1. Introduction
Bartonella spp. Comprise a genus of Gram-negative alphaproteobacteria that are slow growing, fastidious, and facultative intracellular pathogens with zoonotic potential [
1,
2,
3,
4].
Bartonella spp. are transmitted by arthropod vectors, including fleas, lice, sand flies, and ticks, by animal bites and scratches, or via direct contact with
Bartonella-infected clinical specimens [
4,
5,
6]
. Bartonella spp. are associated with a broad spectrum of clinical signs and pathological abnormalities in dogs [
1,
2,
3,
4]. Dogs infected with
Bartonella spp. develop clinical manifestations that are similar or identical to diseases observed in human patients infected with the same
Bartonella spp., including bacteremia, encephalitis, endocarditis, fever of an unknown origin, lymphadenomegaly, myocarditis, ocular disease (uveitis), peliosis hepatis, and vasculitis [
1,
5,
6,
7,
8,
9,
10]. More than 10
Bartonella spp. have been reported to infect dogs, with
B. henselae,
B. koehlerae, and
B. vinsonii subsp.
berkhoffii being the species or subspecies most frequently identified in sick dogs in North America [
1,
4]. Despite the recent advances in biomedical sciences, clinical diagnostic approaches to confirm
Bartonella infection in dogs have not yet been critically investigated.
Currently, the diagnosis of canine and human Bartonelloses is performed by the isolation of
Bartonella by culture, the amplification of
Bartonella DNA by PCR, and the detection of
Bartonella antibodies by serological assays. Although serology can only confirm exposure, immunofluorescent antibody assays (IFAs), Western Blot (WB), and enzyme-linked immunosorbent assays (ELISAs) are most frequently used for the diagnosis of canine and human Bartonelloses [
11,
12,
13,
14,
15,
16,
17,
18,
19,
20]. Previously, we reported that the sensitivity of IFA did not substantially improve despite using a panel consisting of eight
Bartonella IFA antigens, each tested as an independent serological assay [
21]. In that study,
Bartonella antibodies were not detected in 38% of the
Bartonella spp. bacteremic dogs, which indicated that IFA substantially underestimates the true serological prevalence of
Bartonella spp. infections in dogs. In concordance with our initial findings of poor IFA sensitivity, using a panel of 3
Bartonella IFA antigens, we subsequently confirmed even lower IFA sensitivity (6%) in dogs with hemangiosarcoma, despite the amplification of
Bartonella spp. DNA from 73% (80/110) of the dogs [
22]. Similarly, despite confirming
Bartonella spp. infection in most (71%) PCR-positive dogs by qPCR and ddPCR amplification,
B. henselae WB also had a low sensitivity (33%) in dogs with hemangiosarcoma [
23]. Due to the low sensitivity of currently available serological assays for the diagnosis of canine Bartonelloses, the development of a reliable serological assay is of clinical diagnostic importance and of substantial epidemiological importance for the analysis of prevalence and risk factor studies.
Bartonella henselae Pap31, an outer membrane protein, appears to be an important virulence factor for bacterial attachment and the colonization of mammalian cells, thus promoting the establishment of
B. henselae infection in the host [
24,
25,
26].
Bartonella henselae Pap31, homologous to the hemin binding protein family of
B. quintana, is also involved in heme acquisition [
24,
25].
Bartonella henselae Pap31 shares 58.4% identity with the heme binding protein A (HbpA) of
B. quintana and 31.7% identity with OMP31 porin protein of
