
Occurrence of Mycoplasma parvum in German Pigs of Different Age Groups Using a Novel Quantitative Real-Time PCR Assay



Abstract
1. Introduction
2. Materials and Methods
2.1. Blood Samples and DNA Preparation
2.2. 16S rDNA and Amplicon Sequencing
2.3. Phylogenetic Analysis
2.4. M. suis and Ca. M. haemosuis qPCR
2.5. Primer Design
2.6. Cloning of the qPCR Fragment (qPCR-Positive Control)
2.7. Quantitative Real-Time PCR
2.8. Intra- and Interassay Repeatability
3. Results
3.1. Identification of Mycoplasma parvum Positive Samples
3.2. Development of a M. parvum-Specific Quantitative Real-Time PCR
3.3. Hemotrophic mycoplasma Infection in Fattening Pigs, Sows, and Boars
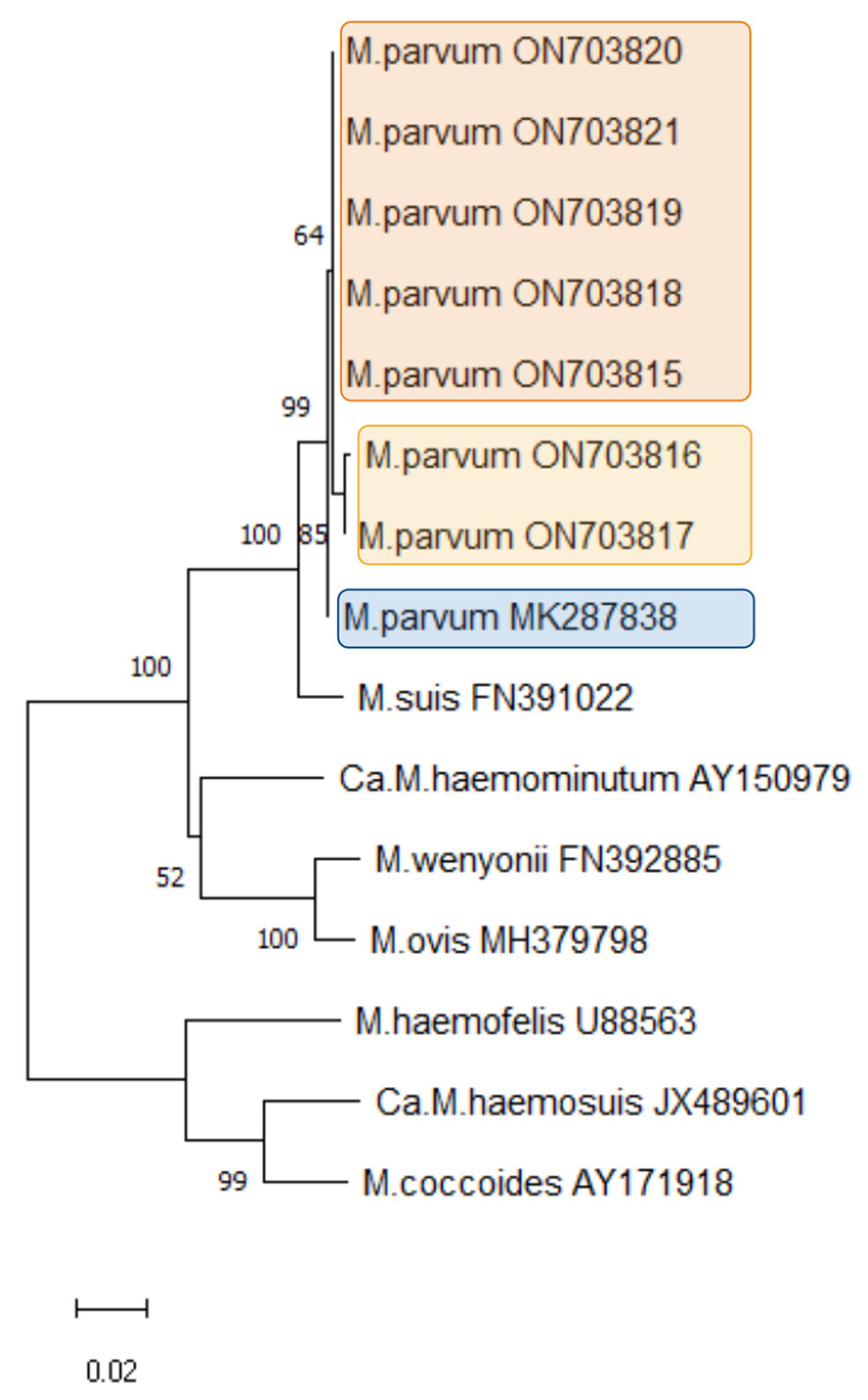
3.4. Mycoplasma parvum Phylogeny
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Messick, J.B. Hemotrophic mycoplasmas (hemoplasmas): A review and new insights into pathogenic potential. Vet. Clin. Pathol. 2004, 33, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Hattori, N.; Kuroda, M.; Katano, H.; Takuma, T.; Ito, T.; Arai, N.; Yanai, R.; Sekizuka, T.; Ishii, S.; Miura, Y.; et al. Candidatus Mycoplasma haemohominis in Human, Japan. Emerg. Infect. Dis. 2020, 26, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Hoelzle, L.E. Haemotrophic mycoplasmas: Recent advances in Mycoplasma suis. Vet. Microbiol. 2008, 130, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Splitter, E.J. Eperythrozoon suis n. sp. and Eperythrozoon parvum n. sp., 2 new blood parasites of swine. Science 1950, 111, 513–514. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Shi, T.; Xu, L.; Wei, W.; Lu, F.; Zhang, X.; Yuan, X.; Li, J.; Lv, J.; Fang, W. Identification of a novel Hemoplasma species from pigs in Zhejiang province, China. J. Vet. Med. Sci. 2017, 79, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Thongmeesee, K.; Kamkong, P.; Thanee, S.; Wattanapansak, S.; Kaewthamasorn, M.; Tiawsirisup, S. Molecular detection and genetic analysis of porcine haemoplasmas in commercial pig farms from Thailand reveal a putative novel species. Transbound. Emerg. Dis. 2022, 69, e2028–e2040. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.G.; Kwon, O.D.; Kwak, D. Prevalence and phylogenetic analysis of hemoplasma species in domestic pigs in Korea. Parasites Vectors 2019, 12, 378. [Google Scholar] [CrossRef]
- Stadler, J.; Ade, J.; Ritzmann, M.; Hoelzle, K.; Hoelzle, L.E. Detection of a novel haemoplasma species in fattening pigs with skin alterations, fever and anaemia. Vet. Rec. 2020, 187, 66. [Google Scholar] [CrossRef]
