
Prevalence of Pathogenic Leptospira spp. in Non-Volant Small Mammals of Hutan Lipur Sekayu, Terengganu, Malaysia
 ,
,  ,
,
Abstract
:1. Introduction
2. Methods
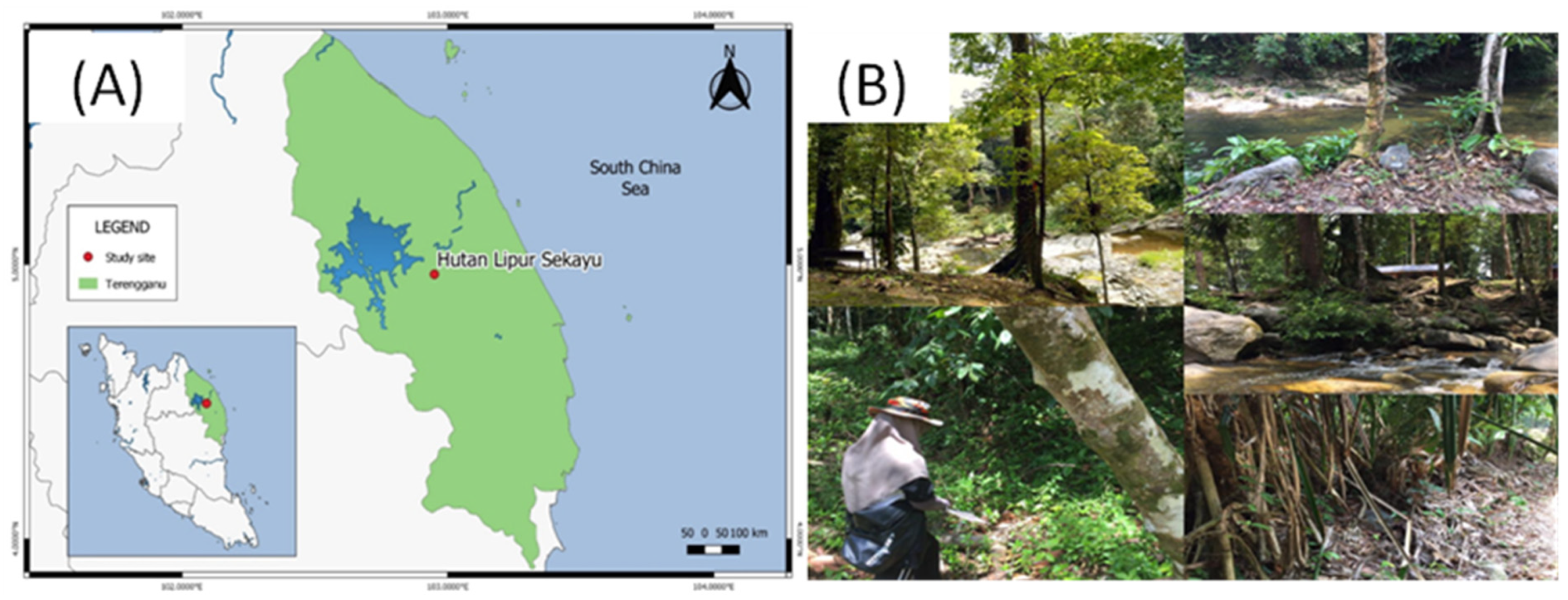
2.1. Study Area
2.2. Animal Trapping and Sampling Collection
2.3. Ethical Statement
2.4. DNA Extraction
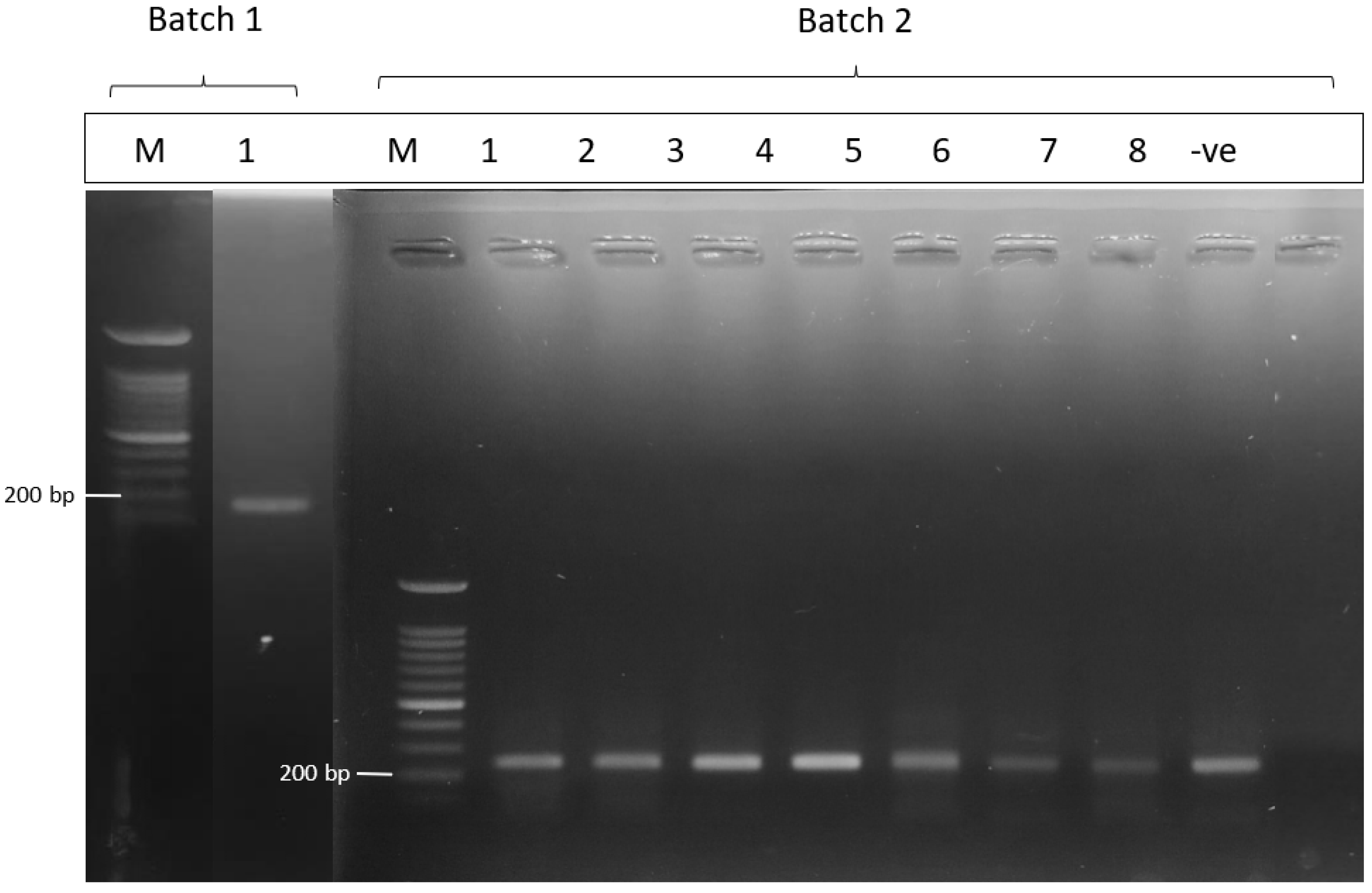
2.5. PCR Detection of Pathogenic Leptospira spp. and Sequencing
2.6. Data Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gentry, A.H. Tropical Forest Biodiversity: Distributional Patterns and Their Conservational Significance. Oikos 1992, 63, 19–28. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853. [Google Scholar] [CrossRef] [PubMed]
- Nazir Khan, N.K.; Mohd Yunus, Z. Sustainable Forest Management in Peninsular Malaysia. In Status of Biological Diversity in Malaysia and Threat Assessment of Plant Species in Malaysia; Chua, L.S.L., Kirton, L.G., Saw, L.G., Eds.; Forest Research Institute Malaysia (FRIM): Selangor Darul Ehsan, Malaysia, 2007; pp. 229–241. [Google Scholar]
