Abstract
The early diagnosis of Helicobacter pylori infection is important for gastric cancer prevention and treatment. Although endoscopic biopsy is widely used for H. pylori diagnosis, an accurate biopsy cannot be performed until a lesion becomes clear, especially in pediatric patients. Therefore, it is necessary to develop convenient and accurate methods for early diagnosis. FlaA, an essential factor for H. pylori survival, shows high antigenicity and can be used as a diagnostic marker. We attempted to identify effective antigens containing epitopes of high diagnostic value in FlaA. Full-sized FlaA was divided into several fragments and cloned, and its antigenicity was investigated using Western blotting. The FlaA fragment of 1345–1395 bp had strong immunogenicity. ELISA was performed with serum samples from children by using the 1345–1395 bp recombinant antigen fragment. IgG reactivity showed 90.0% sensitivity and 90.5% specificity, and IgM reactivity showed 100% sensitivity and specificity. The FlaA fragment of 1345–1395 bp discovered in the present study has antigenicity and is of high value as a candidate antigen for serological diagnosis. The FlaA 1345–1395 bp epitope can be used as a diagnostic marker for H. pylori infection, thereby controlling various gastric diseases such as gastric cancer and peptic ulcers caused by H. pylori.
1. Introduction
Helicobacter pylori (H. pylori) is a pathogen that infects approximately 50% of the global population [1] and colonizes the epithelial surface or the surface mucus of the gastric mucosa [2]. It is known to increase the risk of gastric cancer by causing chronic inflammation and peptic ulcers [1,3,4,5,6]. Therefore, H. pylori was defined as a class 1 carcinogen by the World Health Organization (WHO) in 1994. According to a report released by the International Agency for Research on Cancer in 2020, H. pylori was responsible for the most infectious pathogen-related carcinogenesis in 2018 (https://gco.iarc.fr/causes/infections; accessed on 11 December 2022). Therefore, the early diagnosis of H. pylori infection is of great importance for the early diagnosis of gastric cancer. Early diagnosis is also important for developing a preventive method for peptic ulcers and gastric cancer based on the risk assessment induced by H. pylori infection [7,8]. In addition, given that most H. pylori infections are established in childhood [9,10,11,12], it is necessary to develop novel diagnostic methods applicable to pediatric patients.
Endoscopic biopsy is the most widely used method for diagnosing H. pylori infection [7,8]. The sensitivity of this method varies depending on the operator’s proficiency. In addition, not all H. pylori infections lead to gastric lesions, such as gastritis or peptic ulcers [5,13]; therefore, accurate biopsy is difficult. In pediatric patients, an endoscopic biopsy is difficult to perform. Moreover, in pediatric patients, an endoscopic biopsy is not appropriate as an early diagnostic method for H. pylori infection, considering that most H. pylori infections are established at a young age and there are numerous cases where gastric lesions are not observed, even after infection [5,8,10,11,13,14,15].
Serological diagnosis may be an alternative method for resolving these issues [14]. It could also be an appropriate tool for children in whom gastric lesions have not been observed. However, there is also a limitation in applying serological tools for diagnosing H. pylori infection, as the useful diagnostic antigens of H. pylori have not been identified [5,13,14,16,17]. Moreover, the highly heterogeneous characteristics of virulence factors known to be associated with host immune responses make it difficult to find useful antigen candidates for enzyme linked immunosorbent assay (ELISA) [4,13,17,18,19].
Thus, it is necessary to identify specific H. pylori antigens that show high immunogenicity to develop a more useful and convenient method for serological diagnosis. FlaA is a necessary protein for the survival of H. pylori in the gastric environment [17,20] and is also known to have high immunogenicity [21]. Therefore, this study investigated the effective antigens in the FlaA to develop an ELISA method for the early diagnosis of H. pylori infection as a novel diagnostic approach that can also be applied to pediatric patients.
2. Materials and Methods
2.1. Bacterial Strain
Helicobacter pylori strain 51 (H. pylori 51), isolated at Gyeongsang National University Hospital from a patient with a gastric disorder, was obtained from the H. pylori Korean Type Culture Collection (College of Medicine, Gyeongsang National University, Korea; 2006–2015). H. pylori 51 was grown on Brucella agar (Difco, Franklin Lakes, NJ, USA) containing 10% bovine serum (Difco) at 37 °C under 10% CO2 and 100% humidity [22,23]. All Escherichia coli strains for recombinant protein expression were cultured in Luria-Bertani (LB) broth (Invitrogen, Waltham, MA, USA) containing 1 mM ampicillin (Sigma, St. Louis, MO, USA) at 37 °C.
2.2. Serum Samples of Patients
Sera from pediatric patients with (n = 60) and without (n = 220) H. pylori infection were collected at Gyeongsang National University Hospital (GNUH, Jinju, Korea). Biopsy specimens were smeared onto Brucella agar plates containing 10% bovine serum, vancomycin (Sigma, 10 μg/mL), nalidixic acid (Sigma, 25 μg/mL), and amphotericin B (Sigma, 1 μg/mL) to culture H. pylori [22,23]. The bacteria were identified as H. pylori based on the 16s rRNA PCR using primers of PV31 (5′-CGGCCCAGACTCCTACGGG-3′) and PV32 (5′-TTACCGCGGCTGCTGGCAC-3′) [24]. A campylobacter-like organism (CLO) test was also conducted to measure urease activity using three biopsy samples from the same patient according to a previously described method [25]. Urease activity was determined over 24 h based on the time of color change of the broth. Sixty serum samples from patients with H. pylori infection were divided into two groups based on the results of the CLO test: weak (color change within 6–24 h, n = 30) and strong (color change within 6 h, n = 30). All sera were stored at −70 °C until further use. All biopsy and serum samples were collected under parental consent and provided by Gyeongsang National University Hospital, a member of the National Biobank of Korea, after obtaining permission from the hospital ethics committee (GNUHIRB-2016-04-003-001).
2.3. PCR Amplification of H. pylori FlaA Fragments and Cloning
Total genomic DNA was obtained from H. pylori 51 using the QiaAmp DNA Mini Kit, according to the manufacturer’s instructions (Qiagen, Hilden, Germany). According to the GenBank database, full-length flaA (1533 bp) was found to include NdeI (gagttacacat) and BglII (cccgggagat) in the N- and C-terminal regions, respectively (Supplementary Data S1). The primers for amplification of full-length flaA were designed based on published alignments available through GenBank (Table 1). The amplified full flaA gene was cloned into the pGEM-T vector. The pGEM-T vector was digested with NdeI and BglII, and the fragment of the full flaA gene was purified and retrieved from an agarose gel after electrophoresis. Purified flaA was cloned into the pET15b vector and treated with NdeI and BglII (pET15b/flaA). For the 1–772 bp fragment (Frag 1) of the full flaA gene, the pET15b/flaA vector was digested with BamHI and BglII and ligated (pET15b/Frag 1). For the flaA 778–1533 fragment (Frag 2), PCR was performed using the template of total genomic DNA of H. pylori 51, a forward primer with the NdeI recognition sequence and the BamHI recognition site, and a reverse primer used for amplification of the full flaA gene. The cloning process was performed in the same way as that for full flaA gene cloning.