Brucella melitensis. Pap31 proteins act as adhesins for fibronectin, heparin, and human umbilical endothelial cells (HUVECs), thereby mediating host–pathogen interactions [
24]. The Pap31 protein of
B. bacilliformis, the etiological agent of Carrion’s disease, has been previously evaluated as a candidate antigen for the development of a reliable serological assay for human Bartonelloses [
27]. Another study, characterizing the immunoproteomic profiles of sera collected from cat scratch disease (CSD) and
B. henselae infective endocarditis patients also identified Pap31 as an immunoreactive candidate protein for the serodiagnosis of Bartonelloses [
28].
To our knowledge, B. henselae recombinant Pap31 and selected fragments have not been investigated for the serodiagnosis of canine and human Bartonelloses. Therefore, we evaluated the diagnostic utility of B. henselae full-length recombinant Pap31 (rPap31) and recombinant Pap31 fragments (N-terminal, middle, and C-terminal domains of recombinant Pap31) by ELISA. Sera from dogs and humans exposed to or infected with Bartonella spp. and control groups (presumptively unexposed; Bartonella PCR negative and IFA negative) were used to screen recombinant Pap31 proteins and peptides. We hypothesized that B. henselae Pap31 protein elicits a sensitive and specific humoral immune response in dogs and humans exposed to Bartonella spp. The aims of this study are: (1) to evaluate the sensitivity and specificity of the entire B. henselae rPap31 protein; (2) to compare the sensitivity and specificity of the recombinant Pap31 N-terminal domain (rPap31-NTD), middle domain (rPap31-MD), and C-terminal domain (rPap31-CTD); and (3) to evaluate the diagnostic utility of B. henselae Pap31 linear B-cell epitopes for the serodiagnoses of canine and human Bartonelloses.
3. Discussion
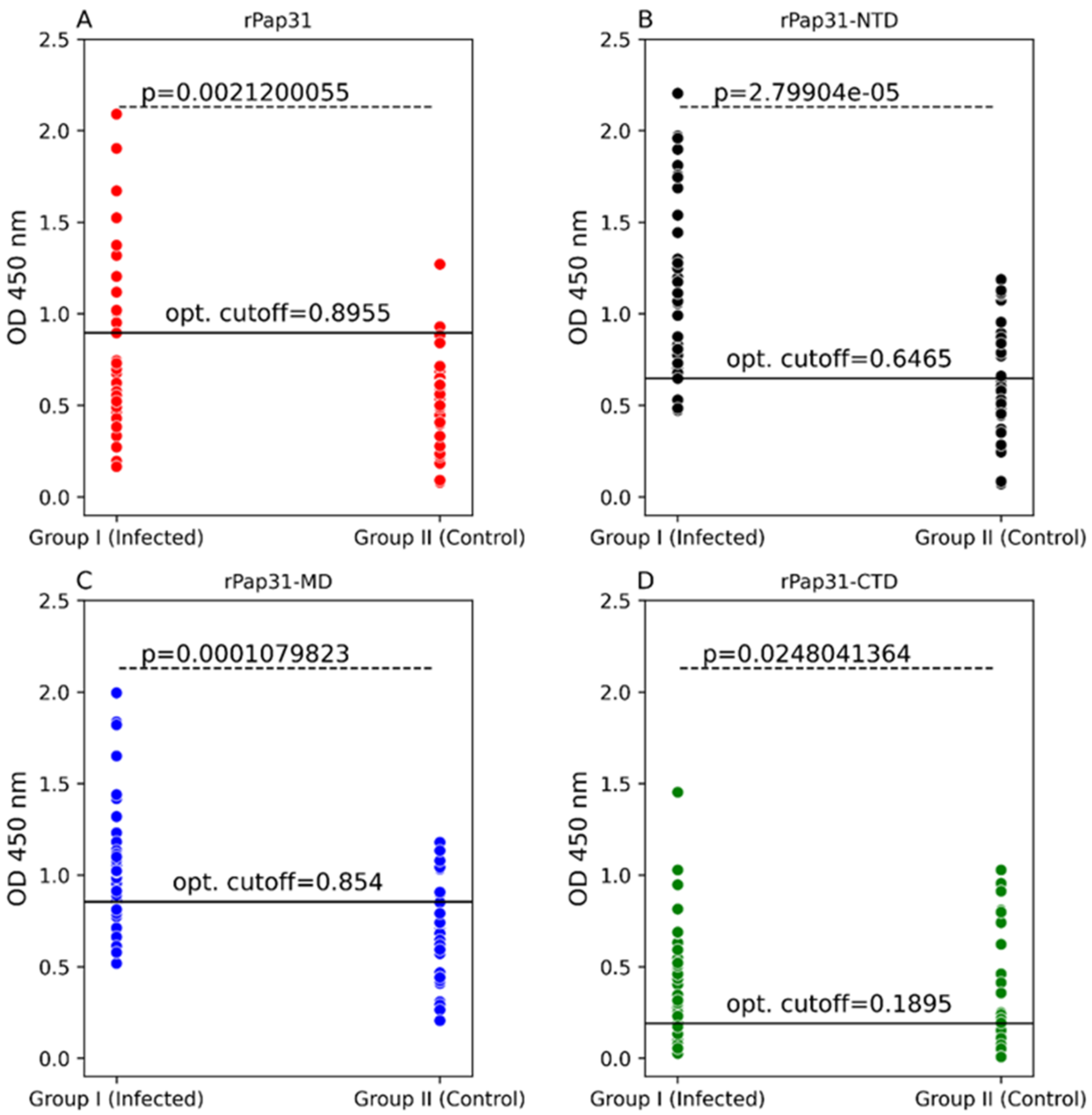
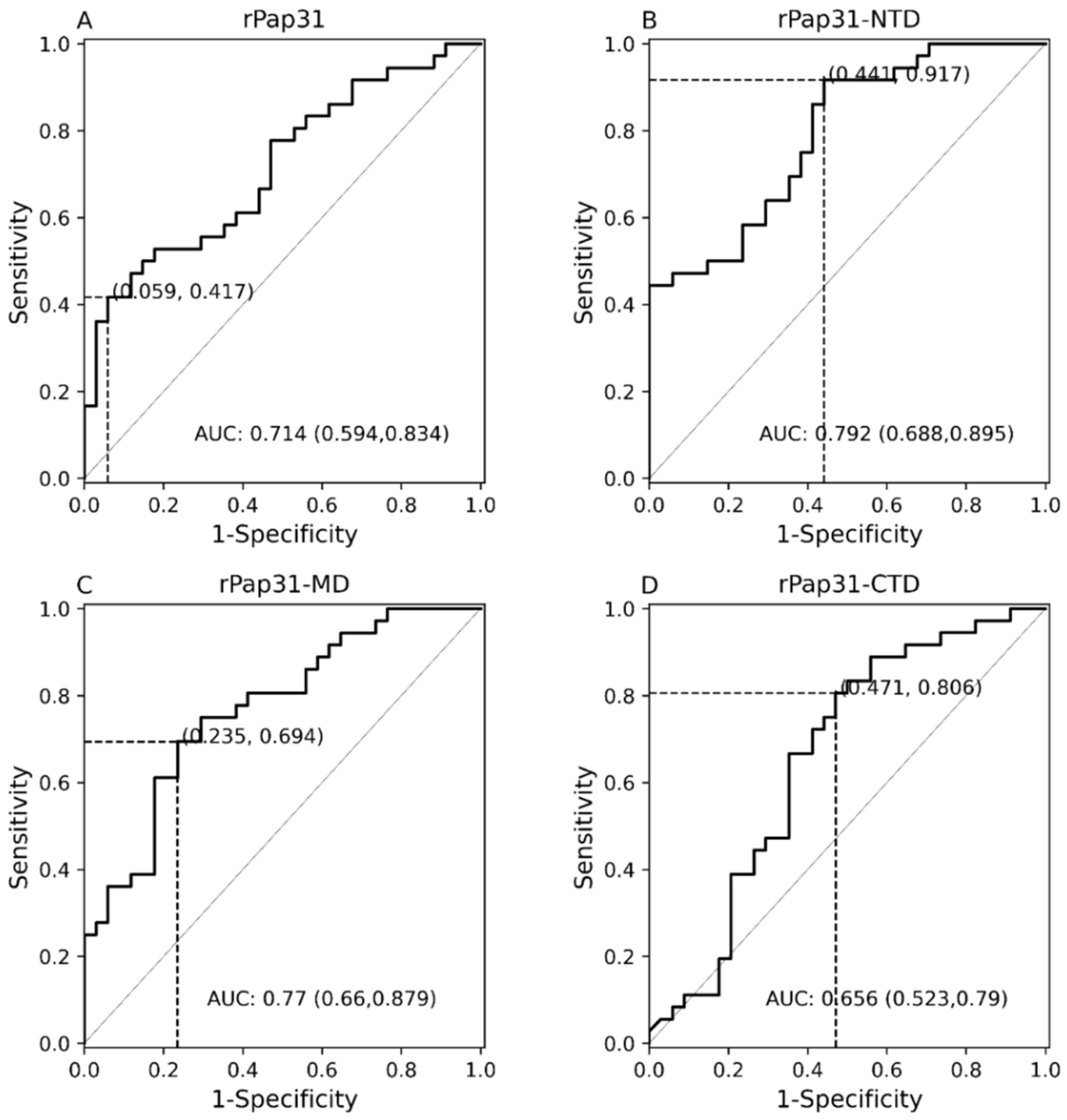
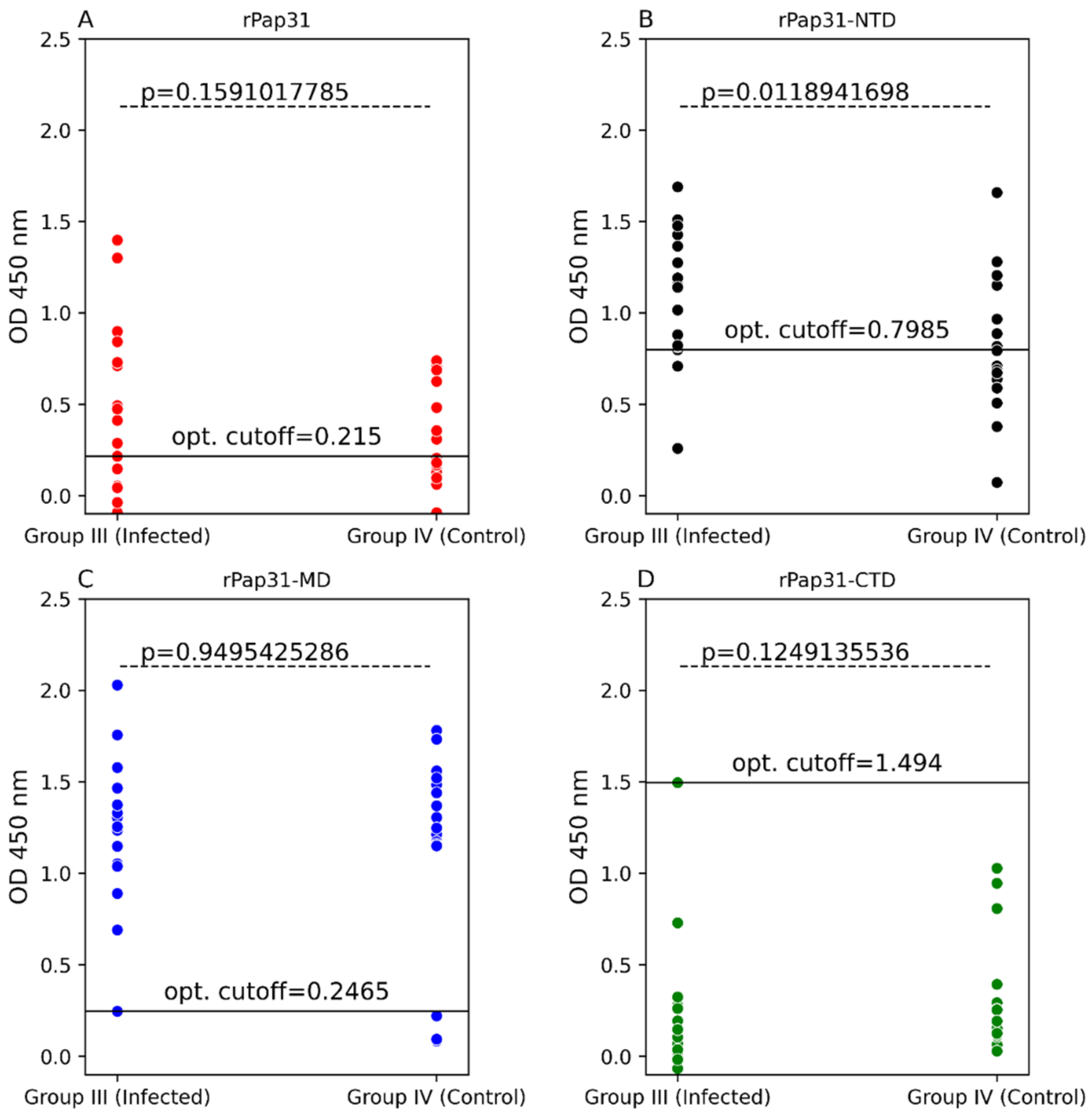
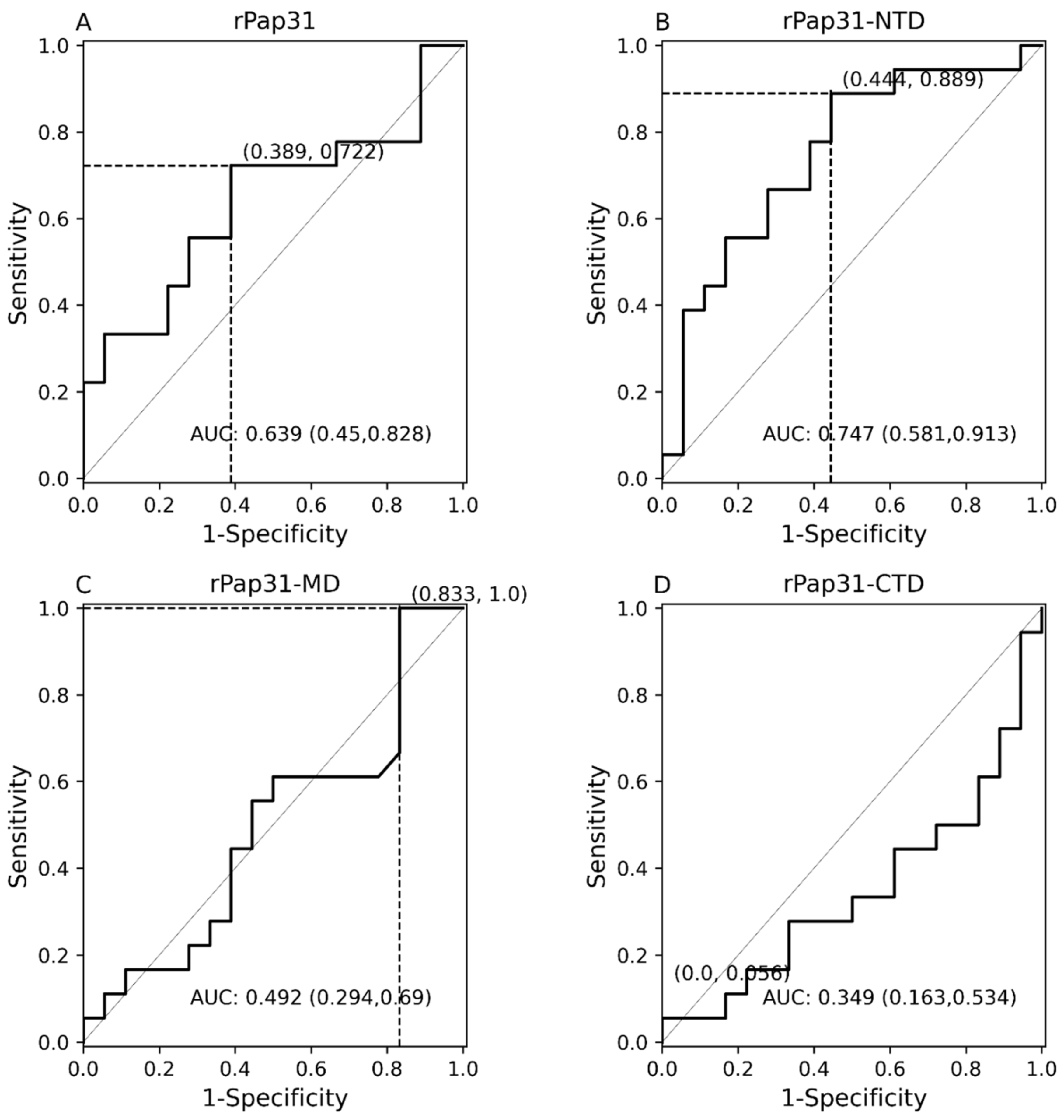
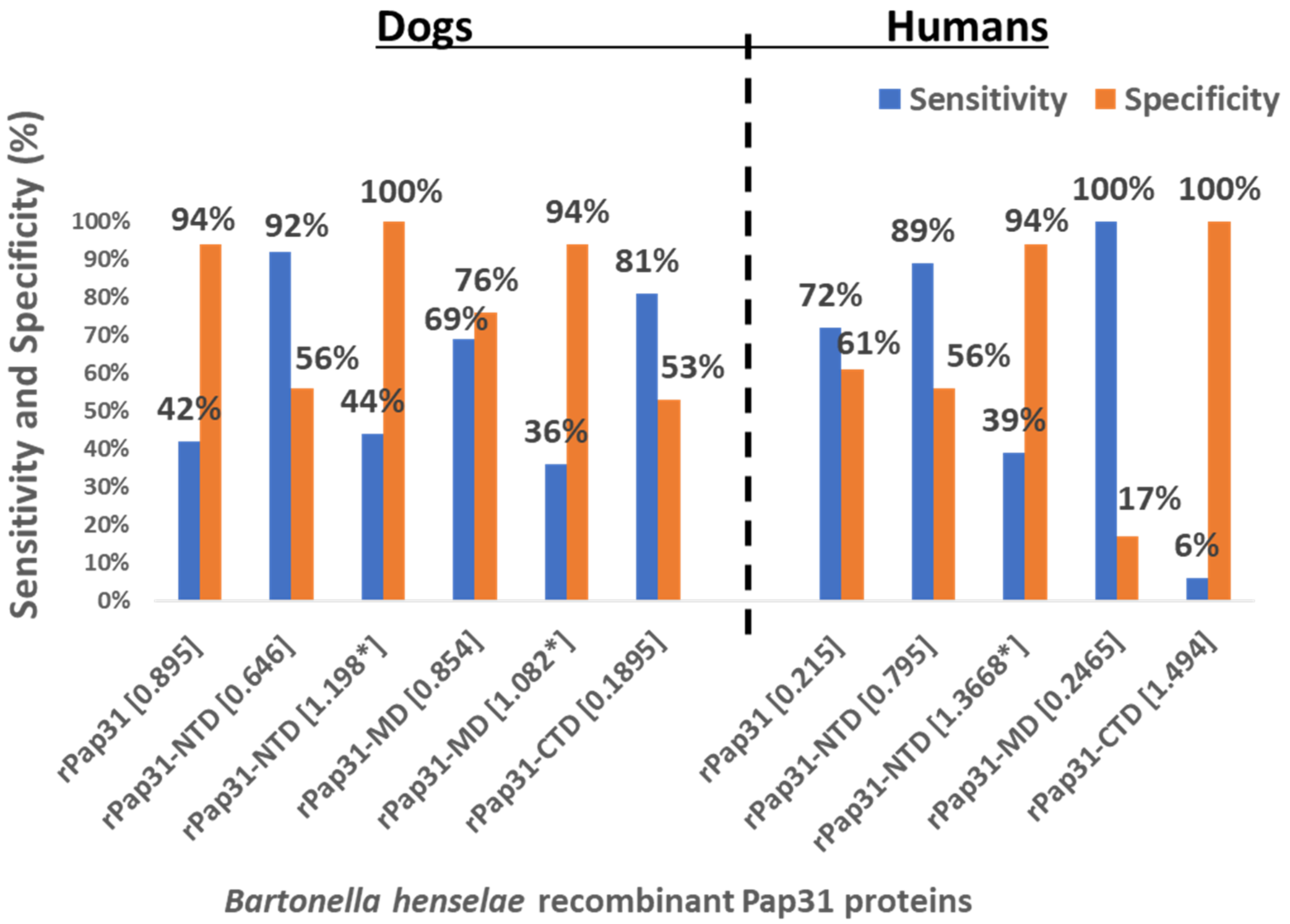
In this study, we evaluated the diagnostic utility of rPap31 and three Pap31 fragments (rPap31-NTD, rPap31-MD, and rPap31-CTD) by ELISA for canine and human Bartonelloses. When testing sera from the dogs and humans that were IFA-positive and/or PCR-positive, the rPap31-NTD and rPap31-MD were the most strongly reactive among the selected recombinant proteins. In dogs, the AUC score of recombinant whole Pap31 (rPap31) was 0.714, with a sensitivity of 42% and specificity of 94% at the OD cutoff value of 0.8955 with a maximum Youden index value, suggesting the diagnostic utility of rPap31 for the diagnosis of Bartonella infection in dogs is questionable due to low sensitivity. Among the rPap31 protein fragments, rPap31-NTD yielded the highest AUC score of 0.792 (95% CI 0.688–0.895) with a sensitivity of 44% and specificity of 100% at a cutoff value of 1.198 when testing dog sera, indicating the relatively poor diagnostic sensitivity of rPap31 and the selected fragments for the diagnosis of canine Bartonelloses. In concordance with the findings from dogs, rPap31-NTD also had the highest AUC score of 0.747 (95% CI 0.581–0.913) among the rPap31 protein fragments for the diagnosis of human Bartonelloses, with 39% sensitivity and 94% specificity at a cutoff value of 1.366. Of the rPap31 protein fragments, the ELISA seroreactivity of rPap31-NTD was significantly different when comparing human Group III (naturally infected) and Group IV (control) individuals. However, there was no difference in the reactivity for the two other rPap31 protein fragments when human IFA-reactive sera were compared to control sera. Due to low sensitivity and questionable specificity, our findings indicate that recombinant Pap31 and none of the selected Pap31 fragments are appropriate as diagnostic targets for detecting anti-Bartonella antibodies in Bartonella-infected dogs or humans.