- Ade, J.; Stadler, J.; Ritzmann, M.; Zübert, C.; Hoelzle, K.; Hoelzle, L.E. Occurrence of ‘Candidatus Mycoplasma haemosuis’ in fattening pigs, sows and piglets in Germany using a novel gap-based quantitative real-time PCR assay. BMC Vet. Res. 2022, 18, 40. [Google Scholar] [CrossRef]
- Petri, F.A.M.; Sonalio, K.; de Souza Almeida, H.M.; Ferraz, M.E.S.; Storino, G.Y.; de Souza, M.R.; André, M.R.; de Oliveira, L.G. Porcine hemothropic mycoplasmas infection associated with productive impact in intensive pig production. Porcine Health Manag. 2020, 6, 33. [Google Scholar] [CrossRef]
- Stadler, J.; Jannasch, C.; Mack, S.L.; Dietz, S.; Zöls, S.; Ritzmann, M.; Hoelzle, K.; Hoelzle, L.E. Clinical and haematological characterisation of Mycoplasma suis infections in splenectomised and non-splenectomised pigs. Vet. Microbiol. 2014, 172, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Stadler, J.; Ade, J.; Hermanns, W.; Ritzmann, M.; Wentzel, S.; Hoelzle, K.; Hoelzle, L.E. Clinical, haematological and pathomorphological findings in Mycoplasma suis infected pigs. BMC Vet. Res. 2021, 17, 214. [Google Scholar] [CrossRef] [PubMed]
- Henry, S.C. Clinical observations on eperythrozoonosis. J. Am. Vet. Med. Assoc. 1979, 174, 601–603. [Google Scholar]
- Seamer, J. Studies with Eperythrozoon parvum Splitter, 1950. Parasitology 1960, 50, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Barnett, S.F. Eperythrozoon parvum in pigs in Kenya. Bull. Epizoot. Dis. Afr. 1963, 11, 185–195. [Google Scholar] [PubMed]
- Do Nascimento, N.C.; dos Santos, A.P.; Chu, Y.; Guimaraes, A.M.; Baird, A.N.; Weil, A.B.; Messick, J.B. Microscopy and genomic analysis of Mycoplasma parvum strain Indiana. Vet. Res. 2014, 45, 86. [Google Scholar] [CrossRef]
- Do Nascimento, N.C.; Guimaraes, A.M.S.; Dos Santos, A.P.; Chu, Y.; Marques, L.M.; Messick, J.B. RNA-Seq based transcriptome of whole blood from immunocompetent pigs (Sus scrofa) experimentally infected with Mycoplasma suis strain Illinois. Vet. Res. 2018, 49, 49. [Google Scholar] [CrossRef]
- Sonalio, K.; Perles, L.; Gatto, I.R.H.; do Amaral, R.B.; Almeida, H.M.S.; Galdeano, J.V.B.; Vieira, R.F.C.; André, M.R.; de Oliveira, L.G. Genetic diversity of emerging hemotropic mycoplasmas in domestic pigs from Brazil. Transbound. Emerg. Dis. 2021, 68, 1162–1174. [Google Scholar] [CrossRef]
- Tasker, S.; Helps, C.R.; Day, M.J.; Gruffydd-Jones, T.J.; Harbour, D.A. Use of real-time PCR to detect and quantify Mycoplasma haemofelis and “Candidatus Mycoplasma haemominutum” DNA. J. Clin. Microbiol. 2003, 41, 439–441. [Google Scholar] [CrossRef]
- Willi, B.; Boretti, F.S.; Cattori, V.; Tasker, S.; Meli, M.L.; Reusch, C.; Lutz, H.; Hofmann-Lehmann, R. Identification, molecular characterization, and experimental transmission of a new hemoplasma isolate from a cat with hemolytic anemia in Switzerland. J. Clin. Microbiol. 2005, 43, 2581–2585. [Google Scholar] [CrossRef]
