


Molecular Cloning and Characterization of a Fasciola gigantica Nuclear Receptor Subfamily 1 (FgNR1)

 ,
,  , ,
, , 

Abstract
1. Introduction
2. Materials and Methods
2.1. Parasites
2.2. Bioinformatic Analysis
2.3. Total RNA Isolation and Synthesis of FgNR1 Complementary DNA
2.4. Molecular Cloning of FgNR1
2.5. Expression and Purification of Recombinant FgNR1 (rFgNR1) and Recombinant Thioredoxin (rTrx) Fusion Proteins
2.6. Production of Polyclonal Antibodies against rFgNR1 and rTdx
2.7. Preparation of Parasite Protein Extracts
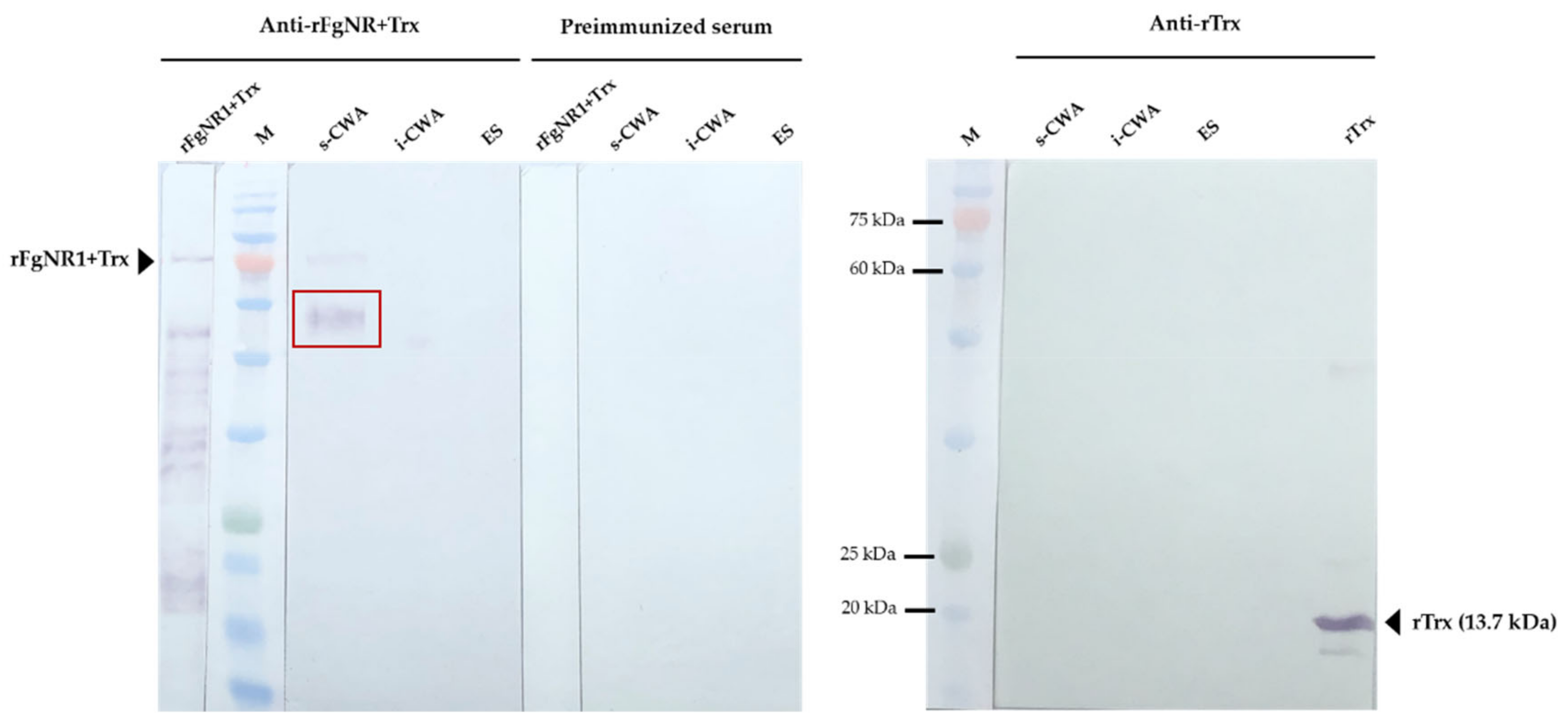
2.8. Western Analysis
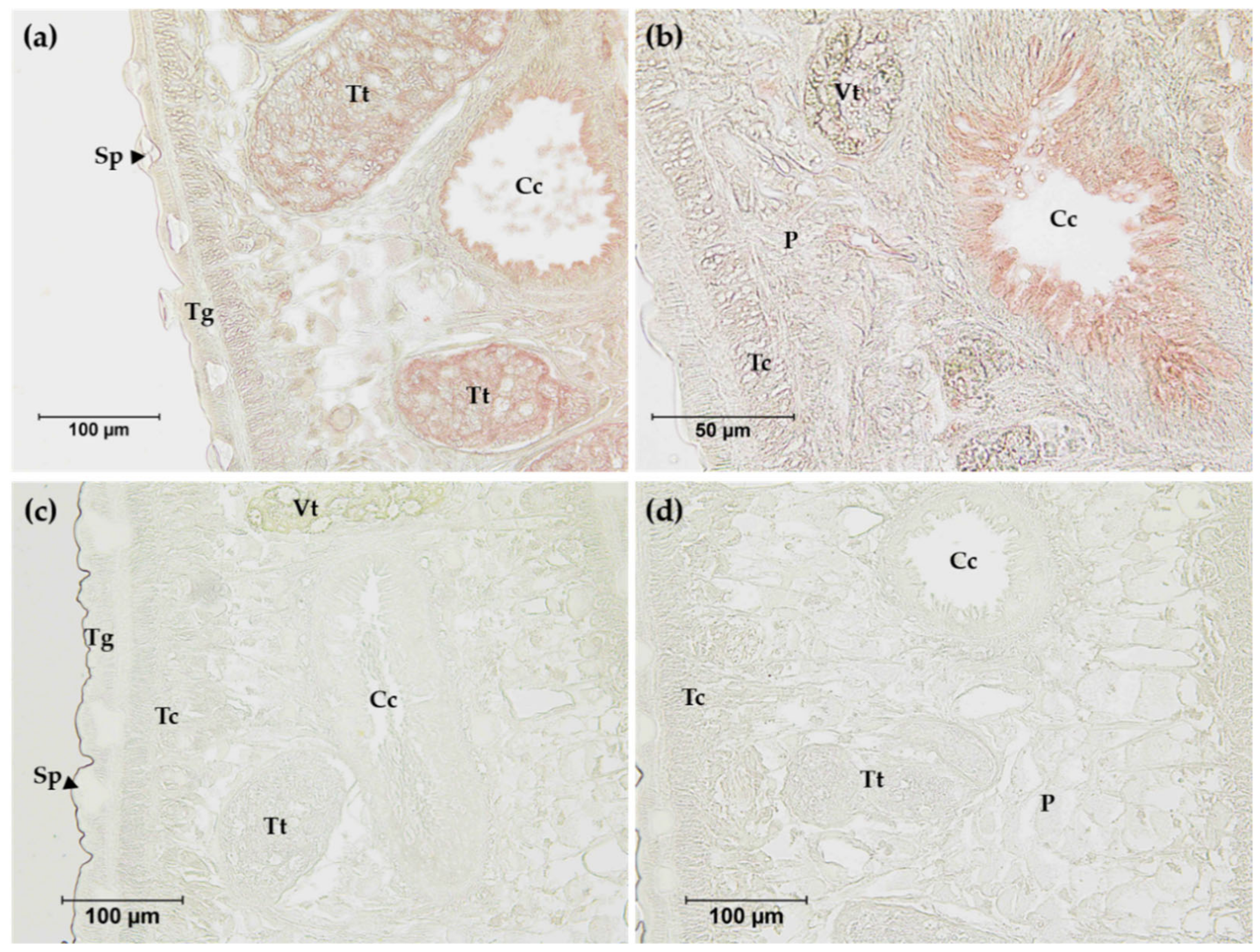
2.9. Immunolocalization of FgNR1
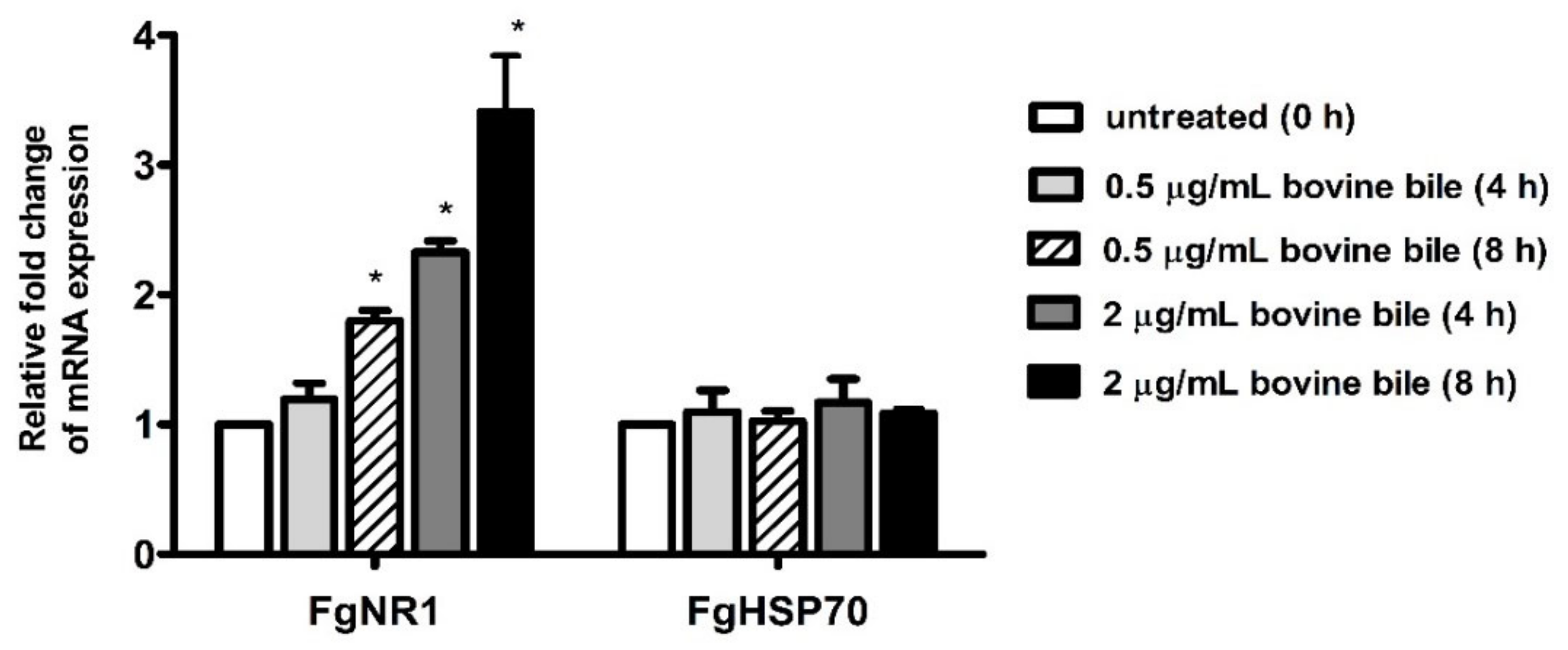
2.10. Parasite Culture with Bile
2.11. Real-Time RT-PCR
3. Results
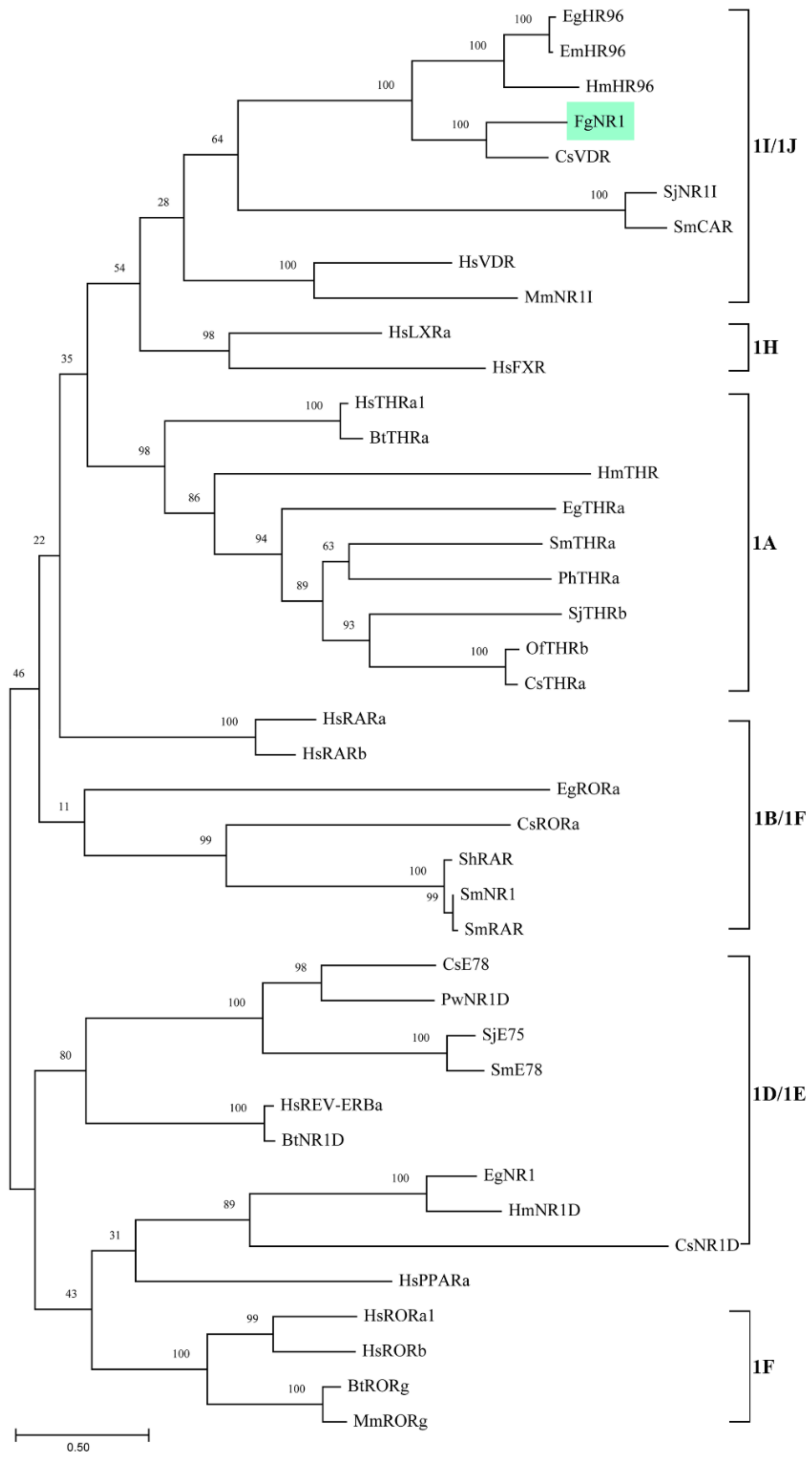
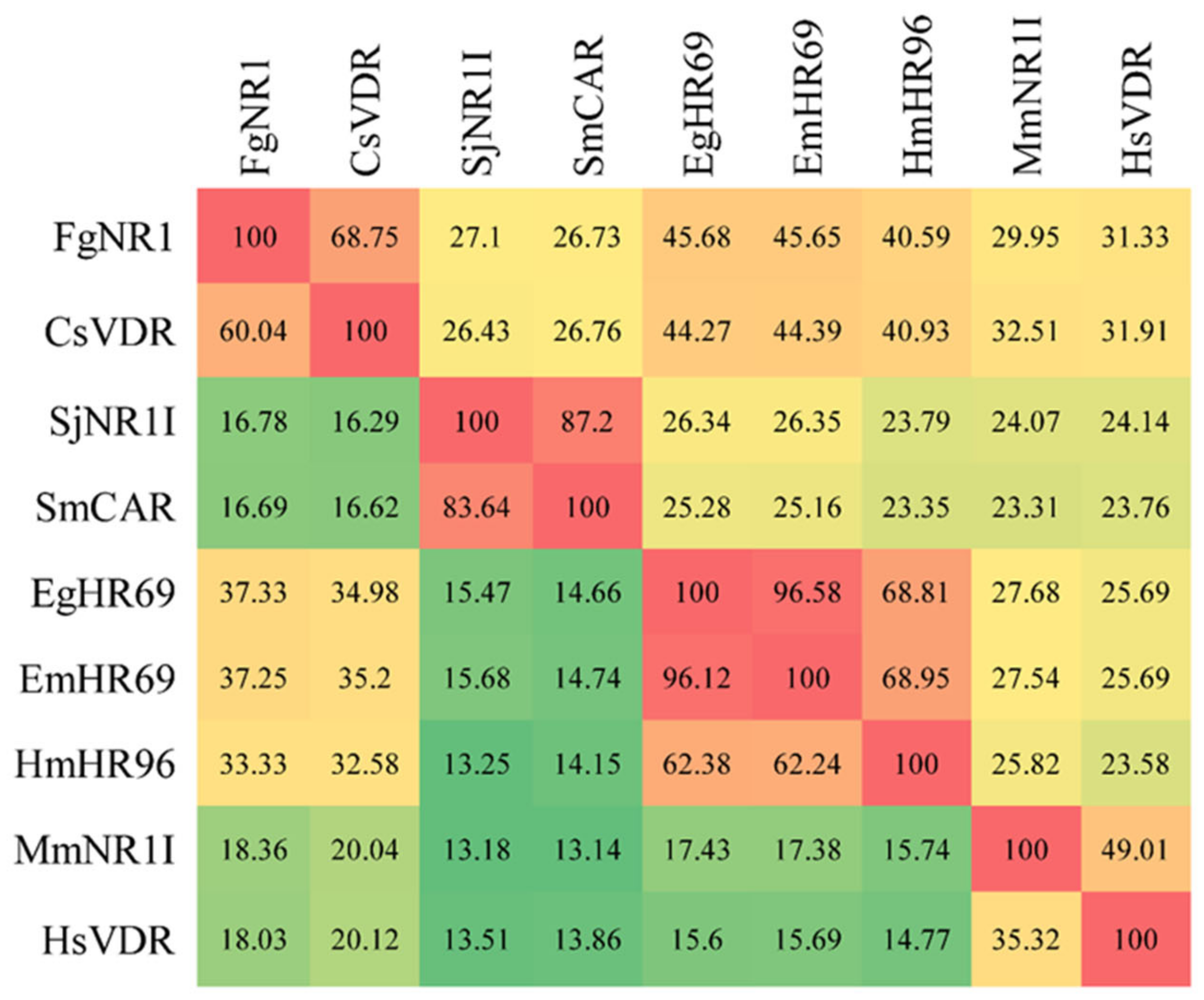
3.1. Molecular Properties of FgNR1
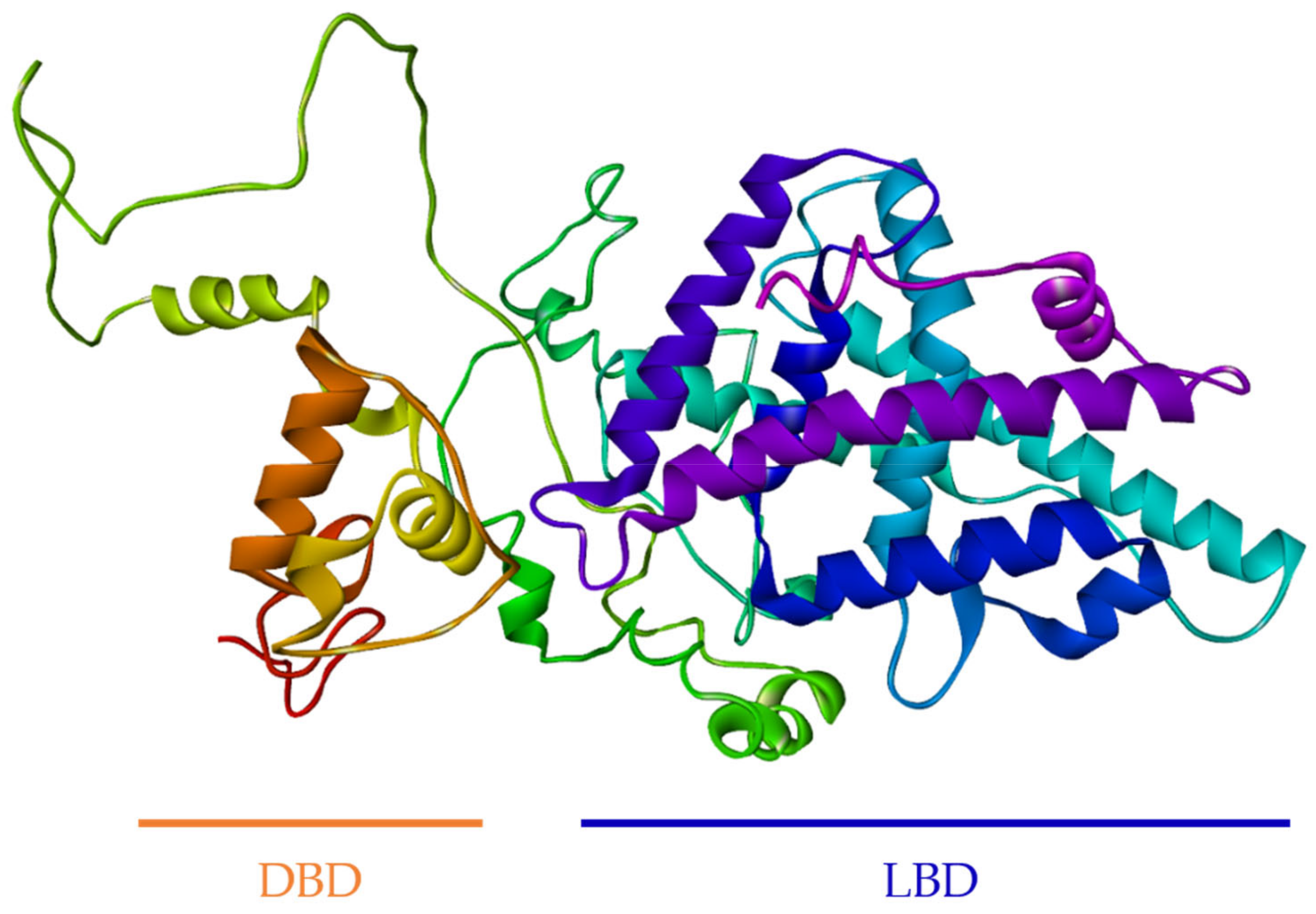
3.2. Computational Modeling of FgNR1
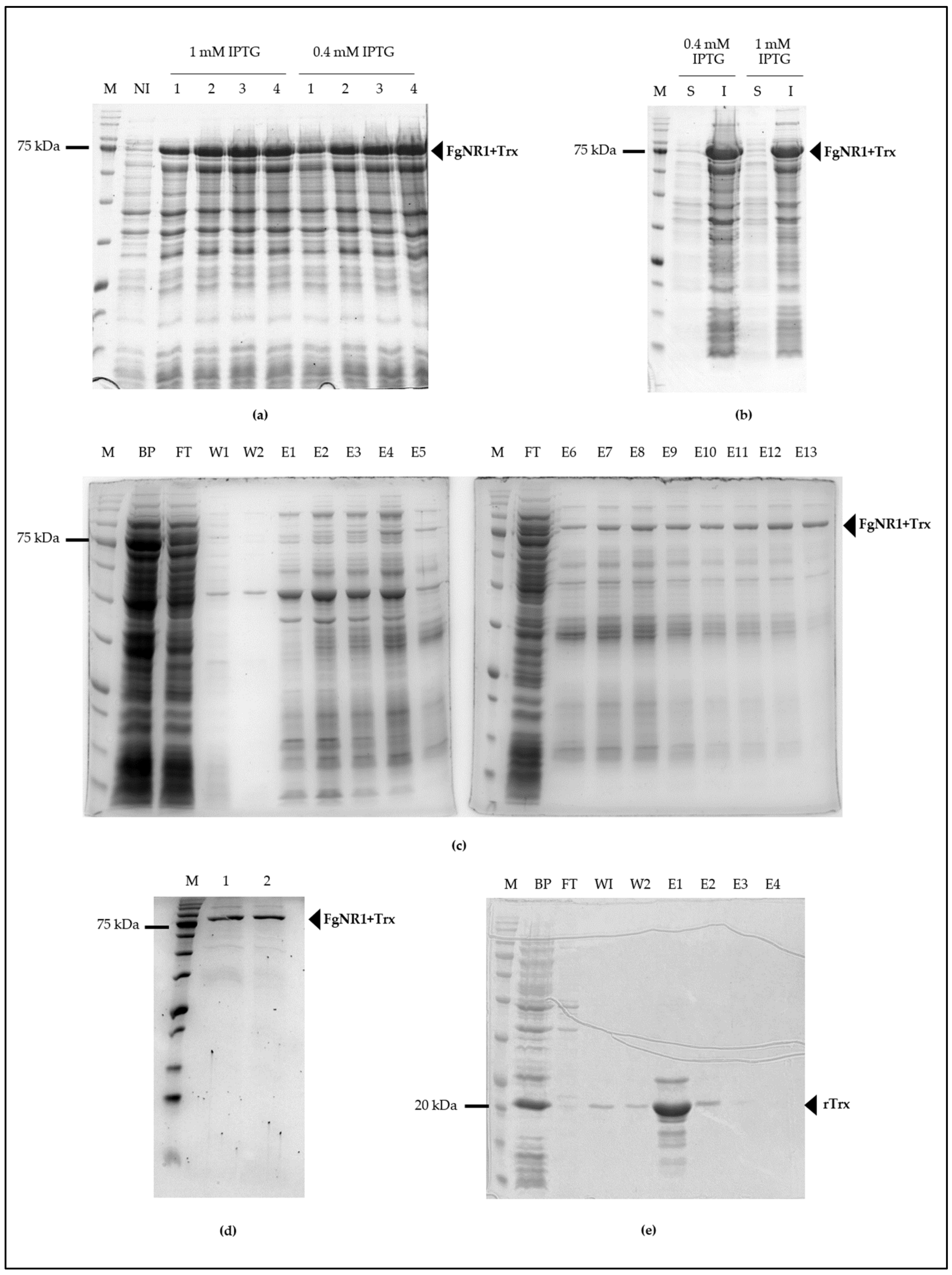
3.3. Molecular Cloning and Production of Recombinant FgNR1 (rFgNR1)
3.4. Detection of Native FgNR1 in Parasite Tissue and Extracts
3.5. Bile Stimulated FgNR1 mRNA Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Species * | Protein Name | GenBank/Accession No. |
|---|---|---|---|
| FgNR1 | F. gigantiga | Nuclear receptor subfamily 1 | TPP57711.1 |
| HsTHRa1 | H. sapiens | Thyroid hormone receptor-α1 | CAA38749.1 |
| HsRARa | H. sapiens | Retinoic acid receptor-α | CAA29829.1 |
| HsRARb | H. sapiens | Retinoic acid receptor-β | CAA30262.1 |
| HsPPARa | H. sapiens | Peroxisome proliferator-activated receptor-α | AAA36469.1 |
| HsREV-ERBa | H. sapiens | Reverse-Erb-α | AAA52335.1 |
| HsRORa1 | H. sapiens | Retinoic acid-related orphan-α1 | AAA62658.1 |
| HsRORb | H. sapiens | Retinoic acid-related orphan-β | CAA69929.1 |
| HsLXRa | H. sapiens | Liver X receptor-α | AAA85856.1 |
| HsFXR | H. sapiens | Farnesoid X receptor | AAB08107.1 |
| HsVDR | H. sapiens | Vitamin D receptor | AAA61273.1 |
| BtNR1D | B. taurus | Nuclear receptor subfamily 1D | NP_001071568.1 |
| BtRORg | B. taurus | Retinoic acid-related orphan-γ | NP_001076920.1 |
| BtTHRa | B. taurus | Thyroid hormone receptor-α | NP_001039794.1 |
| MmNR1I | M. musculus | Nuclear receptor subfamily 1I | NP_035066.1 |
| MmRORg | M. musculus | Retinoic acid-related orphan-γ | NP_035411.2 |
| OfTHRb | O. felineus | Thyroid hormone receptor-β | AFM38060.1 |
| CsTHRa | C. sinensis | Thyroid hormone receptor-α | KAG5441315.1 |
| CsE78 | C. sinensis | Ecdysone-induced protein 78C | KAG5445165.1 |
| CsNR1D | C. sinensis | Nuclear receptor subfamily 1D | GAA57457.1 |
| CsRORa | C. sinensis | Retinoic acid-related orphan-α | GAA58146.1 |