Table 1.
Oligonucleotide primers for PCR amplification of flaA and its fragments.
Frag 2, which showed higher antigenicity than Frag 1, was divided into subfragments of 200–230 bp with 50% overlap. Fragments showing relatively high antigenicity were selected among subfragments of 200–230 bp and cloned as subfragments of 100 bp in size. The 100 bp subfragments showing the highest antigenicity were finally divided into 50 bp subfragments. Antigenicity analysis was performed using Western blotting. The primers used to prepare each subfragment are listed in Table 1. Primers for amplifying all subfragments except Frag 2–2 were designed to include EcoRI and XhoI recognition sequences in the F- and R-primers, respectively. The primers used for Frag 2–2 amplification were designed to contain BamHI and XhoI recognition sites in F- and R-primers, respectively.
All amplified subfragments were cloned into the pGEMT/A vector. These vectors were digested with EcoRI/XhoI or BamHI/XhoI and subsequently purified from an agarose gel after electrophoresis. The purified subfragments were cloned into the pEGexp vector and treated with EcoRI, XhoI, or BamHI and XhoI. The pEGexp vector was constructed based on the pET15b vector, which includes an N-terminal leader consisting of the amino acid sequence of GroEL T1 (1–772 bp of the H. pylori GroEL gene) and chaperonin [26]. GroEL T1 is highly expressed in E. coli and has a HindIII recognition sequence at the 771 bp position of the gene without any reactivity to the sera of H. pylori-infected patients (Figure 1). Thereafter, cloned genes were transformed into competent E. coli BL21 cells for expression [27].
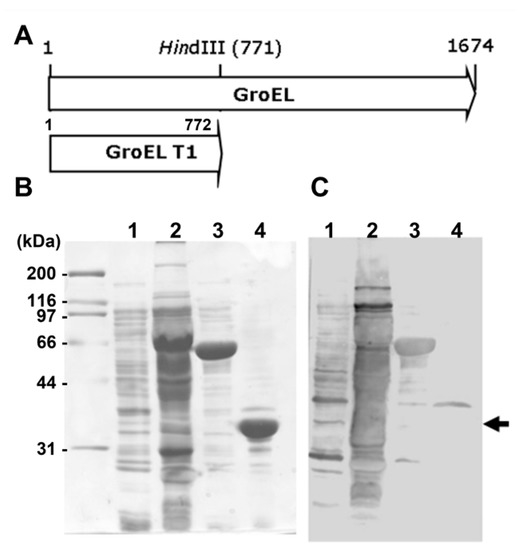
Figure 1.
Immunoblot analysis of recombinant proteins of GroEL and GroEL T1. (A) The map of GroEL and GroEL T1 genes of H. pylori. (B) SDS-PAGE and (C) Western blot analysis using H. pylori-positive pooled sera. Black arrows indicate GroEL T1 (772 bp) recombinant protein, which did not show any reactivity to H. pylori-positive pooled sera. Line 1, whole lysate of E. coli containing pET15B; line 2, whole lysate of H. pylori 51; line 3, recombinant protein of GroEL; line 4, recombinant protein of GroEL T1.
The PCR was performed in a PCR PreMix tube (Accupower PCR PreMix, Bioneer, Daejeon, Korea) containing 250 µM dNTP, 1 U of Taq DNA polymerase, 10 mM Tris-HCl (pH 9.0), 40 mM KCl, 40 mM MgCl2, a stabilizer tracking dye, 40 ng of the template, and 2 pmol of primers, using a PCR Thermal Cycler (Takara, Kusatsu, Japan). PCR amplification was performed under the following conditions: 94 °C for 10 min; 35 cycles of 94 °C for 1 min, 50 °C for 1 min, and 72 °C for 2 min; and 72 °C for 10 min. The amplified PCR products were subjected to electrophoresis on a 1% agarose gel and purified using a QIAquick Gel Extraction Kit (Qiagen) [27]. All restriction enzymes were obtained from Roche Holding AG (Basel, Switzerland).
2.4. Expression and Purification of Recombinant Proteins
E. coli BL21 cells were grown in LB broth until the culture reached an optical density (OD600) of 0.6–0.8. Thereafter, isopropyl β-Ɗ-1-thiogalactopyranoside (1 mM; Duchefa Biochemie, Haariem, Netherlands) was added and subsequently incubated for 4 h. Thereafter, harvested cells were resuspended and lysed by sonication in a lysis buffer (50 mM Tris-hydrogen chloride, 100 mM sodium chloride, pH 8.0). The lysates were collected as supernatants by centrifugation at 5000× g for 30 min. The recombinant proteins were purified using His-Tag purification column, according to the manufacturer’s instructions (Bio-Rad, Hercules, CA, USA). All purified recombinant proteins were stored at –20 °C until use.
2.5. Immunoblot Analysis of the Recombinant FlaA Fragments
Purified recombinant proteins were separated using 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and stained with Coomassie blue R-250 (Bio-Rad) or with Ponceau S (ThermoFisher, Waltham, MA, USA) after electrotransformation onto nitrocellulose membranes (Bio-Rad). For Western blot analysis, recombinant proteins were electrotransferred onto nitrocellulose membranes, which were immunoblotted with pooled 300 H. pylori-positive sera verified previously at a 1:200 dilution [28] and an alkaline phosphatase-conjugated goat anti-human IgG (H + L) at a 1:500 dilution (Bio-Rad). Specific reactions were detected using an AP-conjugated substrate kit (Bio-Rad). The reactivity of immunoblot was compared and graded using the mean gray value function of the ImageJ software (1.53e version) after background compensation. The relative antigenicity values are the ratio of Western blot reactivity to the protein staining levels in SDS-PAGE (loading control).