Nonetheless, rPap31 appears to be highly specific (94%) for the diagnosis of canine Bartonelloses, the specificity of rPap31 was only 61% for the diagnosis of human Bartonelloses. In contrast to the significant differentiation of the infected and control dog groups, there was no significant difference in rPap31 seroreactivity between the infected and control humans. For humans, rPap31 ELISA resulted in 72% sensitivity and 61% specificity, at a cutoff value of 0.215, indicating the potential cross-reactivity of rPap31 with non-
Bartonella antibodies present in the control group sera. Alternatively, some IFA seronegative humans (control group) may have been exposed to a
Bartonella spp., but immunofluorescence was not visualized using cell cultured
Bartonella spp. antigens, as was previously reported (16). In the context of specificity,
B. henselae Pap31 shares homology with
Neisseria opacity proteins (Opa),
Brucella OMP31 (a putative porin), and
Agrobacterium tumefaciens OMP25 (an immunogenic surface protein) [
29,
30]. With the exception of rPap31 for the diagnosis of canine Bartonelloses, the rPap31 protein fragments were relatively non-specific at cutoff OD values with a maximum Youden index for the diagnosis of canine Bartonelloses. For human Bartonelloses, rPap31, rPap31-NTD, and rPap31-MD had a specificity of less than 62% at the maximum Youden index value, while the specificity of rPap31-CTD was 100% at the maximum Youden index value. The high degree of identity shared between the
B. henselae Pap31 protein with proteins of other microorganisms most likely contributed to the low specificity found in this study. These findings indicate that recombinant Pap31 proteins may generate false positive results due to the cross-reactivity with antibodies induced against antigenically similar proteins found in other microorganisms.
A number of factors, such as the selection of cutoff values, antigen preparation, sample population, and standard methods used for the calculation of cutoff values, may influence the assessment of the diagnostic accuracy of an ELISA [
31]. The diagnostic accuracy of serological tests has a considerable impact on animal and human health, as well as important economic and epidemiologic implications. In the context of establishing a cutoff value, the sensitivity and specificity of recombinant Pap31 proteins varied at different ELISA cutoff values in this study. At a cutoff value of 0.6465 (with a maximum Youden index), the rPap31-NTD ROC curve estimated a sensitivity of 92% with a specificity of 56% for the diagnosis of canine Bartonelloses. A higher cutoff value of 1.198 (trade-off between sensitivity and specificity) resulted in 44% sensitivity and 100% specificity for rPap31-NTD. For the diagnosis of human Bartonelloses, the sensitivity and specificity of rPap31-NTD were 39% and 94%, respectively, at a higher cutoff value of 1.366 (trade-off between sensitivity and specificity), while rPap31-NTD resulted in 88% sensitivity and a specificity of 56%, respectively, at a cutoff value of 0.7985 with a maximum Youden index. These findings suggest the ELISA cut-off values must be selected with the utmost care, since the selection of a cut-off value becomes the basis for the calculation of sensitivity and specificity, which determines the accuracy of the test result that will be used diagnostically for patient management and disease control strategies.