| CsVDR | C. sinensis | Vitamin D receptor | KAG5451015.1 |
| ShRAR | S. haematobium | Retinoic acid receptor | XP_035588303.1 |
| SjNR1I | S. japonicum | Nuclear receptor subfamily 1I | TNN17786.1 |
| SjE75 | S. japonicum | Nuclear hormone receptor E75 | KAH8876702.1 |
| SjTHRb | S. japonicum | Thyroid hormone receptor-β | AFP95236.1 |
| SmNR1 | S. mansoni | Nuclear receptor subfamily 1 | AAR29357.1 |
| SmTHRa | S. mansoni | Thyroid hormone receptor-α | AAR29358.1 |
| SmE78 | S. mansoni | Ecdysone-induced protein 78 | AAR30507.2 |
| SmRAR | S. mansoni | Retinoic acid receptor | XP_018649430.1 |
| SmCAR | S. mansoni | Constitutive androstane receptor | AAV80235.1 |
| PwNR1D | P. westermani | Nuclear receptor subfamily 1D | KAA3676691.1 |
| PhTHRa | P. heterotremus | Thyroid hormone receptor-α | KAF5398468.1 |
| EgRORa | E. granulosus | Retinoic acid-related orphan-α | KAH9284012.1 |
| EgHR96 | E. granulosus | Nuclear hormone receptor HR96 | CDS22803.1 |
| EgTHRa | E. granulosus | Thyroid hormone receptor-α | CDS24335.1 |
| EgNR1 | E. granulosus | Nuclear receptor subfamily 1 | ART84255.1 |
| EmHR96 | E. multilocularis | Nuclear hormone receptor HR96 | CDS36018.1 |
| HmNR1D | H. microstoma | Nuclear receptor subfamily 1D | CDS29812.1 |
| HmHR96 | H. microstoma | Nuclear hormone receptor HR96 | CDS28331.1 |
| HmTHR | H. microstoma | Thyroid hormone receptor | CDS30783.1 |
References
- Andrews, N. The Life Cycle of Fasciola hepatica, 3rd ed.; Dalton, J.P., Ed.; CABI Publishing: Oxon, UK, 1999. [Google Scholar]
- Mas-Coma, S.; Agramunt, V.H.; Valero, M.A. Neurological and ocular fascioliasis in humans. Adv. Parasitol. 2014, 84, 27–149. [Google Scholar] [CrossRef] [PubMed]
- Torgerson, P.R.; Devleesschauwer, B.; Praet, N.; Speybroeck, N.; Willingham, A.L.; Kasuga, F.; Rokni, M.B.; Zhou, X.N.; Fèvre, E.M.; Sripa, B.; et al. World Health Organization Estimates of the Global and Regional Disease Burden of 11 Foodborne Parasitic Diseases, 2010: A Data Synthesis. PLoS Med. 2015, 12, e1001920. [Google Scholar] [CrossRef] [PubMed]
- Lalor, R.; Cwiklinski, K.; Calvani, N.E.D.; Dorey, A.; Hamon, S.; Corrales, J.L.; Dalton, J.P.; De Marco Verissimo, C. Pathogenicity and virulence of the liver flukes Fasciola hepatica and Fasciola Gigantica that cause the zoonosis Fasciolosis. Virulence 2021, 12, 2839–2867. [Google Scholar] [CrossRef] [PubMed]
- Fairweather, I.; Brennan, G.P.; Hanna, R.E.B.; Robinson, M.W.; Skuce, P.J. Drug resistance in liver flukes. Int. J. Parasitol. Drugs Drug Resist. 2020, 12, 39–59. [Google Scholar] [CrossRef] [PubMed]
- Savage, J.; Meaney, M.; Brennan, G.P.; Hoey, E.; Trudgett, A.; Fairweather, I. Effect of the P-glycoprotein inhibitor, R(+)-verapamil on the drug susceptibility of a triclabendazole-resistant isolate of Fasciola hepatica. Vet. Parasitol. 2013, 195, 72–86. [Google Scholar] [CrossRef]