2.6. Enzyme Linked Immunosorbent Assay Using an Antigenic Determinant of FlaA
The determined antigen (10 µg/mL) was coated in 96-well microplates at 4 °C for 16 h for the ELISA. Following blocking at 37 °C for 3 h using 3% bovine serum albumin fraction V (BSA; Fisher Scientific, Hampton, NH, USA) in PBS containing 0.05% Tween 20 (PBST; Sigma), the plates were incubated with sera (1:100 diluted with 1% BSA in PBST) from patients at 37 °C for 1 h. Thereafter, they were incubated with horseradish peroxidase-conjugated goat anti-human IgG or IgM (1:10,000 diluted with 1% BSA in PBST; Bethyl, Montgomery, TX, USA) at 37 °C for 1 h. Between each step, plates were washed with PBST. Color development was performed by incubating the samples with o-phenylene diamine as a substrate and subsequently stopped with 2 N sulfuric acid, after which the microplates were read at 492 nm using an Emax Precision microplate reader (Molecular Devices, San Jose, CA, USA) [26].
2.7. Statistics
Data are presented as the mean ± standard deviation (SD). Statistical significance was determined with the Student’s t-test using Statistical Package for Social Sciences software (version 24, SPSS, Chicago, IL, USA). Differences were considered significant if p < 0.05.
3. Results
3.1. Antigenic Determination of H. pylori FlaA
The pooled sera from H. pylori-infected patients showed strong reactivity against the FlaA whole protein on Western blot analysis (Figure 2). The IgG in pooled sera was also bound to Frag 1 and 2; however, it showed a stronger reactivity to Frag 2 than to Frag 1. To effectively produce recombinant proteins and soluble subfragment proteins of Frag 2, they were cloned into the pEGexp vector. The resulting recombinant proteins were expressed and conjugated with the GroEL T1 protein, which is the N-terminal 771 bp portion of the H. pylori GroEL protein. On Western blot analysis using pooled sera from H. pylori-infected patients, the GroEL T1 protein did not show reactivity (Figure 1). Strong reactivity of pooled sera from H. pylori-infected patients was observed against 200–230 bp subfragments of Frag 2: Frag 2–5 (1195–1395 bp) and 2–6 (1297–1533 bp). Between Frag 2–5 and 2–6, a 100 bp subfragment that showed high reactivity with IgG in the sera was Frag 2–5–2 (1297–1395 bp). In Frag 2–5-2, the 50 bp subfragment that exhibited the highest reactivity was Frag 2–5–2–2 (1345 –1395 bp, ISTVNNISITQVNVKAA), which was selected as the effective antigen for ELISA (Table 2, Figure 3 and Figure 4, and Supplementary Data S2).
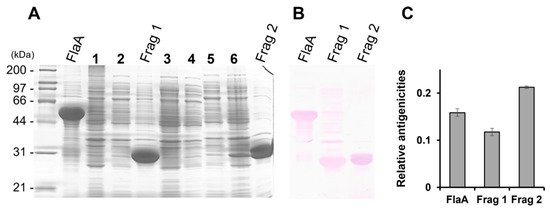
Figure 2.
(A) SDS-PAGE and (B) Western blot analysis of the recombinant proteins, FlaA, Frag 1, and Frag 2. (C) Normalized relative antigenicity of each recombinant protein. All recombinant proteins showed reactivity to H. pylori-positive pooled sera. The positive pooled sera showed a stronger reactivity to Frag 2 than to Frag 1. 1 and 2, uninduced FlaA; 3 and 4, uninduced Frag 1; 5 and 6, uninduced Frag 2.
Table 2.
Measured relative antigenicity of the recombinant subfragment proteins of FlaA.

Figure 3.
Immunoblot analysis of the recombinant subfragment proteins of FlaA. (A) Ponceau S staining and (B) Western blot analysis using H. pylori-positive pooled sera. High levels of relative reactivities were observed in Frag 2–5 (1195–1395 bp), Frag 2–6 (1297–1533 bp), Frag 2–5–2 (1297–1395 bp), and Frag 2–5–2–2 (1345–1395 bp).
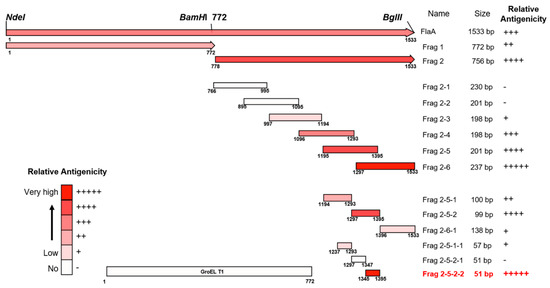
Figure 4.
Maps of antigenic domains of FlaA. Except for Frag 1 and Frag 2, all subfragment proteins of FlaA were expressed by conjugation with GroEL T1 protein (772 bp).
3.2. Analysis of the Reactivity of H. pylori-Infected Pediatric Patients Antibodies to the FlaA Fragment 2–5–2–2
The Frag 2–5–2–2 was purified and used as an ELISA antigen for the diagnosis of H. pylori infection. ELISA was performed with sera from H. pylori-infected patients (CLO weak group, n = 30; CLO strong group, n = 30) and non-infected children (CLO negative group, n = 220). The average absorbance of IgG ELISA was 0.25 ± 0.29, 0.93 ± 0.27, and 0.89 ± 0.33 in the sera of non-infected, CLO weak, and CLO strong groups at 492 nm, respectively (Figure 5, p < 0.000). In the case of IgM ELISA, the average absorbance was 0.09 ± 0.08, 1.04 ± 0.31, and 1.02 ± 0.37 in the sera of non-infected, CLO weak, and CLO strong groups, respectively (Figure 6, p < 0.000). No significant differences were observed between the weak and strong CLO groups. When the serum samples were divided into non-infected and H. pylori-infected (CLO positive) groups, the cut-off value of 0.53 for IgG ELISA was determined via receiver operating characteristic (ROC) analysis, with 90.0% sensitivity and 90.5% specificity, whereas the cut-off value of 0.35 for in IgM ELISA was determined via ROC analysis with 100% sensitivity and specificity.
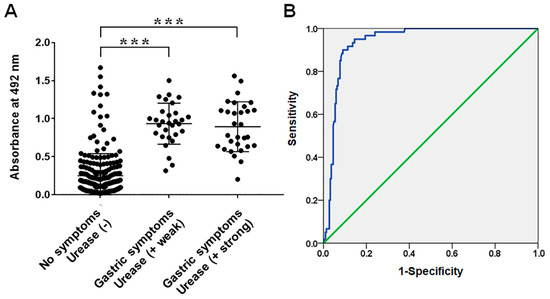
Figure 5.