Logically,
Bartonella spp. proteins that are associated with survival, multiplication, or bacterial adaptation in accidental and reservoir hosts should be useful targets for the diagnosis of Bartonelloses. Additionally, certain proteins that are important for the pathogen’s survival can be useful for vaccine development but not for serological diagnosis, because of their low antigenicity and high immunogenicity. As a hallmark of these intra-erythrocytic bacteria, the penetration of erythrocytes is an invasion strategy used to obtain heme, which is essential for the growth of
Bartonella spp. in vivo and in vitro [
24,
25,
26,
30,
32]. Heme binding proteins (Hbps) of several
Bartonella spp., including the heme binding protein A (HbpA) of
B. quintana and Pap31 of
B. henselae, play an active role in hemin acquisition, survival, and disease pathogenesis [
25,
26,
30]. Therefore, previous studies suggested that heme binding proteins, including Pap31 proteins, are potential candidates for the development of diagnostic and vaccine candidates for Bartonelloses [
27,
33,
34]. Previously, while assessing
B. bacilliformis recombinant Pap31 for the diagnosis of Oroya fever, the authors demonstrated that Pap31 antigens were highly induced in growing cultures of
B. bacilliformis and were immunologically recognized dominant proteins in infected humans, supporting the application of Pap31 for the ELISA and WB diagnosis of human Bartonelloses [
27]. Dichter et al. (2021), using a reverse vaccinology approach in conjunction with an immunoproteomic approach, also highlighted the potential utility of Pap31 as an immunodominant target recognized by the serum samples obtained from Peruvian patients infected with
B. bacilliformis [
35]. In another study,
B. bacilliformis recombinant pap31 was not reactive with sera from patients with
Coxiella burnetti, Brucella spp., or
B. henselae infection
, supporting a lack of recombinant
B. bacilliformis Pap31 cross-reactivity with these microorganisms [
34]. In contrast to
B. bacilliformis findings,
B. henselae recombinant Pap31 proteins were reactive with sera from a number of dog and human controls in this study. Our findings indicate that the optimization of a recombinant Pap31 protein-based serological assay, perhaps by combining rPap31 fragments in a chimeritope may be needed to enhance sensitivity, specificity, and diagnostic accuracy for the diagnosis of human Bartonelloses.
Despite the predicted high antigenicity scores of the four selected antigenic Pap31 B-cell epitopes (P1, P2, P3, and P4), none of these epitope peptides were reactive with sera from
B. henselae IFA-positive dogs or humans. Post-inoculation sera from dogs that were experimentally infected with either
B. henselae San Antonio 2 or
B. henselae CSU 1 strains, and dogs experimentally infected with
B. vinsonii subsp. berkhoffii genotype III, were also not reactive to these synthetic peptides, suggesting that three-dimensional epitope conformation is likely of critical importance for the documentation of Pap31 seroreactivity. These findings are consistent with the poor seroreactivity results of
B. bacilliformis Pap31 linear epitopes reported in a previous study [
35], where only 2 of the 26 Peruvian patients with
B. bacilliformis infection were reactive to
B. bacilliformis Pap31 linear epitopes in line blots. It is also possible that
B. henselae Pap31 peptide epitopes represent a minimally immunogenic region of the
B. henselae Pap31 protein, and hence were not reactive with sera from
B. henselae IFA-positive dogs and humans in this current study.
The selection of true negative control (
Bartonella spp. PCR- and IFA-negative) samples are critical for assessing the specificity of serological assays for the diagnosis of Bartonelloses. A limitation of this study is the potential inclusion of serum samples from sick dogs and humans into the respective
Bartonella spp. PCR- and IFA-negative control groups, which would negatively impact our assessment of ELISA specificity for the diagnosis of canine and human Bartonelloses. Dog sera were submitted to the NCSU-CVM-VBDDL by veterinarians to test for evidence of exposure to or infection with canine vector-borne disease (CVBD) organisms; therefore, despite negative IFA (with a documented poor sensitivity) and PCR results (similarly poor sensitivity in association with chronic infection), these dogs could have been previously exposed to 1 or more of the 40 plus
Bartonella species [
4,
21,
23]. As an example, despite being obtained from specific pathogen-free dogs maintained in a vector free facility, the dogs infected with
Rickettsia rickettsii appeared to reactivate occult and previously undetected
Bartonella spp. infections [
36]. Thus, despite negative IFA and PCR testing, our experimental dog control sera may have originated from dogs that had experienced prior
Bartonella spp. environmental exposures [
36,
37] and Breitschwerdt EB et al. (unpublished data). Similarly, it is possible that our human controls were naturally exposed to
Bartonella infection and misdiagnosed as unexposed to or uninfected by
Bartonella PCR, BAPGM, and IFA testing due to the less than perfect sensitivity of each of these diagnostic assays [
19,
38,
39]. Another limitation of this study is that the blood samples from the 13 Group II control dogs were not processed in BAPGM enrichment blood culture, which is often necessary to confirm
Bartonella infection in healthy and sick dogs [
5,
36,
40].
Another limitation of this study is the lack of historical and clinical information for the diagnostic specimens submitted for NCU-CVM-VBDDL testing. Although all Group III human patients were tested because of a self-reported history of chronic illness, these sera represented a very heterogenous sample set in the context of symptomatology and duration of illness. This information would be of importance to compare Pap31 seroreactivity versus disease status and duration. Additionally, only Bartonella testing results were available (evidence of exposure to phylogenetically-related pathogens, such as Brucella, were not available) for human sera. Therefore, the potential cross-reactivity of recombinant Pap31 proteins with sera from humans infected with phylogenetically related pathogens was not addressed in our study. In addition, the information on infecting Bartonella spp., genotype, or strain was lacking for 33 Group I (IFA-positive) dogs and 9 Group III (IFA-positive) humans. NCBI Blast searches indicate that B. henselae Pap31 shares 50% to 100% identity with Pap31 sequences of other Bartonella spp., including medically important animal and human pathogens: B. henselae Pap31 (92 to 100%), B. koehlerae Pap31 (83%), B. quintana Pap31 (55%), and 50% with B. bacilliformis Pap31 (50%). Based upon IFA testing, the sera used in this study may or may not have detected antibodies to all known Bartonella spp., genotypes, or strains, which further complicates the evaluation of Pap31-based ELISA sensitivity and specificity.
In conclusion, with additional assessment and optimization, the recombinant Pap31 and rPap31-NTD fragment may be appropriate for canine Bartonelloses diagnostic applications, whereas rPap31-NTD may have application for the diagnosis of human Bartonelloses. The findings from this study can be used to further assess the antigenicity and immunogenicity of B. henselae Pap31 as a diagnostic target. Although our results are potentially promising, further studies are needed to optimize a B. henselae Pap31-based ELISA for the diagnosis of Bartonella infection in dogs and humans.
4. Materials and Methods
4.1. Serum Samples for ELISA Testing
The
Bartonella PCR,
Bartonella droplet digital PCR (ddPCR), and
Bartonella IFA testing results for the dog and human clinical samples that were used for comparative ELISA in this study are summarized in
Table 2 and
Supplementary Table S1. IgG titers of ≥1:64 were considered positive for
Bartonella exposure. In this study, the serum samples from 70 dogs (Group I: 36
Bartonella spp. IFA-positive (naturally infected) and Group II: 34
Bartonella spp. IFA- and PCR-negative (control dogs)) and 36 humans (Group III: 18
Bartonella spp. IFA-positive (naturally infected) and Group IV: 18
Bartonella spp. IFA- and PCR-negative (control humans)) were tested by rPap31, rPap31-NTD, rPap31-MD, and rPap31-CTD-based ELISA. All 70 dogs’ sera were submitted to NCSU-VBDDL for diagnostic testing between 2016 and 2019. All 36 humans’ sera were obtained from the repository maintained by the NCSU-CVM-IPRL (NCSU IRB approval# 1960) between 2009 to 2020.
4.1.1. Dog Serum Samples
Seventy archived sera from dogs previously tested at the North Carolina State University, College of Veterinary Medicine, Vector Borne Diseases Diagnostic Laboratory (NCSU-CVM-VBDDL) or the Intracellular Pathogens Research Laboratory, NCSU-CVM (NCSU-CVM-IPRL) were selected for comparative ELISA testing utilizing each of the purified recombinant Pap31 proteins,
Table 2 and
Supplementary Table S1. The serum samples were categorized into two groups to assess the sensitivity and specificity. All the sera were stored frozen after being submitted to the NCSU-CVM-VBDDL for diagnostic testing between 2016 and 2020. After the initial processing by the NCSU-CVM-VBDDL, sera were stored at −80 °C. Group I consisted of 36 stored frozen serum samples from
Bartonella spp. naturally infected dogs (
Bartonella IFA-positive). A cutoff titer of ≥1:64 was used to define an IFA-positive titer. Of the 36 dogs, 1 dog had a
B. henselae IFA IgG titer of 1:64, whereas the remaining 35 dogs had
B. henselae IFA IgG titers of ≥1:128. Of these 36 dogs, 23 were concurrently
B. vinsonii subsp.
berkhoffii genotype I seropositive (IFA titer range ≥ 1:64 to 1:4096) and 32 dogs were
B. koehlerae seropositive (IFA titer range ≥ 1:64 to 1:4096). A total of 21 of the 36 dogs were IFA-positive (titers ≥ 1:64) to all 3
Bartonella spp. (
B. henselae San Antonio type
2, B. vinsonii subsp
. berkhoffii genotype I
, and
B. koehlerae). Of the 36 Group I dogs,
Bartonella DNA was amplified from the blood of 3 dogs: 2 dogs were PCR-positive for
B. vinsonii and 1 dog was PCR-positive for
B. henselae, Table 2.
Based on NCSU-CVM-VBDDL serological and PCR testing, 24 of the 36 Group I dogs were seronegative for evidence of exposure and PCR-negative for evidence of infection with other canine vector-borne disease (CVBD) organisms that are routinely tested for in the NCSU-CVM-VBDDL. Detailed methods for the
Bartonella PCR, BAPGM enrichment blood culture, and the IFA serological panel used to test these study participants were previously published [
14,
21]. Specifically, all sera were IFA-negative (titers < 1:16) to
Rickettsia rickettsii,
Ehrlichia canis,
Babesia canis, and
Babesia gibsoni, and were seronegative to
Anaplasma phagocytophilum,
Anaplasma platys,
Borrelia burgdorferi,
Ehrlichia canis, and
Ehrlichia ewingii by ELISA (SNAP 4Dx PLUS ELISA, IDEXX Laboratories, Westbrook, Maine). The blood from these 24 dogs was PCR-negative after whole blood DNA purification for
Babesia,
Ehrlichia,
Anaplasma,
Rickettsia, hemotropic
Mycoplasma, and
Leishmania spp.