- Novobilský, A.; Averpil, H.B.; Höglund, J. The field evaluation of albendazole and triclabendazole efficacy against Fasciola hepatica by coproantigen ELISA in naturally infected sheep. Vet. Parasitol. 2012, 190, 272–276. [Google Scholar] [CrossRef]
- Ortiz, P.; Scarcella, S.; Cerna, C.; Rosales, C.; Cabrera, M.; Guzmán, M.; Lamenza, P.; Solana, H. Resistance of Fasciola hepatica against Triclabendazole in cattle in Cajamarca (Peru): A clinical trial and an in vivo efficacy test in sheep. Vet. Parasitol. 2013, 195, 118–121. [Google Scholar] [CrossRef]
- Mamani, L.W.; Condori, Q.R. Anthelminthic resistance (Fasciola hepatica) in sheep against albendazole and triclabendazole, La Paz-Bolivia. Rev. Investig. Vet. Perú (RIVEP) 2009, 20, 254–262. [Google Scholar]
- Chávez, A.; Sánchez, L.; Arana, C.; Suárez, F. Resistance to anthelmintics and prevalence of bovine fasciolosis in dairy farms in Jauja, Peru. Rev. Investig. Vet. Perú (RIVEP) 2012, 23, 90–97. [Google Scholar]
- Dawson, P.A.; Karpen, S.J. Intestinal transport and metabolism of bile acids. J. Lipid Res. 2015, 56, 1085–1099. [Google Scholar] [CrossRef]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schütz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P.; et al. The nuclear receptor superfamily: The second decade. Cell 1995, 83, 835–839. [Google Scholar] [CrossRef]
- Robinson-Rechavi, M.; Garcia, H.E.; Laudet, V. The nuclear receptor superfamily. J. Cell Sci. 2003, 116, 585–586. [Google Scholar] [CrossRef]
- Crowder, M.K.; Seacrist, C.D.; Blind, R.D. Phospholipid regulation of the nuclear receptor superfamily. Adv. Biol. Regul. 2017, 63, 6–14. [Google Scholar] [CrossRef]
- Weikum, E.R.; Liu, X.; Ortlund, E.A. The nuclear receptor superfamily: A structural perspective. Protein Sci. 2018, 27, 1876–1892. [Google Scholar] [CrossRef]
- Bain, D.L.; Heneghan, A.F.; Connaghan-Jones, K.D.; Miura, M.T. Nuclear receptor structure: Implications for function. Annu. Rev. Physiol. 2007, 69, 201–220. [Google Scholar] [CrossRef]
- Veras Ribeiro Filho, H.; Tambones, I.L.; Mariano Gonçalves Dias, M.; Bernardi Videira, N.; Bruder, M.; Amorim Amato, A.; Migliorini Figueira, A.C. Modulation of nuclear receptor function: Targeting the protein-DNA interface. Mol. Cell. Endocrinol. 2019, 484, 1–14. [Google Scholar] [CrossRef]
- Schulman, I.G.; Heyman, R.A. The flip side: Identifying small molecule regulators of nuclear receptors. Chem. Biol. 2004, 11, 639–646. [Google Scholar] [CrossRef]
- Farthing, M.J.; Keusch, G.T.; Carey, M.C. Effects of bile and bile salts on growth and membrane lipid uptake by Giardia lamblia. Possible implications for pathogenesis of intestinal disease. J. Clin. Investig. 1985, 76, 1727–1732. [Google Scholar] [CrossRef]
