/A | N/A | [30] |
| Epinephelus coiodes | Larvae whole | natural infection | Microarray | 17 | 47 | [31] |
| Gadus morhua | Brain | 5 dpi; 11 dpi; 25 dpi | Microarray | 1179 | 454 | [32] |
| Oryzias latipes | Whole fish | 24 hpi | Microarray | N/A | N/A | [33] |
| Danio rerio | Embryonic-derived zebrafish cells | 6 hpi | Microarray | N/A | N/A | [34] |
| Lates calcarifer | Epithelial cells | 6 hpi; 12 hpi; 24 hpi; 48 hpi | RNA-Seq | N/A | N/A | [35] |
| Channa striatus | Striped snakehead fish cells | 3 hpi; 24 hpi | RNA-Seq | 1184, 3 hpi 1138 24 hpi | 1456, 3 hpi 2073, 24 hpi | [36] |
| Dicentrarchus labrax | Head-kidney leucocytes | 4 hpi | RNA-Seq | N/A | N/A | [37] |
| Hyporthodus septemfasciatus | Brain | 3 dpi; 4 dpi | RNA-Seq | N/A | N/A | [38] |
| Epinephelus coiodes | Grouper fin cell line | 3 hpi; 24 hpi | RNA-Seq | 43 | 8 | [39] |
| Lateolabrax japonicus | Lateolabrax japonicus brain cells | 48 hpi | RNA-Seq | 1969 | 9858 | [40] |
| Solea senegalensis | Head kidney; eye/Brain | 48 hpi | RNA-Seq | N/A | N/A | [41] |
| Epinephelus malabaricus | Brain | 9 dpi | RNA-Seq | N/A | N/A | [42] |
| Dicentrarchus labra | D. labrax brain cells | 12 hpi; 3 dpi | RNA-Seq | 834, 12 dpi 1053, 72 dpi | 810, 12 dpi 1872, 12 dpi | [43] |
| Epinephelus moara | Liver; spleen; head kidney | natural infection | RNA-Seq | 77, liver 89, spleen 318 kidney | 6, liver 161, spleen 5314, kidney | [44] |
| Dicentrarchus labrax | Brain; head kidney | 24 hpi; 3 dpi | RNA-Seq | N/A | N/A | [45] |
| Dicentrarchus labrax | Brain; head kidney | 24 hpi; 3 dpi | RNA-Seq | 196, head kidney 800, brain | 44, head kidney 193, brain | [46] |
| Lates calcarifer | Head kidney | 2 dpi; 15 dpi | RNA-Seq | N/A | N/A | [47] |
| Epinephelus coiodes | Larvae whole | 5 dpi | RNA-Seq | 17,939 | 2526 | [48] |
| Dicentrarchus labrax | Head kidney; Brain | 1, 3, 5, 7 dpi | OpenArray® chip | N/A | N/A | [6] |
| Solea senegalensis | Head kidney; eye/Brain | 2 dpi; 3 dpi | OpenArray® chip | N/A | N/A | [49] |
| Sparus aurata | Larvae whole | 6 hpi; 12 hpi; 24 hpi; 48 hpi | RNA-Seq | 148, 12 dpi | 158, 12 dpi | [50] |
| Lateolabrax japonicus | Lateolabrax japonicus brain cells | 12 hpi; 48 hpi | RNA-Seq | 35, 12 dpi 52, 48 dpi | 54, 12 dpi 50, 48 dpi | [51] |
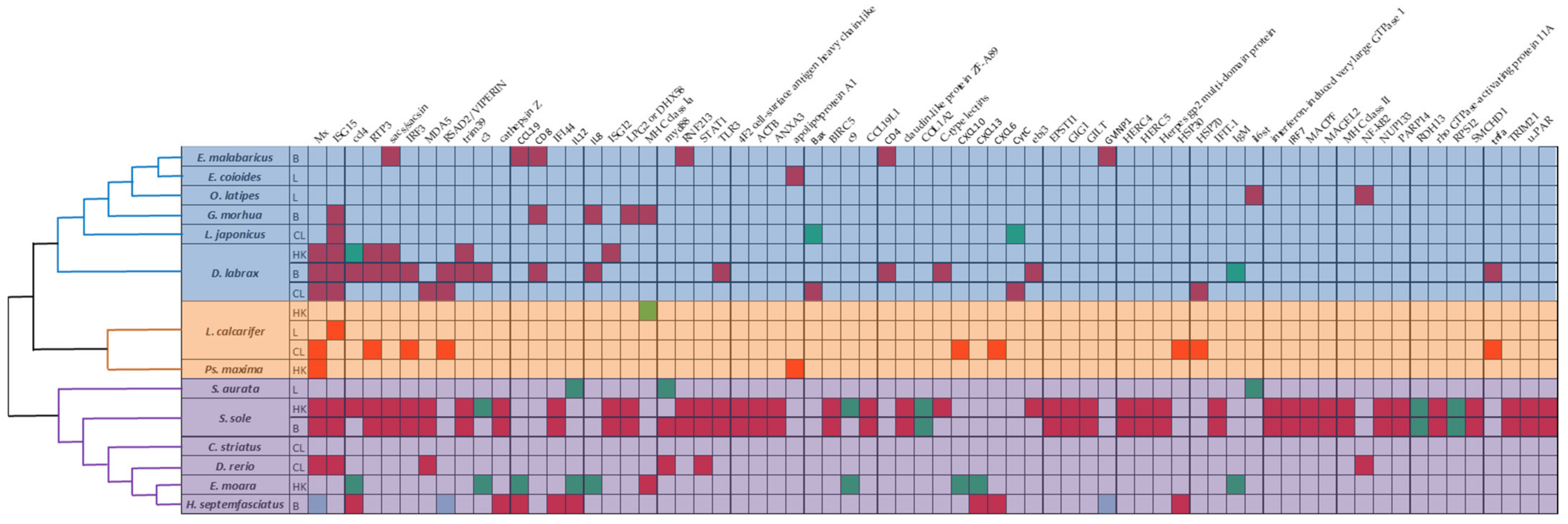
| Fish Species | Up-Regulated DEGs | Down-Regulated DEGs |
|---|---|---|
| Ps. maxima | Mx, IFI35, BiP, serum lectin isoform 4, serum-inducible protein kinase, ceruloplasmin, kininogen I, haptoglobin, thrombin, proteinase activated receptor 3, apolipoprotein A1 | F-box only protein 25, 5-aminolevulinate synthase, phosphatidylinositol 4-kinase |