Of the remaining 12 dogs, 4 were seropositive as follows: 1 dog was seroractive to R. rickettsii (IFA titer 1:128), Babesia canis (IFA titer 1:2048), and Babesia gibsoni (IFA titer 1:2048); 1 dog was seroractive to R. rickettsii (IFA titer 1:512) and Babesia canis (IFA titer 1:64); 1 dog was seropositive to Babesia canis (IFA titer 1:1024) and Babesia gibsoni (IFA titer 1:4096); and 1 dog was seropositive to Ehrlichia canis (IFA titer 1:2048). The CVBD PCR and serological testing results were not available for the remaining eight Bartonella spp. IFA seropositive dogs.
Group II consisted of 34 dogs for which diagnostic testing in the NCSU-CVM-VBDDL and NCSU-CVM-IRPL did not provide evidence of the exposure to or infection with a
Bartonella spp.,
Table 2 and
Supplementary Table S1. These sera were used to assess the specificity of the Pap31 recombinant ELISA assays. All of these sera were IFA nonreactive (titers < 1:16) to the three
Bartonella spp.
(B. henselae San Antonio type
2, B. vinsonii subsp
. berkhoffii genotype I
, and
B. koehlerae). All 34 Group II dogs were PCR-negative after whole blood DNA extraction for
Bartonella spp. Whole blood EDTA from 21 of 34 Group II dogs was processed in
Bartonella Alpha Proteobacteria Growth Medium (BAPGM) in the NCSU-CVM-IPRL, as described previously [
14].
Bartonella DNA was not amplified from any extracted blood sample before and after the BAPGM enrichment blood culture.
Bartonella DNA was also not amplified from reagent controls or BAPGM-negative (un-inoculated) culture controls. Bacterial growth was not visualized following the direct culture of blood or subculture from BAPGM onto Trypticase soy agar II (TSA) supplemented with 5% sheep blood plates. Due to the inadequate blood volumes for BAPGM enrichment blood culture, testing results were not available for the remaining 13 dogs.
Sixteen Group II dogs were PCR-negative after whole blood DNA extraction for
Babesia,
Ehrlichia,
Anaplasma,
Rickettsia, hemotropic
Mycoplasma, and
Leishmania spp., and were seronegative to
Anaplasma phagocytophilum,
Anaplasma platys,
Borrelia burgdorferi,
Ehrlichia canis, and
Ehrlichia ewingii by ELISA (SNAP 4Dx PLUS ELISA, IDEXX Laboratories, Westbrook, Maine) or in-house IFA assays. A total of 10 of these 16 dogs were
R. rickettsii seropositive (IFA titer of ≥1:64).
Rickettsia rickettsii is phylogenetically related to the genus Bartonella, belonging to the class alphaproteobacteria [
41]. Therefore, to further assess specificity,
R. rickettsii IFA-positive dogs were included in the Group II dogs. Additional serological testing for the exposure to CVBD organisms was not available for the remaining for 18 dogs.
4.1.2. Human Serum Samples
To evaluate the diagnostic utility of
B. henselae rPap31 and its recombinant fragments (rPap31-NTD, rPap31-MD, and rPap31-CTD) for the serodiagnosis of human Bartonelloses, a set of 36 diagnostic serum samples obtained from the repository maintained by the NCSU-CVM-IPRL (NCSU IRB approval# 1960) were tested by ELISA. Detailed methods for the
Bartonella PCR, BAPGM enrichment blood culture, and the IFA serological panel used to test these study participants were previously published [
19,
42]. Group III consisted of 18 sera from humans naturally exposed to
Bartonella spp. All 18 individuals were
B. henselae, B. koehlerae, or
B. vinsonii subsp
. berkhoffii genotype III seropositive. Fifteen Group III humans were
B. henselae seropositive (IFA titer of ≥1:64),
Table 2 and
Supplementary Table S1. The remaining three individuals were
B. koehlerae and
B. vinsonii subsp
. berkhoffii genotype III IFA seropositive, but were
B. henselae seronegative. Of the 18 people, 8 were IFA seropositive to
B. vinsonii subsp
. berkhoffii genotype I, 10 to
B. vinsonii subsp
. berkhoffii genotype II, 14 to
B. vinsonii subsp
. berkhoffii genotype III, and 11 to
B. koehlerae. A total of 5 of the 18 people were seropositive to all 5 IFA antigens
B. henselae, B. koehlerae, and
B. vinsonii subsp
. Berkhoffii genotypes I, II, and III.
Based on
Bartonella blood qPCR, or droplet digital PCR (ddPCR) [
43] and IFA serology, 14 of 18 Group III humans were
Bartonella PCR-positive and
B. henselae IFA-positive (IFA titer of ≥1:64. A total of 2 of 18 were
Bartonella PCR-positive, but
B. henselae IFA seronegative, and two were
B. henselae IFA-positive and
Bartonella PCR-negative. Of the 16
Bartonella PCR-positive humans, 6 were infected with
B. henselae (blood DNA extraction) and
B. henselae IFA-positive (IFA titer of ≥1:64); 6 were
Bartonella ddPCR-positive (DNA sequencing for species determination was not technically possible) and
B. henselae IFA seropositive; and 1
B. henselae seropositive individual was infected with
B. vinsonii subsp
. berkhoffii genotype I. The three remaining people were
Bartonella ddPCR-positive,
B. henselae IFA-negative, but were seropositive to both
B. koehlerae and
B. vinsonii subsp
. berkhoffii genotype III.
To evaluate ELISA rPap31 proteins for specificity, Group IV sera (n = 18) were selected from the study participants that were
Bartonella spp. IFA-negative and were
Bartonella PCR-negative from blood and BAPGM enrichment blood culture,
Table 2 and
Supplementary Table S1. A total of 9 of these 18 human sera were from healthy donors tested in a previous study [
19]. All Group IV human sera were nonsereoreactive (IFA titer of ≤1:16) to
B. henselae,
B. vinsonii subsp
. berkhoffii genotypes I, II, and III, and
B. koehlerae. Based on the
Bartonella PCR and BAPGM enrichment culture testing,
Bartonella DNA was not amplified from any blood or BAPGM enrichment blood culture.
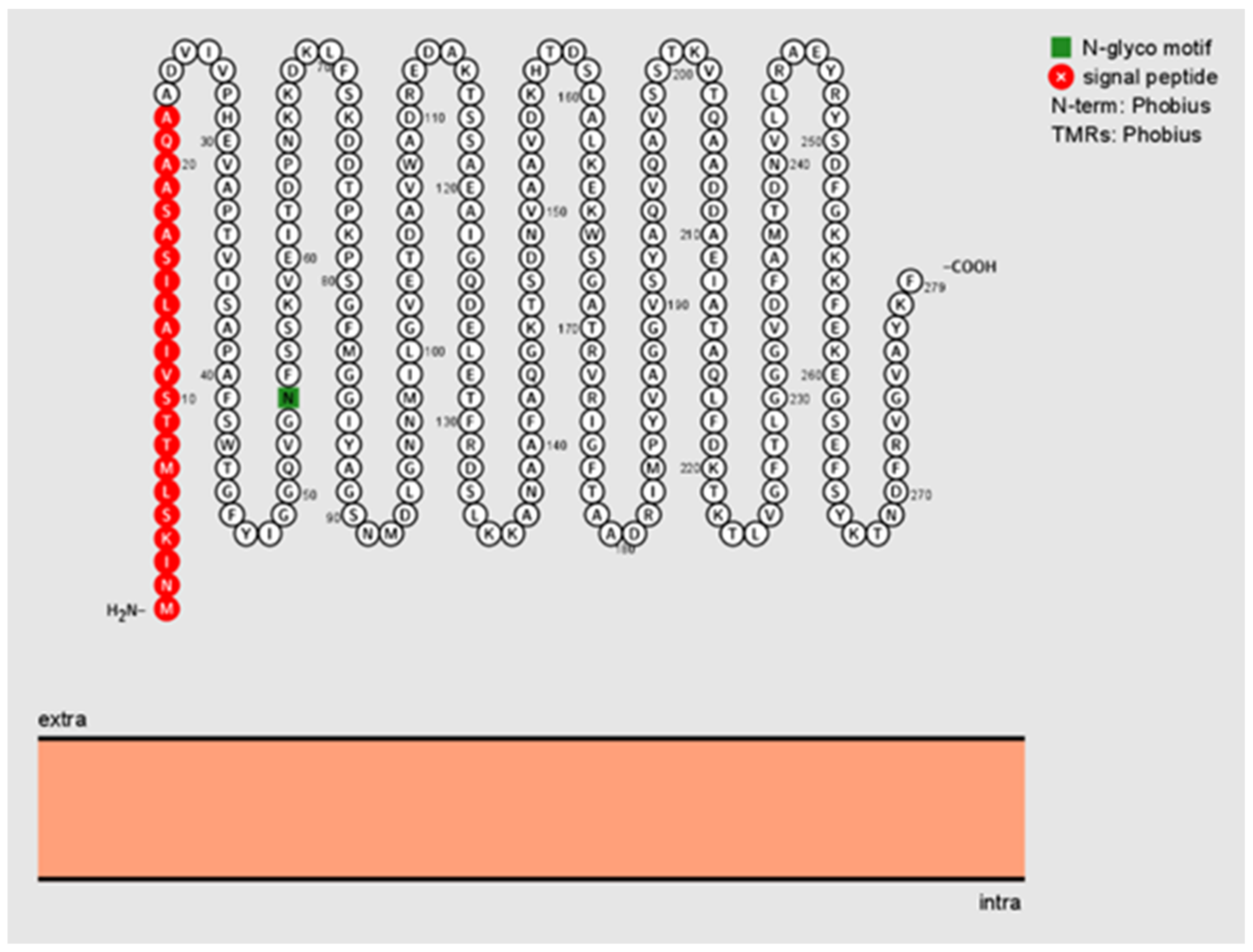
4.2. In Silico Analysis of Bartonella henselae Pap31
The amino acid sequence of the
B. henselae Pap31 protein (1 to 279 amino acids) was derived from the NCBI database. To determine the signal peptide and its cleavage site, signal peptides of the Pap31 protein were determined using the Signal P-5.0 Server (
http://www.cbs.dtu.dk/services/SignalP/ accessed on 8 June 2020). The transmembrane topology of
B. henselae was examined by TMHMM.v2.0 (
http://www.cbs.dtu.dk/services/TMHMM/ accessed on 8 June 2020). PSORTb v3.0.3 was used to predict the Pap31 subcellular localization site of Pap31 (
https://www.psort.org/psortb/ accessed on 8 June 2020). The integration and visualization of annotated and predicted protein sequence features of
B. henselae Pap31 were predicted using Protter (
https://wlab.ethz.ch/protter/start/ accessed on 8 June 2020).