- Halliday, C.E.; Clark, C.; Farthing, M.J. Giardia-bile salt interactions in vitro and in vivo. Trans. R. Soc. Trop. Med. Hyg. 1988, 82, 428–432. [Google Scholar] [CrossRef]
- Halliday, C.E.; Inge, P.M.; Farthing, M.J. Characterization of bile salt uptake by Giardia lamblia. Int. J. Parasitol. 1995, 25, 1089–1097. [Google Scholar] [CrossRef]
- Badr, S.G.; Pica-Mattoccia, L.; Moroni, R.; Angelico, M.; Cioli, D. Effect of bile salts on oviposition in vitro by Schistosoma mansoni. Parasitol. Res. 1999, 85, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Shaker, Y.M.; Hamdy, M.A.; Ismail, M.; Draz, H.M.; Ashour, E.; Gouda, W. Effect of host portal and peripheral sera fractions on cell proliferation of Schistosoma mansoni schistosomules. Trop. Biomed. 2011, 28, 630–637. [Google Scholar] [PubMed]
- De Mendonça, R.L.; Bouton, D.; Bertin, B.; Escriva, H.; Noël, C.; Vanacker, J.M.; Cornette, J.; Laudet, V.; Pierce, R.J. A functionally conserved member of the FTZ-F1 nuclear receptor family from Schistosoma mansoni. Eur. J. Biochem. 2002, 269, 5700–5711. [Google Scholar] [CrossRef] [PubMed]
- De Mendonça, R.L.; Escriva, H.; Bouton, D.; Zelus, D.; Vanacker, J.M.; Bonnelye, E.; Cornette, J.; Pierce, R.J.; Laudet, V. Structural and functional divergence of a nuclear receptor of the RXR family from the trematode parasite Schistosoma mansoni. Eur. J. Biochem. 2000, 267, 3208–3219. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Li, J.; Wu, J.; Wang, H.; Guo, B.; Wu, C.; Shou, X.; Yang, N.; Zhang, Z.; McManus, D.P.; et al. Cloning and characterization of an Echinococcus granulosus ecdysteroid hormone nuclear receptor HR3-like gene. Parasite 2017, 24, 36. [Google Scholar] [CrossRef]
- Alvite, G.; Riera, X.; Cancela, S.; Paulino, M.; Esteves, A. Bioinformatic analysis of a novel Echinococcus granulosus nuclear receptor with two DNA binding domains. PLoS ONE 2019, 14, e0224703. [Google Scholar] [CrossRef]
- Wu, W.; LoVerde, P.T. Identification and evolution of nuclear receptors in Platyhelminths. PLoS ONE 2021, 16, e0250750. [Google Scholar] [CrossRef]
- Wang, Z.; Stoltzfus, J.; You, Y.J.; Ranjit, N.; Tang, H.; Xie, Y.; Lok, J.B.; Mangelsdorf, D.J.; Kliewer, S.A. The nuclear receptor DAF-12 regulates nutrient metabolism and reproductive growth in nematodes. PLoS Genet. 2015, 11, e1005027. [Google Scholar] [CrossRef]
- Gerisch, B.; Rottiers, V.; Li, D.; Motola, D.L.; Cummins, C.L.; Lehrach, H.; Mangelsdorf, D.J.; Antebi, A. A bile acid-like steroid modulates Caenorhabditis elegans lifespan through nuclear receptor signaling. Proc. Natl. Acad. Sci. USA 2007, 104, 5014–5019. [Google Scholar] [CrossRef]