| S. sole | Mx, LPG2 (DHX58), ISG15, STAT1, HERC5, IFI44, IFIT-1, NUP133, TRIM21, MACPF, u-PAR, PARP14, EPSTI1, ISG12, RNF213, HERC4, MHC class II, ANXA3, RTP3, CCL19L1, SMCHD1, ACTB, GIG1, interferon-induced very large, GTPase 1, IRF3, IRF7, TLR3, MAGEL2, GILT, Herpes gp2 multi-domain protein, claudin-like protein ZF-A89, 4F2 cell-surface antigen heavy chain-like, cathepsin Z, BIRC5, rho GTPase-activating protein 11A, v-ets erythroblastosis virus E26 oncogene homolog 1, C-type lectins, MDA5, LITAF, SACS/SACSIN, TRIM39, CCL4, CCK, EBI3 | COL1A2, RDH13, RPS12, LMAN1/ERGIC-53, C3, C9, CFHR3 |
| E. moara | MHC class I, HLA-A, MR1, lysozyme, hepcidin | C3, C9, IgM, IgD, IgT/IgZ, perforin, IL1, IL2, IL8, IL11, IL12, IL15, C1, C7, C8, CCL4, CXCL13, CXCL10, CCL3, CCL19, CXCL12, CCL20, CCL25, CCR9, CXCR4, CXCR3, CXR1 |
| D. labrax | Mx, ISG15, ISG12, RTP3, SACS/SACSIN, TRIM39, CYP11b1, STAR, CYP21a, CYP17a1, 3BHSD, CYP11a1, IFI56 | CCL4, ERO1a, HYOU1, PDIA4, QORX, CXC114, VLIG |
| L. calcarifer | beclin-1, IP3R, CYBA, C3aR, NCF2, OSBP1, cathepsin C/CTSC, IL5Rα/CD125 | CD53, MHC class Ia, CH25H, BTK, STAT5b, SOCS1 |
| Fish Species | Up-Regulated DEGs | Down-Regulated DEGs |
|---|---|---|
| G. morhua | LPG2 (DHX58), ISG15, IL8, MHC class Ia, NILT, NITR, CD3, CD8, TCR, RHOG1, ITK, FYB, LCK, granzyme, CC chemokines type-3, IL17a, MHC I complex-B2M, tapasins | |
| S. sole | Mx, LPG2 or DHX58, ISG15, STAT1, HERC5, IFI44, IFIT-1, NUP133, TRIM21, MACPF, u-PAR, PARP14, EPSTI1, ISG12, RNF213, HERC4, MHC class II, ANXA3, RTP3, CCL19L1, SMCHD1, ACTB, GIG1, interferon-induced very large GTPase 1, IRF3, IRF7, TLR3, MAGEL2, GILT, Herpes gp2 multi-domain protein, claudin-like protein ZF-A89, 4F2 cell-surface antigen heavy chain-like, cathepsin Z, BIRC5, rho GTPase-activating protein 11A, v-ets erythroblastosis virus E26 oncogene homolog 1, MDA5, SACS/SACSIN, TRIM39, MyD88, TBKBP1/SINTAD, PKR, HERC5 | COL1A2, RDH13, RPS12 |
| H. septemfasciatus | Mx, IFI44, CCL4, CXCL13, CCL19, CCL2, CCL34, CXCL6, CXCL8, CXCL9, IL12A, IL12B, IL18B, cathepsin L, cathepsin H, cathepsin K, cathepsin O, cathepsin S, cathepsin Z, CLEC4M, CLEC10A, CD209, LGALS9, LGALS3, FUCL4, MBL, RSAD2/VIPERIN, NK-Lysin, IFIT5, GVINP1, EIF2Ak2, IFIH1, HSP30 | |
| E. malabaricus | SACS/SACSIN, CCL19, GVINP1, PD-L1, LAG3, CD3E, CD4, CD8A, RNF213, TREX, ZNFX1 | NPTN, SF3B1, EWSR1, NLRX1, GNG3, MGEA5 |
| D. labrax | Mx, ISG15, RTP3, IRF3, TLR3, C-type lectins, SACS/SACSIN, TRIM39, C3, CCL4, EBI3, IL8, RSAD2/VIPERIN, CD4, LECT2, NKTR, PTX4, C1QT1, PRL, SL, SOMA, GH, GLHA, TSH, POMC, 5HT3E, GABA, cathepsin, TLR8, TLR21, TNFa, IL1b, NCCRP-1, TCR-γ, CD8 | IgM, STING, NLRC3 |