(A) IgG reactivities of sera from infant patients with/without H. pylori infection measured via ELISA using recombinant protein Frag 2–5-2–2 of FlaA. (B) Receiver operating characteristic (ROC) curves of ELISA using Frag 2–5-2–2 for the diagnosis of H. pylori infection. The area under the ROC of Frag 2–5-2–2 ELISA was 0.939 (95% confidence interval; *** p < 0.000).
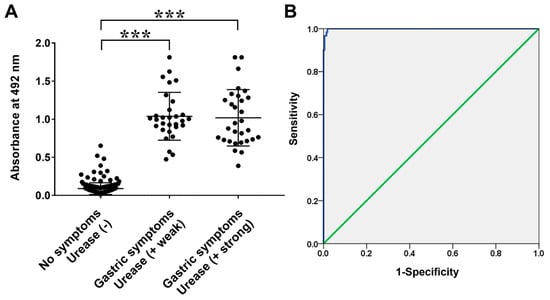
Figure 6.
(A) IgM reactivities of sera from infant patients with/without H. pylori infection measured via ELISA using recombinant protein Frag 2–5-2–2 of FlaA. (B) Receiver operating characteristic (ROC) curves of ELISA using Frag 2–5-2–2 for the diagnosis of H. pylori infection. The area under the ROC of Frag 2–5-2–2 ELISA was 0.999 (95% confidence interval; *** p < 0.000).
4. Discussion
H. pylori is a highly heterogeneous bacterium with various virulence factor antigens that may be involved in the pathogenicity and immune responses [4,13,17,18]. Several virulence factors have been suggested as potential diagnostic antigens [17,29,30,31,32]. However, not all virulence factor antigens are present in all H. pylori strains. Indeed, the existence and expression of several virulence factors differ depending on the strain distribution in different areas. Several previous studies have reported geographical differences in the prevalence of H. pylori virulence factors [4,6,11,15,16,17,19,33,34,35,36,37]. The expression of these virulence factors can be regulated via their own modulatory system according to changes in environmental conditions [5,38,39,40]. These results imply that different types of antigens are exposed to the immune system of the host at various levels, depending on the environment in which H. pylori adapts. Moreover, not all virulence factor antigens induce an immune response following infection [13].
Flagellar antigens have a high possibility of being diagnostic antigens because the flagella play an important role in bacterial survival in the gastric environment [17,20]. After entering the stomach of the host, H. pylori must neutralize the hostile acidic conditions at the beginning of infection [41,42] and move towards the gastric epithelium of the host to establish successful colonization [17,20]. Urease and the flagella mediate acid neutralization and bacterial motility, respectively [17,20,41,42]. Of these, urease is an enzyme whose expression level may vary depending on the characteristics of the strain and environmental pH; however, as flagellar antigens are structural proteins, it is considered more valuable as diagnostic antigens [43,44]. Therefore, this study focused on the antigenicity of the flagella.
Flagellar antigens are essential structural proteins for the survival and colonization of H. pylori; therefore, they might be exposed to the immune system of the host continuously, thereby inducing specific immune responses including humoral immunity [17,45]. These results are consistent with those of a previous study demonstrating that mice immunized with flagellar sheath proteins exhibited significantly reduced H. pylori colonization [20]. Moreover, the high antigenicity of flagella-related proteins was reported in previous studies, which suggested that the flagella antigen is a suitable diagnostic and vaccine target [21,32,46]. The flagellar filament consists of two flagellins (FlaA and FlaB) encoded by flaA and flaB. Between FlaA and FlaB, FlaA has been suggested as a marker for the presence of H. pylori infection and gastric cancer [32,46]. Altogether, previously reported characteristics of FlaA show a high possibility of being a specific antigen required for the serological diagnosis of H. pylori. Therefore, in the current study, we identified a specific antigen in the FlaA protein and analyzed whether the identified antigen could be used in ELISA for the diagnosis of H. pylori.
There have been several attempts to use FlaA as a diagnostic antigen [32,46]. However, the sensitivity and specificity of the diagnostic methods depend on the ability of a specific antigen to induce the most specific and detectable immune responses [47,48]. Considering that the FlaA size is 56.7 KDa and that the size of the antigen in major histocompatibility complex (MHC) I or MHC II does not exceed 30 amino acids, the large size of FlaA (511 amino acids) should be fragmented to determine the specific antigen that has a high serological diagnostic value without a non-specific reaction. Therefore, in the current study, the flaA gene was divided into pieces, step-by-step, and each of them was cloned to produce recombinant proteins (Figure 4). While the recombinant proteins of full-sized FlaA, Frag 1, and 2 were expressed in the soluble form, the others were not. Therefore, other fragment antigens were expressed by binding them to the GroEL T1 protein of H. pylori to produce a soluble form [26]. The H. pylori groEL gene has a high expression level after transformation into E. coli and it has a HindIII restriction enzyme site at the 771 bp position, thereby easily removing the sequence below the 771 bp position. The recombinant protein of the N-terminal 1–772 bp of H. pylori GroEL (GroEL T1) did not show antigenicity on Western blotting using pooled sera from H. pylori-infected patients. All recombinant proteins conjugated with GroEL T1 were subjected to Western blotting using pooled sera of H. pylori-infected patients, thereby identifying the Frag 2–5-2–2 (1345–1395 bp, ISTVNNISITQVNVKAA) of having major antigenicity. As mentioned above, it is very important that the antigen used for diagnosis is a conserved antigen of H. pylori. Diagnostic methods using an antigen of a variable region have many disadvantages in their sensitivity and specificity. The Frag 2–5–2–2 was found to be a highly conserved antigen according to the NCBI database.
The recombinant protein Frag 2–5–2–2 showed specific immune responses in the sera of H. pylori-infected patients (CLO positive groups) as a diagnostic antigen in ELISA. The IgG ELISA OD values of the H. pylori-infected and non-infected groups were 0.91 ± 0.30 and 0.25 ± 0.29, respectively (Figure 5, p < 0.000). For IgM, OD values of 1.03 ± 0.34 and 0.09 ± 0.08 were observed in the H. pylori-infected and non-infected groups with a significant difference, respectively (Figure 6, p < 0.000). Moreover, ROC analysis showed a cut-off value of 0.53 for IgG ELISA with 90.0% sensitivity and 90.5% specificity, and a cut-off value of 0.35 for IgM ELISA, with 100% sensitivity and specificity. These results show that the determined antigen in the present study has high antigenicity and is an important candidate antigen for serological diagnosis, especially in pediatric patients. There were no significant differences in the OD values of IgG and IgM between the weak and strong CLO groups. It appears to be due to the different expression levels of ureA and ureB according to the environment of survival and colonization of H. pylori [43,44].