- Groen, A.K.; Kuipers, F. Bile acid look-alike controls life span in C. elegans. Cell Metab. 2013, 18, 151–152. [Google Scholar] [CrossRef]
- Wang, Z.; Cheong, M.C.; Tsien, J.; Deng, H.; Qin, T.; Stoltzfus, J.D.; Jaleta, T.G.; Li, X.; Lok, J.B.; Kliewer, S.A.; et al. Characterization of the endogenous DAF-12 ligand and its use as an anthelmintic agent in Strongyloides stercoralis. eLife 2021, 10, e73535. [Google Scholar] [CrossRef]
- Cheong, M.C.; Wang, Z.; Jaleta, T.G.; Li, X.; Lok, J.B.; Kliewer, S.A.; Mangelsdorf, D.J. Identification of a nuclear receptor/coactivator developmental signaling pathway in the nematode parasite Strongyloides stercoralis. Proc. Natl. Acad. Sci. USA 2021, 118, e2021864118. [Google Scholar] [CrossRef]
- Wang, Z.; Schaffer, N.E.; Kliewer, S.A.; Mangelsdorf, D.J. Nuclear receptors: Emerging drug targets for parasitic diseases. J. Clin. Investig. 2017, 127, 1165–1171. [Google Scholar] [CrossRef]
- Wu, W.; LoVerde, P.T. Nuclear hormone receptors in parasitic helminths. Mol. Cell. Endocrinol. 2011, 334, 56–66. [Google Scholar] [CrossRef]
- Martviset, P.; Grams, R. A single parathyroid hormone receptor-like member of family B1 G-protein coupled receptors in Fasciola gigantica. Southeast Asian J. Trop. Med. Public Health 2017, 48, 762–776. [Google Scholar]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef]
- Hamby, S.E.; Hirst, J.D. Prediction of glycosylation sites using random forests. BMC Bioinform. 2008, 9, 500. [Google Scholar] [CrossRef]
- Cheng, J.; Saigo, H.; Baldi, P. Large-scale prediction of disulphide bridges using kernel methods, two-dimensional recursive neural networks, and weighted graph matching. Proteins 2006, 62, 617–629. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Stothard, P. The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. Biotechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; Jabłońska, J.; Pravda, L.; Vařeková, R.S.; Thornton, J.M. PDBsum: Structural summaries of PDB entries. Protein Sci. 2018, 27, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Porollo, A.A.; Adamczak, R.; Meller, J. POLYVIEW: A flexible visualization tool for structural and functional annotations of proteins. Bioinformatics 2004, 20, 2460–2462. [Google Scholar] [CrossRef]
- Heo, L.; Park, H.; Seok, C. GalaxyRefine: Protein structure refinement driven by side-chain repacking. Nucleic Acids Res. 2013, 41, W384–W388. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y. Protein Structure and Function Prediction Using I-TASSER. Curr. Protoc. Bioinform. 2015, 52, 5.8.1–5.8.15. [Google Scholar] [CrossRef]
- Wang, J.; Youkharibache, P.; Zhang, D.; Lanczycki, C.J.; Geer, R.C.; Madej, T.; Phan, L.; Ward, M.; Lu, S.; Marchler, G.H.; et al. iCn3D, a web-based 3D viewer for sharing 1D/2D/3D representations of biomolecular structures. Bioinformatics 2020, 36, 131–135. [Google Scholar] [CrossRef]