4.3. Amplification of B. henselae pap31 Gene and pap31 Gene Fragments: N-Terminal Domain (NTD), Middle Domin (MD), and C-Terminal Domain (CTD)
The entire
B. henselae San Antonio (
Bh SA2) Pap31 (rPap31) protein and each of the three Pap31 fragments (C-terminal-, middle, and N-terminal domains of Pap31) were cloned and expressed using the
Escherichia coli expression system. Four Pap31 primer sets were used for conventional PCR amplification, as shown in
Table 3. Conventional PCR was performed in a 25 μL final reaction volume containing 12.5 μL of Q5 High-Fidelity 2X Master mix (New England Biolabs, Ipswich, MA, USA, cat. No. M0492S); 0.2 μL of 100 μM of each forward and reverse primer (IDT-DNA Technology); 7.3 μL of molecular-grade water; and 5 μL of DNA from each sample tested. A total of 5 μL of Ultra-Pure, molecular grade water (Genesee Scientific, San Diego, CA, USA) and 5 μL of DNA extracted from
Escherichia coli were used as negative controls. Genomic DNA from
B. henselae Houston-1 was used as a positive control. For all 4 PCR assays, conventional PCR was performed in an Eppendorf Mastercycler EP gradient under the following conditions: a single hot-start cycle at 95 °C for 2 min followed by 30 cycles of denaturing at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 30 s. Amplification was completed by an additional cycle at 72 °C for 2 min. The PCR products were analyzed by 2% agarose gel electrophoresis with detection using gel red (Thermo Scientific, Rockford, IL, USA). Prior to the ligation reaction for cloning, PCR products were purified by gel extraction using PureLink quick gel extraction and a PCR purification combo kit (Invitrogen, Carlsbad, CA, USA).
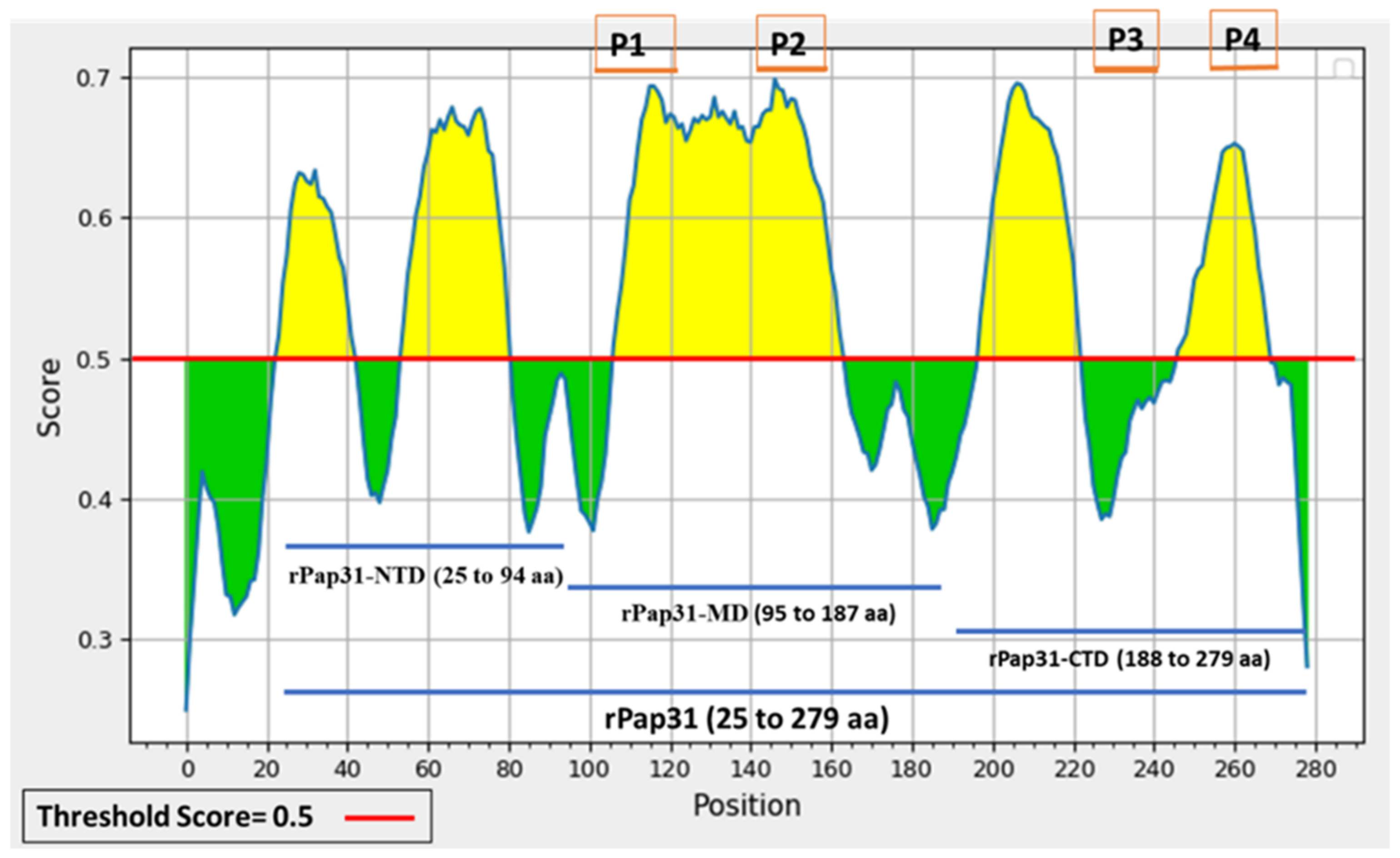
4.4. Cloning of B. henselae Recombinant pap31 Protein and pap31 Gene Fragments: N-Terminal Domain (NTD), Middle Domain (MD), and C-Terminal Domain (CTD)
The
pap31 gene was cloned and expressed using a Champion
TM pET200 Directional TOPO
® Expression kit (Invitrogen, Carlsbad, CA, SUA). Whole
pap31 (encoding 25 to 279 amino acids (aa)) and 3 fragments of the
pap31 gene, C-terminal (25 to 94 aa), middle (95 to 187 aa), and N-terminal (188 to 279 aa) domains, were cloned according to the manufacturer’s instructions (Invitrogen, Carlsbad, CA, USA),
Table 3.
Recombinant plasmids were subsequently transformed into E. coli BL21 (DE3) Star™ chemically competent cells and plated on LB plates containing 50 µg/mL of kanamycin. Positive clones were confirmed by plasmid extraction followed by sequencing. Amplicon products were sequenced using T7 and T7 reverse primers by Sanger sequencing (Genewiz, Research Triangle Park, NC, USA). Chromatogram evaluation and sequence alignment was performed using SnapGene software (GSL Biotech; available at snapgene.com; accessed on 8 June 2020) to confirm the in-frame cloning of pap31 and pap31 gene fragments. The nucleotide sequence homology of the PCR amplicon was performed with available nucleotide sequences at the NCBI databases using the NCBI BLAST program (v2.0).