H. pylori infection increases the risk of gastric cancer, the fourth leading cause of cancer-related deaths worldwide. However, when gastric cancer is diagnosed early, the five-year relative survival rate increases [49,50]. Therefore, the early diagnosis of H. pylori infection can provide important clues for the prevention and treatment of gastric cancer. Moreover, H. pylori infection increases the risk of gastritis and gastric ulcers; therefore, early diagnostic tools for H. pylori infection with high sensitivity and specificity, before the development of clinical symptoms, are required to effectively overcome gastric diseases.
As major diagnostic methods, gastroscopy and biopsy have low sensitivity when considering that many cases of H. pylori infection are established in childhood without any clinical symptoms. Moreover, they are stressful for pediatric patients. As a non-invasive diagnostic method, the urea breath test and stool antigen test have also been used in clinical practice. The urea breath test is an exhalation investigation method to measure the hydrolysis of 13C or 14C isotope-labeled urea by H. pylori [51,52]. However, as the cutoff value of this method has not been determined, the results need to be validated differently considering the targeted population [51,53,54]. In addition, because the urea breath test requires expensive equipment, it is difficult to use widely due to its high cost [51]. The stool antigen test is a diagnostic tool based on H. pylori antigen detection in feces. It can be used for patients who cannot use the urea breath test, such as pediatric or asthmatic patients [55,56,57,58]. However, the accuracy of this diagnostic method can be affected by various gastrointestinal factors, such as bleeding ulcers, antibiotic treatment, and bismuth-containing compounds [57,59]. Therefore, it is necessary to develop a simple and accurate method for the early diagnosis of H. pylori infection that can be applied to pediatric patients.
The purpose of the present study was to discover a specific antigen of FlaA that can be used in ELISA for the serological diagnosis of H. pylori infection, which can be easily applied to all patients and is convenient for application in children. Previous studies showed different results regarding the sensitivity of the detection of IgG and IgA for H. pylori diagnosis [51,60,61,62,63]. Nevertheless, the measurement of IgG levels in sera can be said to be the gold standard for diagnosis [62,63,64]. Furthermore, considering the possible use of the FlaA Frag 2–5–2–2 as a diagnostic antigen for further multi-epitope ELISA, as indicated by the results of the current study, the IgG results will be more useful than those of IgA. Therefore, including IgM, our study focused on the IgG responses to the discovered recombinant antigens.
Among 220 sera samples in the non-infected group, 21 samples showed higher OD values of IgG ELISA than the cut-off value. Of these, ten samples showed values higher than 0.91 ± 0.30, the average IgG ELISA OD of sera samples from infected patients. Misdiagnosis of H. pylori infection is considered the primary reason for these high OD values measured in the sera of some patients in the non-infected group. The negative pediatric sera we used were collected from patients diagnosed to be negative for H. pylori through endoscopic biopsy. However, in some pediatric patients, H. pylori might not be accurately diagnosed. Therefore, even though they were positive, an endoscopic biopsy could be used to diagnose them as being negative for H. pylori. According to a previous study, the sensitivity of endoscopic biopsy in Korea has been estimated to be 69% [65]. It can be expected that true positive sera may be included in the non-infected group. For accurate identification of the reason, follow-up medical records are required. However, since there was no record of subsequent diagnosis, we could not confirm whether the patients of the non-infected group showing high OD levels in sera were subsequently diagnosed to be H. pylori positive. Another possibility is a non-specific reaction to Frag 2–5–2–2. Commonly, serological tests of ELISA cannot exclude non-specific responses completely. However, considering that the size of antigens associated with reactivity, except for the conjugated GroEL T1, is 50 bp, the possibility of a non-specific reaction seemed low. Moreover, most sera in the non-infected group showed lower OD values than the cut-off value. Therefore, it is believed that the misdiagnosis of H. pylori infection caused noise in the results by disturbing sample grouping.
The antigen of Frag 2–5–2–2 (1345–1395 bp, ISTVNNISITQVNVKAA) in FlaA was discovered in this study, and the ELISA results showed its potential as a diagnostic antigen. The Frag 2–5–2–2 can be used as an antigen in diagnostic kits for H. pylori infection, thereby helping control various gastric diseases caused by H. pylori. Further studies are needed to discover other effective antigens; thus, it will be possible to develop ELISA or immunochromatography kits with improved sensitivity and specificity based on the multi-epitopes.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/pathogens11121544/s1. Supplementary Data S1. FlaA full sequence. Supplementary Data S2. Immunoblot analysis of the recombinant subfragment proteins of FlaA. (A) Ponceau S staining and (B) Western blot analysis using H. pylori-positive pooled sera. Frag 2–13 is a subfragment recombinant protein of FlaA 1237–1347 bp. Because Frag 2–13 was divided into Frag 2–5-1–1 and Frag 2–5-2–1 to analyze the antigen reactivity in the present study, Frag 2–13 has not been explained.