- Wang, J.; Youkharibache, P.; Marchler-Bauer, A.; Lanczycki, C.; Zhang, D.; Lu, S.; Madej, T.; Marchler, G.H.; Cheng, T.; Chong, L.C.; et al. iCn3D: From Web-Based 3D Viewer to Structural Analysis Tool in Batch Mode. Front. Mol. Biosci. 2022, 9, 831740. [Google Scholar] [CrossRef]
- Tarasuk, M.; Vichasri Grams, S.; Viyanant, V.; Grams, R. Type I cystatin (stefin) is a major component of Fasciola gigantica excretion/secretion product. Mol. Biochem. Parasitol. 2009, 167, 60–71. [Google Scholar] [CrossRef]
- Martviset, P.; Panrit, L.; Chantree, P.; Muhamad, P.; Na-Bangchang, K. Suppression of Cholangiocarcinoma Cell Growth and Proliferation by Atractylodes lancea (Thunb) DC. through ERK-Signaling Cascade. Asian Pac. J. Cancer Prev. 2021, 22, 3633–3640. [Google Scholar] [CrossRef]
- Sever, R.; Glass, C.K. Signaling by nuclear receptors. Cold Spring Harb. Perspect. Biol. 2013, 5, a016709. [Google Scholar] [CrossRef]
- Wärnmark, A.; Treuter, E.; Wright, A.P.H.; Gustafsson, J.-A.k. Activation Functions 1 and 2 of Nuclear Receptors: Molecular Strategies for Transcriptional Activation. Mol. Endocrinol. 2003, 17, 1901–1909. [Google Scholar] [CrossRef]
- Porter, B.A.; Ortiz, M.A.; Bratslavsky, G.; Kotula, L. Structure and Function of the Nuclear Receptor Superfamily and Current Targeted Therapies of Prostate Cancer. Cancers 2019, 11, 1852. [Google Scholar] [CrossRef]
- Wu, W.; Niles, E.G.; El-Sayed, N.; Berriman, M.; LoVerde, P.T. Schistosoma mansoni (Platyhelminthes, Trematoda) nuclear receptors: Sixteen new members and a novel subfamily. Gene 2006, 366, 303–315. [Google Scholar] [CrossRef]
- Escobedo, G.; Larralde, C.; Chavarria, A.; Cerbón, M.A.; Morales-Montor, J. Molecular mechanisms involved in the differential effects of sex steroids on the reproduction and infectivity of Taenia crassiceps. J. Parasitol. 2004, 90, 1235–1244. [Google Scholar] [CrossRef]
- Pakharukova, M.Y.; Ershov, N.I.; Vorontsova, E.V.; Shilov, A.G.; Merkulova, T.I.; Mordvinov, V.A. Identification of thyroid hormone receptor homologs in the fluke Opisthorchis felineus (Platyhelminthes). Mol. Biochem. Parasitol. 2014, 194, 64–68. [Google Scholar] [CrossRef]
- Young, N.D.; Jex, A.R.; Cantacessi, C.; Hall, R.S.; Campbell, B.E.; Spithill, T.W.; Tangkawattana, S.; Tangkawattana, P.; Laha, T.; Gasser, R.B. A portrait of the transcriptome of the neglected trematode, Fasciola gigantica–biological and biotechnological implications. PLoS Negl. Trop. Dis. 2011, 5, e1004. [Google Scholar] [CrossRef]
- Luisi, B.F.; Xu, W.X.; Otwinowski, Z.; Freedman, L.P.; Yamamoto, K.R.; Sigler, P.B. Crystallographic analysis of the interaction of the glucocorticoid receptor with DNA. Nature 1991, 352, 497–505. [Google Scholar] [CrossRef]