- Meli, M.L.; Kaufmann, C.; Zanolari, P.; Robert, N.; Willi, B.; Lutz, H.; Hofmann-Lehmann, R. Development and application of a real-time TaqMan(®) qPCR assay for detection and quantification of ‘Candidatus Mycoplasma haemolamae’ in South American camelids. Vet. Microbiol. 2010, 146, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Meli, M.L.; Willi, B.; Dreher, U.M.; Cattori, V.; Knubben-Schweizer, G.; Nuss, K.; Braun, U.; Lutz, H.; Hofmann-Lehmann, R. Identification, molecular characterization, and occurrence of two bovine hemoplasma species in Swiss cattle and development of real-time TaqMan quantitative PCR assays for diagnosis of bovine hemoplasma infections. J. Clin. Microbiol. 2010, 48, 3563–3568. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, A.M.; Vieira, R.F.; Poletto, R.; Vemulapalli, R.; Santos, A.P.; de Moraes, W.; Cubas, Z.S.; Santos, L.C.; Marchant-Forde, J.N.; Timenetsky, J.; et al. A quantitative TaqMan PCR assay for the detection of Mycoplasma suis. J. Appl. Microbiol. 2011, 111, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Ade, J.; Ritzmann, M.; Wöstmann, C.; Eddicks, M.; Reese, S.; Hoelzle, K.; Hoelzle, L.E.; Stadler, J. Update on shedding and transmission routes of porcine haemotrophic mycoplasmas in naturally and experimentally infected pigs. Porcine Health Manag. 2021, 7, 49. [Google Scholar] [CrossRef]
- Stadler, J.; Willi, S.; Ritzmann, M.; Eddicks, M.; Ade, J.; Hoelzle, K.; Hoelzle, L.E. Detection of Mycoplasma suis in pre-suckling piglets indicates a vertical transmission. BMC Vet. Res. 2019, 15, 252. [Google Scholar] [CrossRef]
- Volokhov, D.V.; Norris, T.; Rios, C.; Davidson, M.K.; Messick, J.B.; Gulland, F.M.; Chizhikov, V.E. Novel hemotrophic mycoplasma identified in naturally infected California sea lions (Zalophus californianus). Vet. Microbiol. 2011, 149, 262–268. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Hoelzle, L.E.; Helbling, M.; Hoelzle, K.; Ritzmann, M.; Heinritzi, K.; Wittenbrink, M.M. First LightCycler real-time PCR assay for the quantitative detection of Mycoplasma suis in clinical samples. J. Microbiol. Methods 2007, 70, 346–354. [Google Scholar] [CrossRef]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Ade, J.; Niethammer, F.; Schade, B.; Schilling, T.; Hoelzle, K.; Hoelzle, L.E. Quantitative analysis of Mycoplasma wenyonii and ‘Candidatus Mycoplasma haemobos’ infections in cattle using novel gapN-based realtime PCR assays. Vet. Microbiol. 