Author Contributions
Conceptualization, M.-J.C., W.-K.L. and M.J.; Methodology, J.-I.S., W.B. and J.-S.P. (Jin-Sik Park); Validation, H.-E.P.; formal analysis, H.-E.P., S.P. and D.N.; Investigation, H.-E.P., S.P., D.N. and M.J.; Resources, J.-S.J., J.-H.S., J.-S.P. (Ji-Shook Park) and H.-S.Y.; Data curation, H.-E.P., S.P. and M.J.; Writing—original draft preparation, H.-E.P., S.P. and M.J.; Writing—review and editing, M.-K.S., S.-C.B., H.-L.K., W.-K.L. and M.J.; Supervision, W.-K.L. and M.J.; Project administration, M.J.; Funding acquisition, W.-K.L. and M.J.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Research Foundation of Korea (NRF) grants funded by the Korean government (2019R1I1A3A01059312 and 2021R1I1A3059179) and a grant from the Institute of Health Science, Gyeongsang National University (2021).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Gyeongsang National University Hospital, a member of the National Biobank of Korea, after obtaining permission from the hospital ethics committee (GNUHIRB-2016-04-003-001).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data supporting the conclusions of this article are provided within the article. The original data in the present study are available from the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Malaty, H.M. Epidemiology of Helicobacter pylori infection. Best Pract. Res. Clin. Gastroenterol. 2007, 21, 205–214. [Google Scholar] [CrossRef]
- Blaser, M.J.; Atherton, J.C. Helicobacter pylori persistence: Biology and disease. J. Clin. Investig. 2004, 113, 321–333. [Google Scholar] [CrossRef]
- Matsuo, Y.; Kido, Y.; Yamaoka, Y. Helicobacter pylori outer membrane protein-related pathogenesis. Toxins 2017, 9, 101. [Google Scholar] [CrossRef]
- Šterbenc, A.; Jarc, E.; Poljak, M.; Homan, M. Helicobacter pylori virulence genes. World J. Gastroenterol. 2019, 25, 4870–4884. [Google Scholar] [CrossRef]
- Yamaoka, Y. Mechanisms of disease: Helicobacter pylori virulence factors. Nat. Rev. Gastroenterol. Hepat. 2010, 7, 629–641. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Graham, D.Y. Helicobacter pylori virulence and cancer pathogenesis. Future Oncol. 2014, 10, 1487–1500. [Google Scholar] [CrossRef]
- Axon, A. Helicobacter pylori and public health. Helicobacter 2014, 19, 68–73. [Google Scholar] [CrossRef]
- Drumm, B.; Koletzko, S.; Oderda, G. Helicobacter pylori infection in children: A consensus statement. J. Pediatr. Gastroenterol. Nutr. 2000, 30, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Shin, A.; Kim, J.; Park, S. Gastric cancer epidemiology in Korea. J. Gastric Cancer 2011, 11, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Youn, H.S.; Baik, S.C.; Cho, Y.K.; Woo, H.O.; Ahn, Y.O.; Kim, K.; Cho, M.J.; Lee, W.K.; Ko, G.H.; Okada, K. Comparison of Helicobacter pylori infection between Fukuoka, Japan and Chinju, Korea. Helicobacter 1998, 3, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.S.; Kim, K.M.; Oh, Y.L.; Seo, J.K. cagA, vacA, and iceA genotypes of Helicobacter pylori in Korean children. Pediatr. Int. 2008, 50, 628–631. [Google Scholar] [CrossRef]
- Homan, M.; Šterbenc, A.; Kocjan, B.J.; Luzar, B.; Zidar, N.; Orel, R.; Poljak, M. Prevalence of the Helicobacter pylori babA2 gene and correlation with the degree of gastritis in infected Slovenian children. Anton. Leeuw. 2014, 106, 637–645. [Google Scholar] [CrossRef]
- Kalali, B.; Mejías Luque, R.; Javaheri, A.; Gerhard, M. H. pylori virulence factors: Influence on immune system and pathology. Mediat. Inflamm. 2014, 2014, 426309. [Google Scholar] [CrossRef] [PubMed]
- Khanna, B.; Cutler, A.; Israel, N.R.; Perry, M.; Lastovica, A.; Fields, P.I.; Gold, B.D. Use caution with serologic testing for Helicobacter pylori infection in children. J. Infect. Dis. 1998, 178, 460–465. [Google Scholar] [CrossRef][Green Version]
- Kim, N.; Park, R.Y.; Cho, S.I.; Lim, S.H.; Lee, K.H.; Lee, W.; Kang, H.M.; Lee, H.S.; Jung, H.C.; Song, I.S. Helicobacter pylori infection and development of gastric cancer in Korea: Long-term follow-up. J. Clin. Gastroenterol. 2008, 42, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Shiota, S.; Suzuki, R.; Yamaoka, Y. The significance of virulence factors in Helicobacter pylori. J. Dig. Dis. 2013, 14, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.Y.; Sheu, B.S.; Wu, J.J. Helicobacter pylori infection: An overview of bacterial virulence factors and pathogenesis. Biomed. J. 2016, 39, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Alm, R.A.; Trust, T.J. Analysis of the genetic diversity of Helicobacter pylori: The tale of two genomes. J. Mol. Med. 1999, 77, 834–846. [Google Scholar] [CrossRef]
- Lee, D.H.; Ha, J.H.; Shin, J.I.; Kim, K.M.; Choi, J.G.; Park, S.; Park, J.S.; Seo, J.H.; Park, J.S.; Shin, M.K.; et al. Increased Risk of Severe Gastric Symptoms by Virulence Factors vacAs1c, alpA, babA2, and hopZ in Helicobacter pylori Infection. J. Microbiol. Biotechnol. 2021, 31, 368–379. [Google Scholar] [CrossRef]
- Skene, C.; Young, A.; Every, A.; Sutton, P. Helicobacter pylori flagella: Antigenic profile and protective immunity. FEMS Immunol. Med. Microbiol. 2007, 50, 249–256. [Google Scholar] [CrossRef]
- Gholi, M.K.; Kalali, B.; Formichella, L.; Göttner, G.; Shamsipour, F.; hassan Zarnani, A.; Hosseini, M.; Busch, D.H.; Shirazi, M.H.; Gerhard, M. Helicobacter pylori FliD protein is a highly sensitive and specific marker for serologic diagnosis of H. pylori infection. Int. J. Med. Microbiol. 2013, 303, 618–623. [Google Scholar] [CrossRef]
- Baik, S.C.; Youn, H.S.; Chung, M.H.; Lee, W.K.; Cho, M.J.; Ko, G.H.; Park, C.K.; Kasai, H.; Rhee, K.H. Increased oxidative DNA damage in Helicobacter pylori-infected human gastric mucosa. Cancer Res. 1996, 56, 1279–1282. [Google Scholar]
- Song, G.Y.; Chang, M.W. Antibiotic susceptibility of Helicobacter pylori and the combination effect of antibiotics on the antibiotic-resistant H. pylori strains. J. Korean Soc. Microbiol. 1999, 34, 543–554. Available online: https://www.koreascience.or.kr/article/JAKO199917153873117.pdf (accessed on 8 December 2022).