- Zechel, C.; Shen, X.Q.; Chambon, P.; Gronemeyer, H. Dimerization interfaces formed between the DNA binding domains determine the cooperative binding of RXR/RAR and RXR/TR heterodimers to DR5 and DR4 elements. EMBO J. 1994, 13, 1414–1424. [Google Scholar] [CrossRef]
- Germain, P.; Staels, B.; Dacquet, C.; Spedding, M.; Laudet, V. Overview of nomenclature of nuclear receptors. Pharmacol. Rev. 2006, 58, 685–704. [Google Scholar] [CrossRef] [PubMed]
- Rastinejad, F.; Huang, P.; Chandra, V.; Khorasanizadeh, S. Understanding nuclear receptor form and function using structural biology. J. Mol. Endocrinol. 2013, 51, T1–T21. [Google Scholar] [CrossRef] [PubMed]
- Egea, P.F.; Mitschler, A.; Rochel, N.; Ruff, M.; Chambon, P.; Moras, D. Crystal structure of the human RXRalpha ligand-binding domain bound to its natural ligand: 9-cis retinoic acid. EMBO J. 2000, 19, 2592–2601. [Google Scholar] [CrossRef] [PubMed]
- Dowd, D.R.; MacDonald, P.N. Vitamin D Receptor. In Encyclopedia of Biological Chemistry, 2nd ed.; Lennarz, W.J., Lane, M.D., Eds.; Academic Press: Waltham, MA, USA, 2013; pp. 540–544. [Google Scholar]
- Kato, S. The function of vitamin D receptor in vitamin D action. J. Biochem. 2000, 127, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Young, N.D.; Campbell, B.E.; Hall, R.S.; Jex, A.R.; Cantacessi, C.; Laha, T.; Sohn, W.M.; Sripa, B.; Loukas, A.; Brindley, P.J.; et al. Unlocking the transcriptomes of two carcinogenic parasites, Clonorchis sinensis and Opisthorchis viverrini. PLoS Negl. Trop. Dis. 2010, 4, e719. [Google Scholar] [CrossRef]
- Förster, S.; Günthel, D.; Kiss, F.; Brehm, K. Molecular characterisation of a serum-responsive, DAF-12-like nuclear hormone receptor of the fox-tapeworm Echinococcus multilocularis. J. Cell. Biochem. 2011, 112, 1630–1642. [Google Scholar] [CrossRef]
- Wen, J.; Mercado, G.P.; Volland, A.; Doden, H.L.; Lickwar, C.R.; Crooks, T.; Kakiyama, G.; Kelly, C.; Cocchiaro, J.L.; Ridlon, J.M.; et al. Fxr signaling and microbial metabolism of bile salts in the zebrafish intestine. Sci. Adv. 2021, 7, eabg1371. [Google Scholar] [CrossRef]
- Romero, A.A.; Cobb, S.A.; Collins, J.N.R.; Kliewer, S.A.; Mangelsdorf, D.J.; Collins, J.J., 3rd. The Schistosoma mansoni nuclear receptor FTZ-F1 maintains esophageal gland function via transcriptional regulation of meg-8.3. PLoS Pathog. 2021, 17, e1010140. [Google Scholar] [CrossRef]
- Shi, W.; Wei, Z.-Y.; Elsheikha, H.M.; Zhang, F.-K.; Sheng, Z.-A.; Lu, K.-J.; Wang, D.-Y.; Huang, W.-Y.; Zhu, X.-Q. Dynamic expression of cytokine and transcription factor genes during experimental Fasciola gigantica infection in buffaloes. Parasites Vectors 2017, 10, 602. [Google Scholar] [CrossRef]