2018, 220, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Barker, E.N.; Tasker, S.; Day, M.J.; Warman, S.M.; Woolley, K.; Birtles, R.; Georges, K.C.; Ezeokoli, C.D.; Newaj-Fyzul, A.; Campbell, M.D.; et al. Development and use of real-time PCR to detect and quantify Mycoplasma haemocanis and “Candidatus Mycoplasma haematoparvum” in dogs. Vet. Microbiol. 2010, 140, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Elbers, A.R.; Geudeke, M.J.; van Rossem, H.; Kroon, M.C.; Counotte, G.H. Haematology and biochemistry reference values for sows kept under modern management conditions. Vet. Q. 1994, 16, 127–130. [Google Scholar] [CrossRef]
- Schollenberger, A.; Degórski, A.; Bielecki, W.; Stempniak, M. Lymphocyte subpopulations in peripheral blood of pregnant sows. Arch. Vet. Pol. 1992, 32, 35–46. [Google Scholar]
- Luppi, P. How immune mechanisms are affected by pregnancy. Vaccine 2003, 21, 3352–3357. [Google Scholar] [CrossRef]
- Gatto, I.R.H.; Sonálio, K.; Amaral, R.B.D.; Morés, N.; Dalla Costa, O.A.; André, M.R.; de Oliveira, L.G. High frequency and molecular characterization of porcine hemotrophic mycoplasmas in Brazil. Vet. Microbiol. 2019, 231, 33–39. [Google Scholar] [CrossRef]
- Watanabe, Y.; Fujihara, M.; Suzuki, J.; Sasaoka, F.; Nagai, K.; Harasawa, R. Prevalence of swine hemoplasmas revealed by real-time PCR using 16S rRNA gene primers. J. Vet. Med. Sci. 2012, 74, 1315–1318. [Google Scholar] [CrossRef][Green Version]
- Tagawa, M.; Takeuchi, T.; Fujisawa, T.; Konno, Y.; Yamamoto, S.; Matsumoto, K.; Yokoyama, N.; Inokuma, H. A clinical case of severe anemia in a sheep coinfected with Mycoplasma ovis and ‘Candidatus Mycoplasma haemovis’ in Hokkaido, Japan. J. Vet. Med. Sci. 2012, 74, 99–102. [Google Scholar] [CrossRef]
- Hornok, S.; Hajtós, I.; Meli, M.L.; Farkas, I.; Gönczi, E.; Meili, T.; Hofmann-Lehmann, R. First molecular identification of Mycoplasma ovis and ‘Candidatus M. haemoovis’ from goat, with lack of haemoplasma PCR-positivity in lice. Acta Vet. Hung. 2012, 60, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Roblejo-Arias, L.; Díaz-Sánchez, A.A.; Corona-González, B.; Meli, M.L.; Fonseca-Rodríguez, O.; Rodríguez-Mirabal, E.; Marrero-Perera, R.; Vega-Cañizares, E.; Lobo-Rivero, E.; Hofmann-Lehmann, R. First molecular evidence of Mycoplasma haemocanis and ‘Candidatus Mycoplasma haematoparvum’ infections and its association with epidemiological factors in dogs from Cuba. Acta Trop. 2022, 228, 106320. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Díaz, V.L.; Silvestre-Ferreira, A.C.; Vilhena, H.; Pastor, J.; Francino, O.; Altet, L. Prevalence and co-infection of haemotropic mycoplasmas in Portuguese cats by real-time polymerase chain reaction. J. Feline Med. Surg. 2013, 15, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.; Marques, V.; Correia, J.H.; Neto, I.; Bráz, B.S.; Rodrigues, C.; Martins, T.; Rosado, R.; Ferreira, J.P.; Santos-Reis, M.; et al. Molecular detection of haemotropic Mycoplasma species in urban and rural cats from Portugal. J. Feline Med. Surg. 2015, 17, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Ritzmann, M.; Grimm, J.; Heinritzi, K.; Hoelzle, K.; Hoelzle, L.E. Prevalence of Mycoplasma suis in slaughter pigs, with correlation of PCR results to hematological findings. Vet. Microbiol. 2009, 133, 84–91. [Google Scholar] [CrossRef]
- Brissonnier, M.; Normand, V.; Lebret, A.; Moalic, P.Y.; Guyomard, A.S.; Bachy, V.; Berton, P.; Auvigne, V.; Bouchet, F.; Boulbria, G. Frequency of infection with Mycoplasma suis in gestating sows using qPCR on ten commercial French herds, and impact of the infection on clinical, haematological and biochemical parameters. Porcine Health Manag. 2020, 6, 13. [Google Scholar] [CrossRef]
- Hoelzle, L.E.; Zeder, M.; Felder, K.M.; Hoelzle, K. Pathobiology of Mycoplasma suis. Vet. J. 2014, 202, 20–25. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| HM Species and Name in the Phylogenetic Tree | Acc. Number (Version) | Host Species | Country of Origin |
|---|---|---|---|
| Mycoplasma parvum | |||
| M. parvum ON703815 * | ON703815.1 | pig | Germany |
| M. parvum ON703816 * | ON703816.1 | pig | Germany |
| M. parvum ON703817 * | ON703817.1 | pig | Germany |
| M. parvum ON703818 * | ON703818.1 | pig | Germany |
| M. parvum ON703819 * | ON703819.1 | pig | Germany |
| M. parvum ON703820 * | ON703820.1 | pig | Germany |
| M. parvum ON703821 * | ON703821.1 | pig | Germany |
| M. parvum MK287838 | MK287838.1 | pig | Brazil |
| M. suis FN391022 | FN391022.1 | pig | Germany |
| Ca. M. haemosuis JX489601 | JX489601.1 | pig | China |
| M. wenyonii FN392885 | FN392885.1 | cattle | Germany |
| M. ovis MH379798 | MH379798.1 | sheep | Brazil |
| M. coccoides AY171918.1 | AY171918.1 | mouse | USA |
| M. haemofelis U88563 | U88563.2 | cat | USA |
| Ca. M. haemominutum AY150979 | AY150979.1 | cat | South Africa |
| Intra-Assay Repeatability | Interassay Repeatability | |
|---|---|---|
| Plasmid Copy Number | Mean CT (±SD) * | Mean CT (±SD) |
| 107 | 15.23 (±0.44) | 15.02 (±0.92) |
| 106 | 19.01 (±0.27) | 18.94 (±0.92) |
| 105 | 22.78 (±0.11) | 21.92 (±0.94) |
| 104 | 26.20 (±0.15) | 25.25 (±0.96) |
| 103 | 29.93 (±0.07) | 29.28 (±0.75) |
| 102 | 32.81 (±0.73) | 31.89 (±0.99) |
| 101 | 35.92 (±0.81) | 35.51 (±0.54) |
| Fattening Pigs | ||||||
|---|---|---|---|---|---|---|
| M. parvum1 | M. suis2 | Ca. M. haemosuis 2 | ||||
| qPCR positive pigs (total) | 72/200 (36.00%) | 38/200 (19.00%) | 35/200 (17.50%) | |||