- Monstein, H.J.; Nikpour Badr, S.; Jonasson, J. Rapid molecular identification and subtyping of Helicobacter pylori by pyrosequencing of the 16S rDNA variable V1 and V3 regions. FEMS Microbiol. Lett. 2001, 199, 103–107. [Google Scholar] [CrossRef]
- Seo, J.H.; Park, J.S.; Yeom, J.S.; Lim, J.Y.; Park, C.H.; Woo, H.O.; Baik, S.C.; Lee, W.K.; Cho, M.J.; Rhee, K.H. Correlation between positive rate and number of biopsy samples on urease test in childhood Helicobacter pylori infection. J. Korean Med. Sci. 2014, 29, 106–109. [Google Scholar] [CrossRef]
- Shin, M.K.; Jun, J.S.; Kwon, S.W.; Lee, D.H.; Ha, J.H.; Park, J.S.; Kang, H.L.; Baik, S.C.; Park, J.S.; Seo, J.H.; et al. Characterizing antigenic determinants in Helicobacter pylori CagA capable of detecting serum antibodies in children. Pathog. Dis. 2017, 75, ftx103. [Google Scholar] [CrossRef]
- Shin, M.K.; Jun, J.S.; Kwon, S.W.; Lee, D.H.; Ha, J.H.; Park, J.S.; Song, D.H.; Jung, M.; Kang, H.L.; Baik, S.C. Characterization of Specific IgA Response to Antigenic Determinants of Helicobacter pylori Urease Encoded by ureA and ureB in Children. J. Bacteriol. Virol. 2018, 48, 14–22. [Google Scholar] [CrossRef][Green Version]
- Baik, S.C.; Kim, K.M.; Song, S.M.; Kim, D.S.; Jun, J.S.; Lee, S.G.; Song, J.Y.; Park, J.U.; Kang, H.L.; Lee, W.K.; et al. Proteomic analysis of the sarcosine-insoluble outer membrane fraction of Helicobacter pylori strain 26695. J. Bacteriol. 2004, 186, 949–955. [Google Scholar] [CrossRef]
- Long, M.; Luo, J.; Li, Y.; Zeng, F.Y.; Li, M. Detection and evaluation of antibodies against neutrophil-activating protein of Helicobacter pylori in patients with gastric cancer. World J. Gastroenterol. 2009, 15, 2381–2388. [Google Scholar] [CrossRef]
- Maeda, S.; Kanai, F.; Ogura, K.; Yoshida, H.; Ikenoue, T.; Takahashi, M.; Kawabe, T.; Shiratori, Y.; Omata, M. High seropositivity of anti-CagA antibody in Helicobacter pylori-infected patients irrelevant to peptic ulcers and normal mucosa in Japan. Dig. Dis. Sci. 1997, 42, 1841–1847. [Google Scholar] [CrossRef]
- Tanaka, A.; Kamada, T.; Yokota, K.; Shiotani, A.; Hata, J.; Oguma, K.; Haruma, K. Helicobacter pylori heat shock protein 60 antibodies are associated with gastric cancer. Pathol. Res. Pract. 2009, 205, 690–694. [Google Scholar] [CrossRef]
- Tian, W.; Jia, Y.; Yuan, K.; Huang, L.; Nadolny, C.; Dong, X.; Ren, X.; Liu, J. Serum Antibody Against Helicobacter pylori FlaA and Risk of Gastric Cancer. Helicobacter 2014, 19, 9–16. [Google Scholar] [CrossRef]
- Fock, K.M.; Ang, T.L. Epidemiology of Helicobacter pylori infection and gastric cancer in Asia. J. Gastroenterol. Hepatol. 2010, 25, 479–486. [Google Scholar] [CrossRef]
- Kim, S.Y.; Woo, C.W.; Lee, Y.M.; Son, B.R.; Kim, J.W.; Chae, H.B.; Youn, S.J.; Park, S.M. Genotyping CagA, VacA subtype, IceA1, and BabA of Helicobacter pylori isolates from Korean patients, and their association with gastroduodenal diseases. J. Korean Med. Sci. 2001, 16, 579–584. [Google Scholar] [CrossRef]
- McClain, M.S.; Beckett, A.C.; Cover, T.L. Helicobacter pylori vacuolating toxin and gastric cancer. Toxins 2017, 9, 316. [Google Scholar] [CrossRef]
- Yakoob, J.; Abbas, Z.; Khan, R.; Salim, S.A.; Abrar, A.; Awan, S.; Ahmad, Z. Helicobacter pylori: Correlation of the virulence marker iceA allele with clinical outcome in a high prevalence area. Br. J. Biomed. Sci. 2015, 72, 67–73. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Kodama, T.; Gutierrez, O.; Kim, J.G.; Kashima, K.; Graham, D.Y. Relationship between Helicobacter pylori iceA, cagA, and vacA status and clinical outcome: Studies in four different countries. J. Clin. Microbiol. 1999, 37, 2274–2279. [Google Scholar] [CrossRef]
- Ando, T.; Wassenaar, T.M.; Peek, R.M., Jr.; Aras, R.A.; Tschumi, A.I.; van Doorn, L.J.; Kusugami, K.; Blaser, M.J. A Helicobacter pylori restriction endonuclease-replacing gene, hrgA, is associated with gastric cancer in Asian strains. Cancer Res. 2002, 62, 2385–2389. [Google Scholar]
- Fujimoto, S.; Ojo, O.O.; Arnqvist, A.; Wu, J.Y.; Odenbreit, S.; Haas, R.; Graham, D.Y.; Yamaoka, Y. Helicobacter pylori BabA expression, gastric mucosal injury, and clinical outcome. Clin. Gastroenterol. Hepatol. 2007, 5, 49–58. [Google Scholar] [CrossRef]
- Kennemann, L.; Brenneke, B.; Andres, S.; Engstrand, L.; Meyer, T.F.; Aebischer, T.; Josenhans, C.; Suerbaum, S. In vivo sequence variation in HopZ, a phase-variable outer membrane protein of Helicobacter pylori. Infect. Immun. 2012, 80, 4364–4373. [Google Scholar] [CrossRef]
- Miller, E.F.; Maier, R.J. Ammonium metabolism enzymes aid Helicobacter pylori acid resistance. J. Bacteriol. 2014, 196, 3074–3081. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.T.; Allen, L.A.H. Role of urease in megasome formation and Helicobacter pylori survival in macrophages. J. Leukoc. Biol. 2006, 79, 1214–1225. [Google Scholar] [CrossRef]
- Akada, J.K.; Shirai, M.; Takeuchi, H.; Tsuda, M.; Nakazawa, T. Identification of the urease operon in Helicobacter pylori and its control by mRNA decay in response to pH. Mol. Microbiol. 2000, 36, 1071–1084. [Google Scholar] [CrossRef]
- De la Cruz, M.A.; Ares, M.A.; Von Bargen, K.; Panunzi, L.G.; Martínez-Cruz, J.; Valdez Salazar, H.A.; Jiménez Galicia, C.; Torres, J. Gene expression profiling of transcription factors of Helicobacter pylori under different environmental conditions. Front. Microbiol. 2017, 8, 615. [Google Scholar] [CrossRef] [PubMed]