- Dimauro, I.; Pearson, T.; Caporossi, D.; Jackson, M.J. A simple protocol for the subcellular fractionation of skeletal muscle cells and tissue. BMC Res. Notes 2012, 5, 513. [Google Scholar] [CrossRef]
- Cox, B.; Emili, A. Tissue subcellular fractionation and protein extraction for use in mass-spectrometry-based proteomics. Nat. Protoc. 2006, 1, 1872–1878. [Google Scholar] [CrossRef]
- Dar, J.S.; Ganai, B.A.; Shahardar, R.A.; Zargar, U.R. Molecular Characterization and Immunodiagnostic Potential of Various Antigenic Proteins of Fasciola Gigantica Species Isolated from Sheep of North West Himalayan Region. Helminthologia 2019, 56, 93–107. [Google Scholar] [CrossRef]
- Reschly, E.J.; Ai, N.; Ekins, S.; Welsh, W.J.; Hagey, L.R.; Hofmann, A.F.; Krasowski, M.D. Evolution of the bile salt nuclear receptor FXR in vertebrates. J. Lipid Res. 2008, 49, 1577–1587. [Google Scholar] [CrossRef]
- Li, T.; Chiang, J.Y. Nuclear receptors in bile acid metabolism. Drug Metab. Rev. 2013, 45, 145–155. [Google Scholar] [CrossRef]
- Schmidt, D.R.; Kliewer, S.A.; Mangelsdorf, D.J. Chapter 43—The role of the vitamin D receptor in bile acid homeostasis. In Vitamin D, 3rd ed.; Feldman, D., Pike, J.W., Adams, J.S., Eds.; Academic Press: San Diego, CA, USA, 2011; pp. 763–767. [Google Scholar]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martviset, P.; Chantree, P.; Chaimon, S.; Torungkitmangmi, N.; Prathaphan, P.; Ruangtong, J.; Sornchuer, P.; Thongsepee, N.; Sangpairoj, K.; Adisakwattana, P. Molecular Cloning and Characterization of a Fasciola gigantica Nuclear Receptor Subfamily 1 (FgNR1). Pathogens 2022, 11, 1458. https://doi.org/10.3390/pathogens11121458
Martviset P, Chantree P, Chaimon S, Torungkitmangmi N, Prathaphan P, Ruangtong J, Sornchuer P, Thongsepee N, Sangpairoj K, Adisakwattana P. Molecular Cloning and Characterization of a Fasciola gigantica Nuclear Receptor Subfamily 1 (FgNR1). Pathogens. 2022; 11(12):1458. https://doi.org/10.3390/pathogens11121458
Chicago/Turabian StyleMartviset, Pongsakorn, Pathanin Chantree, Salisa Chaimon, Nattaya Torungkitmangmi, Parisa Prathaphan, Jittiporn Ruangtong, Phornphan Sornchuer, Nattaya Thongsepee, Kant Sangpairoj, and Poom Adisakwattana. 2022. "Molecular Cloning and Characterization of a Fasciola gigantica Nuclear Receptor Subfamily 1 (FgNR1)" Pathogens 11, no. 12: 1458. https://doi.org/10.3390/pathogens11121458
APA StyleMartviset, P., Chantree, P., Chaimon, S., Torungkitmangmi, N., Prathaphan, P., Ruangtong, J., Sornchuer, P., Thongsepee, N., Sangpairoj, K., & Adisakwattana, P. (2022). Molecular Cloning and Characterization of a Fasciola gigantica Nuclear Receptor Subfamily 1 (FgNR1). Pathogens, 11(12), 1458. https://doi.org/10.3390/pathogens11121458