| Fish Species | Up-Regulated DEGs | Down-Regulated DEGs |
|---|---|---|
| E. coioides | apolipoprotein A1, MyBP-H, MLC2, white muscle troponin T, ADK1-2, MLC3, tropomyosin, parvalbumin PV1, parvalbumin PV2, trypsinogen, tubulin beta-1 chain, serine/threonine-protein phosphatase 5/AURKB, PAST-1, integrin alpha-V isoform X1, integrin beta-4, ANO10, chymotrypsinogen, claudin-15-like, afadin isoform X8, cadherin-17, collagen type I alpha 3 chains, collagen alpha-1(XVIII) chain isoform X2, collagen alpha-2(V) chain, 2-oxoglutarate 5-dioxygenase 1 isoform X2 | C5, retinol-binding protein 4-A like, HEAT repeat-containing protein 1, importin-9 |
| S. aurata | HSPA5, HSPA9, HSC70, RAB10, RAB11a, AP2a1, CHMP2a, TRAP1, UNC45b, FKBP4, PTGES3a, PEX5la, ATG4, ATG16, BNIP3, DPCP1 | MyD88, IL12b, DNAJ4, IRF5, PIK3R1, STAT3, JAK1, IL6st, PEX3, PEX26, PEX11, EPHX2, ACOT8, GSTK1, LC3, ATG9L, STX17 |
| O. latipes | PVALB, CEBPA, IFIM, IFN, IL-6ST, NF-kB2, SOC3, SP1, TGFB1 |
| Fish Species | Up-Regulated DEGs | Down-Regulated DEGs |
|---|---|---|
| D. labrax | Mx, ISG15, MDA5, RSAD2/VIPERIN, HSP70, HSP90, Fas, TRAF2, AIP, Bid, Bax, CASP3, CASP6, CASP7, CASP9, CASP10, CytC, DIABLO, Apaf-1 | GLUD1, CDD, HMOX1, MAP6 |
| L. japonicus | ISG15, MDM2, MDM4 | Bax, CytC, CASP8 |
| L. calcarifer | Mx, RTP3, IRF3, CXCL10, CXCL6, RSAD2/VIPERIN, HSP30, TNFa, HSP70, CC chemokine, IL17Fl, TNFAIP3, IFN-a1, GVIN1, IFI27L2, VSG | |
| D. rerio | ISG15, STAT1, MDA5, MyD88, NF-kB2, LGP2, TRAF3, TRAF6, IRAK1, IFNPHI1, IFNPHI3, MxA, MxC, PKZ, STAT2 | |
| E. spp. | BiP, PERK, ATF6, IRE1, XBP-1, CHOP, GADD34 | |
| C. striatus | Endonuclease G |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toubanaki, D.K.; Efstathiou, A.; Karagouni, E. Transcriptomic Analysis of Fish Hosts Responses to Nervous Necrosis Virus. Pathogens 2022, 11, 201. https://doi.org/10.3390/pathogens11020201
Toubanaki DK, Efstathiou A, Karagouni E. Transcriptomic Analysis of Fish Hosts Responses to Nervous Necrosis Virus. Pathogens. 2022; 11(2):201. https://doi.org/10.3390/pathogens11020201
Chicago/Turabian StyleToubanaki, Dimitra K., Antonia Efstathiou, and Evdokia Karagouni. 2022. "Transcriptomic Analysis of Fish Hosts Responses to Nervous Necrosis Virus" Pathogens 11, no. 2: 201. https://doi.org/10.3390/pathogens11020201
APA StyleToubanaki, D. K., Efstathiou, A., & Karagouni, E. (2022). Transcriptomic Analysis of Fish Hosts Responses to Nervous Necrosis Virus. Pathogens, 11(2), 201. https://doi.org/10.3390/pathogens11020201