- Carroll, B.L.; Liu, J. Structural conservation and adaptation of the bacterial flagella motor. Biomolecules 2020, 10, 1492. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Liang, S.H.; Mao, Y.F.; Li, L.W.; Li, S.P. Construction of expression systems for flaA and flaB genes of Helicobacter pylori and determination of immunoreactivity and antigenicity of recombinant proteins. World J. Gastroenterol. 2003, 9, 2240–2250. [Google Scholar] [CrossRef]
- Hjelm, B. Epitope Mapping of Antibodies towards Human Protein Targets; KTH Royal Institute of Technology: Stockholm, Sweden, 2011. [Google Scholar]
- Ahmad, T.A.; Eweida, A.E.; Sheweita, S.A. B-cell epitope mapping for the design of vaccines and effective diagnostics. Trials Vaccinol. 2016, 5, 71–83. [Google Scholar] [CrossRef]
- Allemani, C.; Matsuda, T.; Di Carlo, V.; Harewood, R.; Matz, M.; Nikšić, M.; Bonaventure, A.; Valkov, M.; Johnson, C.J.; Estève, J. Global surveillance of trends in cancer survival 2000–14 (CONCORD-3): Analysis of individual records for 37 513 025 patients diagnosed with one of 18 cancers from 322 population-based registries in 71 countries. Lancet 2018, 391, 1023–1075. [Google Scholar] [CrossRef]
- Ito, Y.; Miyashiro, I.; Ishikawa, T.; Akazawa, K.; Fukui, K.; Katai, H.; Nunobe, S.; Oda, I.; Isobe, Y.; Tsujitani, S. Determinant factors on differences in survival for gastric cancer between the United States and Japan using nationwide databases. J. Epidemiol. 2021, 31, 241–248. [Google Scholar] [CrossRef]
- Ansari, S.; Yamaoka, Y. Helicobacter pylori Infection, Its Laboratory Diagnosis, and Antimicrobial Resistance: A Perspective of Clinical Relevance. Clin. Microbiol. Rev. 2022, 35, e00258-21. [Google Scholar] [CrossRef]
- Savarino, V.; Mela, G.S.; Zentilin, P.; Bisso, G.; Pivari, M.; Mansi, C.; Mele, M.R.; Bilardi, C.; Vigneri, S.; Celle, G. Comparison of isotope ratio mass spectrometry and nondispersive isotope-selective infrared spectroscopy for 13C-urea breath test. Am. J. Gastroenterol. 1999, 94, 1203–1208. [Google Scholar] [CrossRef]
- Li, Z.X.; Huang, L.L.; Liu, C.; Formichella, L.; Zhang, Y.; Wang, Y.M.; Zhang, L.; Ma, J.L.; Liu, W.D.; Ulm, K.; et al. Cut-off optimization for 13C-urea breath test in a community-based trial by mathematic, histology and serology approach. Sci. Rep. 2017, 7, 2072. [Google Scholar] [CrossRef]
- Yang, H.R.; Seo, J.K. Diagnostic accuracy of the 13C-urea breath test in children: Adjustment of the cut-off value according to age. J. Gastroenterol. Hepatol. 2005, 20, 264–269. [Google Scholar] [CrossRef]
- Cardinali, L.d.C.C.; Rocha, G.A.; Rocha, A.M.C.; de Moura, S.B.; de Figueiredo Soares, T.; Esteves, A.M.B.; Nogueira, A.M.M.F.; Cabral, M.M.D.A.; de Carvalho, A.S.T.; Bitencourt, P. Evaluation of [13C] urea breath test and Helicobacter pylori stool antigen test for diagnosis of H. pylori infection in children from a developing country. J. Clin. Microbiol. 2003, 41, 3334–3335. [Google Scholar] [CrossRef]
- Kato, S.; Ozawa, K.; Okuda, M.; Fujisawa, T.; Kagimoto, S.; Konno, M.; Maisawa, S.; Iinuma, K. Accuracy of the stool antigen test for the diagnosis of childhood Helicobacter pylori infection: A multicenter Japanese study. Am. J. Gastroenterol. 2003, 98, 296–300. [Google Scholar] [CrossRef]
- Manes, G.; Balzano, A.; Iaquinto, G.; Ricci, C.; Piccirillo, M.; Giardullo, N.; Todisco, A.; Lioniello, M.; Vaira, D. Accuracy of the stool antigen test in the diagnosis of Helicobacter pylori infection before treatment and in patients on omeprazole therapy. Aliment. Pharmacol. Ther. 2001, 15, 73–79. [Google Scholar] [CrossRef]
- Oderda, G.; Rapa, A.; Ronchi, B.; Lerro, P.; Pastore, M.; Staiano, A.; De’Angelis, G.; Strisciuglio, P. Detection of Helicobacter pylori in stool specimens by non-invasive antigen enzyme immunoassay in children: Multicentre Italian study. BMJ 2000, 320, 347–348. [Google Scholar] [CrossRef][Green Version]
- Garza González, E.; Perez Perez, G.I.; Maldonado Garza, H.J.; Bosques Padilla, F.J. A review of Helicobacter pylori diagnosis, treatment, and methods to detect eradication. World J. Gastroenterol. 2014, 20, 1438. [Google Scholar] [CrossRef]
- Gisbert, J.; Hermida, C.; Cabrera, M.; Pajares, J. Antibodies against Helicobacter pylori in saliva. Study of their validity versus breath test and its agreement with serology. Aten. Primaria 2000, 25, 390–394. [Google Scholar]
- Herbrink, P.; Van Doorn, L. Serological methods for diagnosis of Helicobacter pylori infection and monitoring of eradication therapy. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 164–173. [Google Scholar] [CrossRef]
- Urita, Y.; Hike, K.; Torii, N.; Kikuchi, Y.; Kurakata, H.; Kanda, E.; Sasajima, M.; Miki, K. Comparison of serum IgA and IgG antibodies for detecting Helicobacter pylori infection. Intern. Med. 2004, 43, 548–552. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Patel, S.K.; Pratap, C.B.; Jain, A.K.; Gulati, A.K.; Nath, G. Diagnosis of Helicobacter pylori: What should be the gold standard? World J. Gastroenterol. 2014, 20, 12847. [Google Scholar] [CrossRef] [PubMed]
- Darma, A.; Nugroho, B.S.T.; Yoanna, V.; Sulistyani, I.; Athiyyah, A.F.; Ranuh, R.G.; Sudarmo, S.M. Comparison of Helicobacter pylori stool antigen, salivary IgG, serum IgG, and serum IgM as diagnostic markers of H. pylori infection in children. Iran. J. Microbiol. 2019, 11, 206. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6711868/ (accessed on 8 December 2022). [CrossRef]
- Choi, K.S.; Jun, J.K.; Park, E.C.; Park, S.; Jung, K.W.; Han, M.A.; Choi, I.J.; Lee, H.Y. Performance of different gastric cancer screening methods in Korea: A population-based study. PLoS ONE 2012, 7, e50041. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).