| bacterial blood-loads (per mL blood) | Average: | 9.34 × 106 | 7.62 × 107 | 1.61 × 105 | ||
| Range: | 3.41 × 103–4.82 × 108 | 9.58 × 102–1.14 × 109 | 5.52 × 103–1.55 × 106 | |||
| Co-infections | ||||||
| M. parvum | + | M. suis | 17/200 (8.50%) | |||
| M. parvum | + | Ca. M. haemosuis | 19/200 (9.50%) | |||
| M. suis | + | Ca. M. haemosuis | 12/200 (6.00%) | |||
| all species (triple infection) | 5/200 (2.50%) | |||||
| Sows | ||||||
| M. parvum1 | M. suis1 | Ca. M. haemosuis 1 | ||||
| qPCR positive pigs (total) | 15/60 (25.00%) | 4/60 (6.67%) | 13/60 (21.67%) | |||
| bacterial blood-loads (per mL blood) | Average: | 1.18 × 106 | 3.58 × 104 | 4.83 × 105 | ||
| Range: | 5.48 × 103–1.53 × 107 | 5.66 × 103–5.49 × 104 | 2.39 × 103–3.56 × 106 | |||
| Co-infections | ||||||
| M. parvum | + | M. suis | 0/60 (0.00%) | |||
| M. parvum | + | Ca. M. haemosuis | 3/60 (5.00%) | |||
| M. suis | + | Ca. M. haemosuis | 1/60 (1.66%) | |||
| all species (triple infection) | 0/60 (0.00%) | |||||
| Boars | ||||||
| M. parvum1 | M. suis3 | Ca. M. haemosuis 3 | ||||
| qPCR positive pigs (total) | 8/183 (4.37%) | 0/183 (0.00%) | 0/183 (0.00%) | |||
| bacterial blood-loads (per mL blood) | Average: | 1.12 × 105 | / | / | ||
| Range: | 6.23 × 103–6.26 × 106 | / | / | |||
| Co-infections | ||||||
| M. parvum | + | M. suis | 0/183 (0.00%) | |||
| M. parvum | + | Ca. M. haemosuis | 0/183 (0.00%) | |||
| M. suis | + | Ca. M. haemosuis | 0/183 (0.00%) | |||
| all species (triple infection) | 0/183 (0.00%) | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ade, J.; Hoelzle, K.; Stadler, J.; Ritzmann, M.; Hoelzle, L.E. Occurrence of Mycoplasma parvum in German Pigs of Different Age Groups Using a Novel Quantitative Real-Time PCR Assay. Pathogens 2022, 11, 1374. https://doi.org/10.3390/pathogens11111374
Ade J, Hoelzle K, Stadler J, Ritzmann M, Hoelzle LE. Occurrence of Mycoplasma parvum in German Pigs of Different Age Groups Using a Novel Quantitative Real-Time PCR Assay. Pathogens. 2022; 11(11):1374. https://doi.org/10.3390/pathogens11111374
Chicago/Turabian StyleAde, Julia, Katharina Hoelzle, Julia Stadler, Mathias Ritzmann, and Ludwig E. Hoelzle. 2022. "Occurrence of Mycoplasma parvum in German Pigs of Different Age Groups Using a Novel Quantitative Real-Time PCR Assay" Pathogens 11, no. 11: 1374. https://doi.org/10.3390/pathogens11111374
APA StyleAde, J., Hoelzle, K., Stadler, J., Ritzmann, M., & Hoelzle, L. E. (2022). Occurrence of Mycoplasma parvum in German Pigs of Different Age Groups Using a Novel Quantitative Real-Time PCR Assay. Pathogens, 11(11), 1374. https://doi.org/10.3390/pathogens11111374