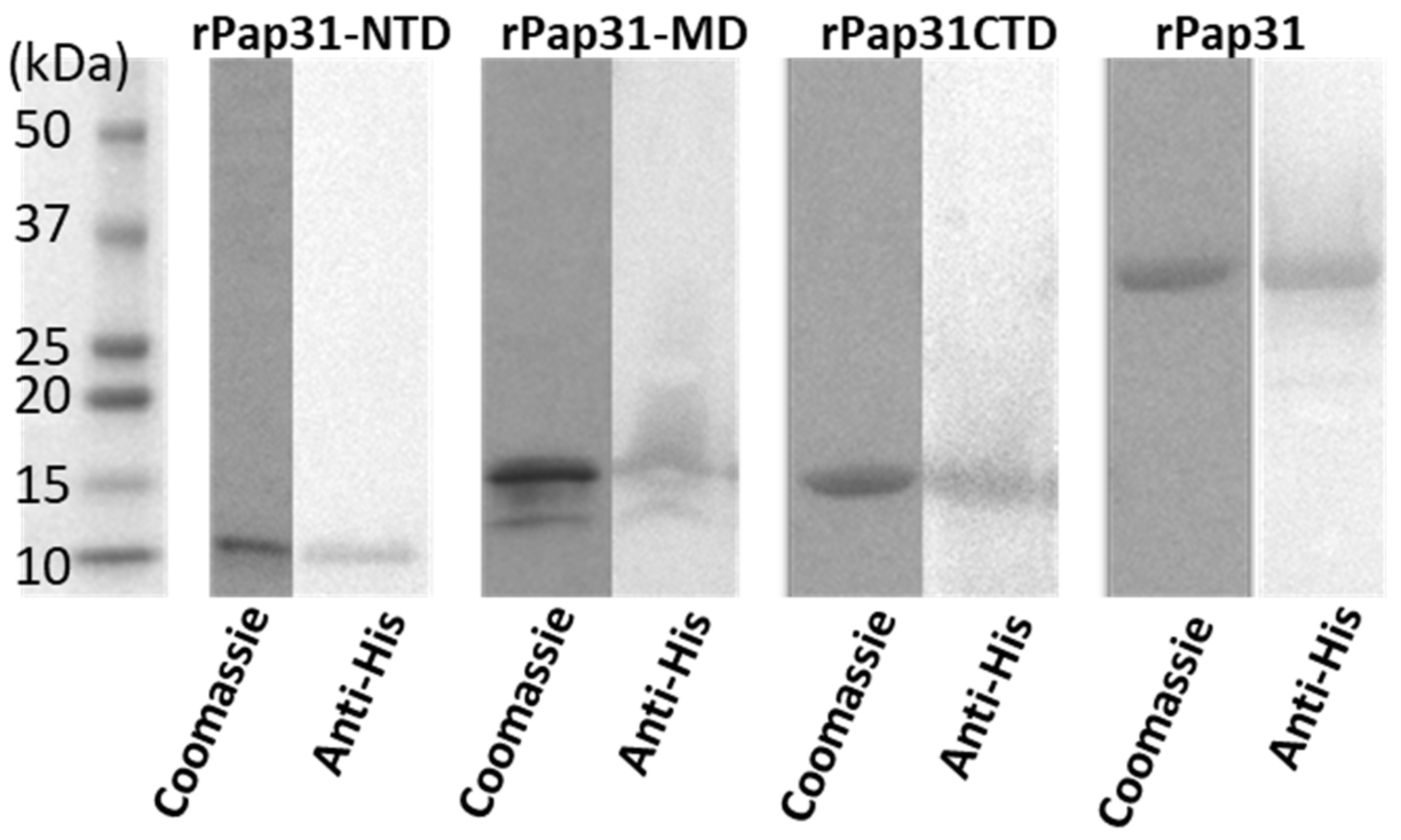
4.5. Expression and Purification of Recombinant Pap31 and Pap31 Domains (N-Terminal, Middle, and C-Terminal Domains)
To confirm and monitor the expression of recombinant proteins, each E. coli BL21 (DE3) clone containing pap31 inserts was grown in 25 mL of LB broth at 37 °C containing 50 µg/mL of kanamycin. When the optical density (OD600 nm) of the culture bacteria reached 0.6 to 0.8, 1mM IPTG was added to induce protein expression. Cultures were examined for protein expression at 0, 3, 6, and 8 h post-IPTG induction at 30 °C by immunoblotting of whole cell lysates using a Pierce™ 6X-His Epitope-Tag mouse monoclonal antibody (Thermo Scientific, Rockford, IL, USA) and an anti-mouse IgG secondary antibody (Rockland, Gilbertsville, PA, USA).
Recombinant rPap31s (rPap31, rPap31-NTD, rPap31-MD, and rPap31-CTD) were purified by column-chromatography using HisPurTM Cobalt spin columns, according to the manufacturer’s instructions (Thermo Scientific, Rockford, IL, USA). For purification, each recombinant clone was grown at 37 °C overnight in a 25 mL LB broth containing 50 µg/mL of kanamycin in a shaker at 200 rpm. After overnight incubation, 5ml of the overnight culture was transferred to fresh 1000 mL LB broth containing 50 µg/mL of kanamycin and was incubated at 37 °C for 2 h in a shaking platform at 200 rpm. After 2 h, IPTG was added to a final concentration of 1 mM and incubated at 30 °C for 8 h. The cultures were then centrifuged at 5000× g to obtain the pellets. The pellets were processed using BugBuster Master Mix (EMD Millipore Corp., Billerica, MA, USA) to purify inclusion bodies according to the manufacturer’s instructions. After the inclusion body purification, the pellets were then subsequently solubilized in 1× PBS containing 6 M of urea. His-tagged proteins were extracted from inclusion body suspensions using HisPur™ Cobalt spin columns (Thermo Scientific, Rockford, IL, USA). Wash buffer (1× PBS with 6 M urea and 5 mM imidazole) and Elution buffer (1× PBS with 6 M urea and 100 mM imidazole) were used during the His spin column purification steps. Eluted proteins were analyzed by SDS-PAGE in Criterion™ using 4–15% gradient polyacrylamide tris-glycine precast midi gels (Bio-Rad, Hercules, CA, USA) at a constant current (100 V) for 1 h 50 min in a 1× running buffer (25 mM tris, 192 mM glycine, 0.1% SDS). A pre stained broad-range (10–250 kDa) molecular weight protein marker (Bio-Rad, Hercules, CA, USA) was used as a standard. Fractionated proteins were visualized by staining the gel overnight with Bio-safe™ Coomassie brilliant blue (Bio-Rad, Hercules, CA, USA). Purified recombinant proteins were verified by WB using Pierce™ 6X-His Epitope-Tag mouse monoclonal antibody (Thermo Scientific, Rockford, IL, USA) and anti-mouse IgG secondary antibody (Rockland, Gilbertsville, PA, USA). Proteins were dialyzed into phosphate-buffered saline (PBS; overnight) using Slide-A-Lyzer™ Dialysis Cassettes, (2–10 kDa MWCO cutoff; Thermo Scientific, Rockford, IL, USA).
4.6. Evaluate the Sensitivity and Specificity of B. henselae Recombinant Proteins Recombinant Pap31 and Pap31 Domains (N-Terminal, Middle, and C-Terminal Domains) by ELISA
The sensitivity and specificity of purified recombinant Pap31 proteins were evaluated by ELISA using sera from Group I (n = 36) and II (control group; n = 34) dogs, and from Group III (n = 18) and IV (control group; n = 18) humans, as described in the Methods Sections for canine and human Bartonelloses, respectively. Each protein was immobilized in duplicate in ELISA plate wells (500 ng/well), using carbonate buffer and standard methods that have been well described [
44]. Sera from naturally infected and unexposed (control) dogs and humans with
Bartonella spp. were used to screen the purified recombinant proteins. In brief, ELISA plate wells were coated with 100 µL of 10 µg/mL recombinant purified proteins using a carbonate buffer, pH 9.6. Plates were incubated at 4 °C overnight. After overnight coating, plates were washed with 1× PBS 4 times. Plates were then incubated with a blocking solution (1× PBS containing 3% milk) at room temperature (RT) for 2 h, followed by washing with 1× PBS containing 0.05% Tween-20 (Bio-Rad, Hercules, CA, USA) 4 times. Plates were incubated with 100 µL of dog or human sera at 1:100 dilution in 1× PBS containing 3% milk at RT for 1 h. Plates were washed with 1× PBS containing 0.05% Tween-20 4 times, followed by incubation with 100 µL of secondary antibody for 1 h at RT. HRP-conjugated goat anti-dog IgG (1:2,000 dilution; Invitrogen, Carlsbad, CA, USA) and HRP-conjugated goat anti-human IgG (1:5,000; Abcam, Cambridge, MA, USA) in 1× PBS containing 3% milk were used as secondary antibodies for dog and human ELISA testing, respectively. After secondary antibody incubation, the plates were washed 5 times with 1× PBS containing 0.05% Tween. A total of 50 µL of 1-Step™ Ultra TMB-ELISA substrate solution (Invitrogen, Carlsbad, CA, USA) was then added to the plate wells and incubated at RT for 15 min, followed by the addition of 2M H
2SO
4 to stop the reaction. The absorbance was measured at 450 nm of the wavelength using a Tecan plate reader. Sera from dogs naturally infected with
B. henselae (
B. henselae IFA titer = 1: 512) and
Bartonella PCR-negative and IFA-negative dogs were used as the positive and negative controls, respectively. The plate wells coated with only coating buffer (without protein) were used as blanks to determine the subtract background noise. Based on the results from the negative controls, a baseline was established for scoring individual samples as positive or negative for infection with
Bartonella. The average absorbance value was calculated for each set of duplicate samples.
4.7. Development of Pap31 B-Cell Epitope-Based ELISA for Bartonelloses
Several B-cell epitope prediction tool methods have been developed over the years, based on a Hidden Markov Model (HMM) with an amino acid propensity scale, Neural Networks, Support Vector Machines, and SVM models trained on the frequency of amino acid pairs (AAPs) [
45,
46,
47]. BepiPred 2.0 with default parameter settings provided by IEDB (Immune Epitope Database and Analysis Resource) were applied to the
B. henselae Pap31 protein sequence to predict the linear B-cell epitopes [
45,
48]. Subsequently, five additional algorithms, AAP [
49], ABCPred [
50], BCPreds [
49], FBCPred [
51], and SVMTriP [
52], were also employed to predict the linear B-cell epitopes of
B. henselae Pap31,
Supplementary Table S2. The molecular weight, amino acid composition, estimated lifetime, and grand average of hydrophobicity (gravy) was determined using the Expasy ProtParam program (
https://web.expasy.org/protparam/ accessed on 20 January 2020). The protein secondary structure of
B. henselae Pap31 was determined using PSIPRED (
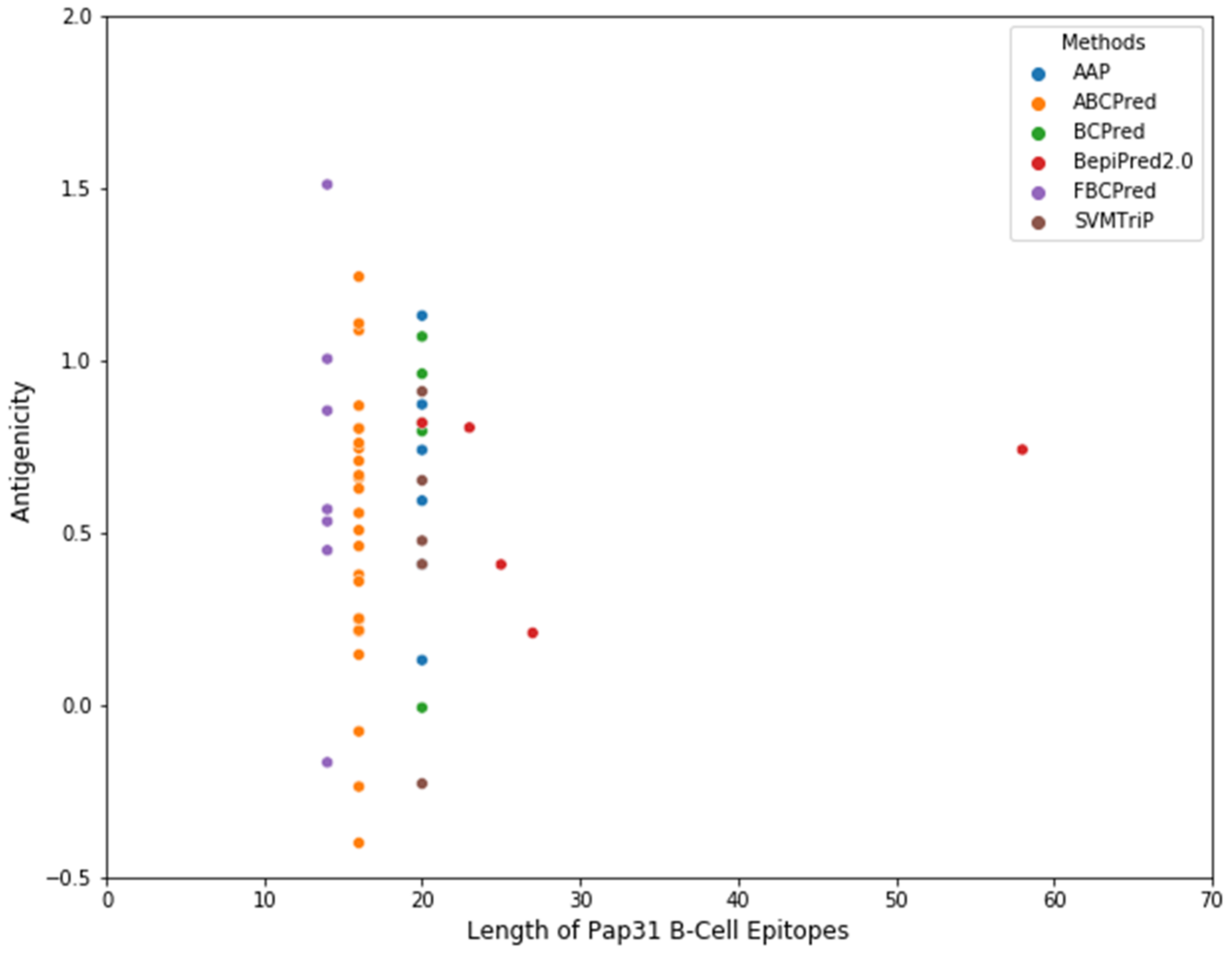
http://bioinf.cs.ucl.ac.uk/psipred/ accessed on 22 January 2020). The antigenicity of the predicted linear B-cell epitopes was evaluated by VaxiJen 2.0. A total of 53 linear B-cell epitopes were predicted by the above-mentioned 6 algorithms,
Figure 9 and
Supplementary Table S2. Based on the predicted linear B-cell epitopes, membrane topology, and antigenicity scores (>0.75), four highly antigenic Pap31 B-cell epitopes (P1, P2, P3, and P4) were selected and synthesized using a commercial company (GenScript, Piscataway, NJ, USA),
Table 4.
To assess the antigenicity and seroreactivity of four Pap31 B-cell epitopes for canine Bartonelloses, Pap31 B-cells epitopes were tested by ELISA and dot blot. Dog ELISA was performed using three serum samples from three experimentally inoculated dogs, five
B. henselae IFA-positive Group I dogs (IFA titer of ≥1:256), and four Group II dogs (
Bartonella PCR-negative and IFA-negative). Twenty-eight-day post-inoculation sera from two dogs that were infected subcutaneously with
B. henselae SA2 and one dog infected with
B. vinsonii subsp
. berkhoffii genotype III [
53], were obtained from NCSU-CVM-IPRL. A total of 1 sera collected on post-inoculation day 21 from a dog intravenously inoculated with
B. henselae CSU 1 strain was kindly provided Dr. Michael Lappin, the Department of Clinical Sciences, Colorado State University. For human testing, sera from eight Group III
B. henselae IFA-positive (IFA titer of ≥1:128) and four Group IV (control) humans were used. Of the eight
B. henselae IFA-positive human samples, two serum samples were from
B. henselae PCR-positive humans.
ELISA was performed, as described above in the Methods Section, with minor modifications. ELISA plate wells were coated with 100 µL of 10 and 20 µg/mL peptides using a carbonate buffer. Sera from dogs and humans were used at two dilutions (1:50 and 1:100 dilution). Secondary antibodies were used as described above in the Methods Section. Dot blot was performed as follows: peptides at a concentration of 15 μg and 20 μg were spotted onto PVDF membranes and air dried. The strips were blocked overnight with 5% nonfat dry milk in 1× TBS containing 0.05% Tween-20 (TBST). After overnight blocking, each strip containing the peptides was incubated with 2 dilutions (1:50 and 1:100) of primary antibody (serum samples from naturally exposed dogs (Bartonella IFA-positive dogs; n = 8) and humans (Bartonella IFA-positive n = 8) with a B. henselae IgG titer of ≥1:128) in 1× TBST containing 3% nonfat dry milk for 1 h. A negative-control dog or negative human serum sample (B. henselae, B. vinsonii subsp. berkhoffii, and B. koehlerae IgG IFA titers < 1:16) were chosen from the VBDDL or IPRL archives, respectively. After washing with 1× TBST, a secondary antibody (alkaline phosphatase (AP)-conjugated goat anti-dog whole IgG or AP-conjugated anti-human) was added at serial dilutions (1:5000 to 1:10,000) in 1× TBS-T containing 3% nonfat dry milk, and then incubated at room temperature for an hour. After washing, the membrane strips were developed using a commercially available substrate solution containing nitroblue tetrazolium chloride (NBT) and 5-bromo-4-chloro-3′-indolyphosphate p-toluidine salt (BCIP; Pierce, Rockford, IL, USA). Image acquisition was performed using a ChemiDoc imaging system (Bio-Rad, Hercules, CA, USA). The blots were analyzed with Image Lab software (v4.1; Bio-Rad, Hercules, CA, USA).
4.8. Statistical Analysis
To determine the sensitivity, specificity, and cutoff values for ELISA, the receiver operating characteristic (ROC) analysis was performed with 95% Cis, as previously described [
54,
55]. Since IFA is considered as a gold-standard assay for the diagnosis of canine and human Bartonelloses, the IFA results were considered as evidence of the exposure to
Bartonella spp. for the calculation of the sensitivity and specificity of ELISA in this study. The optimal density (OD) cutoff values were determined to maximize the Youden index, as previously described (53). The Youden index is the metric for assessing the performance of a diagnostic test. The index is defined by Equation (1), and a, b, c, and d in (1), respectively, denote the numbers of true positives, false negatives, false positives, and true negatives. J = 0 represents a diagnostic test, which gives the same proportion of positives for both the control and infected groups, and J = 1 represents a diagnostic test with no false positive and false negative detected. The optimal cutoff value is the one that maximizes the
J.
To compare the agreement between the ELISA and
Bartonella IFA testing results, the positive, negative, and overall percent agreement between ELISA and IFA assays were calculated as previously described [
56]. To measure the level of agreement between the
Bartonella IFA and ELISA tests, the kappa statistic was calculated as previously described [
57]. Differences in the IgG reactivity to target proteins between the infected and control groups were analyzed using the Mann–Whitney U test. The scatterplots and ROC curve analysis for ELISA seroreactivity were generated in the Windows 10 operating system with the help of the Anaconda Navigator, v1.9.12. The scatter plots were generated using Python 3. 6.13 in Jupyterlab 3.2.1 and the ROC curves were analyzed using R v3.6.1 in R studio 1.1.456 (accessed on November 25, 2021). The
p-values of less than 0.05 were considered statistically significant.


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}