- Department of Wildlife and National Parks (DWNP), Peninsular Malaysia. Red List of Mammals for Peninsular Malaysia: Red List Mammalia Semenanjung Malaysia, 1st ed.; Department of Wildlife and National Parks (DWNP), Peninsular Malaysia: Kuala Lumpur, Malaysia, 2009. [Google Scholar]
- Francis, C.; Barrett, P. Guide to the Mammals of Southeast Asia, 1st ed.; Princeton University Press: Princeton, NJ, USA, 2008. [Google Scholar]
- Costa, F.; Porter, F.H.; Rodrigues, G.; Farias, H.; de Faria, M.T.; Wunder, E.A.; Childs, J.E. Infections by Leptospira interrogans, Seoul virus, and Bartonella spp. among Norway rats (Rattus norvegicus) from the urban slum environment in Brazil. Vector-Borne Zoonotic Dis. 2014, 14, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battersby, S. Rodents as carriers of diseases. In Rodent Pests and Their Control; Buckle, A.P., Smith, R.H., Eds.; CABI International: Wallingford, UK, 2015; pp. 81–101. [Google Scholar]
- Cosson, J.-F.; Picardeau, M.; Mielcarek, M.; Tatard, C.; Chaval, Y.; Suputtamongkol, Y.; Buchy, P.; Jittapalapong, S.; Herbreteau, V.; Morand, S. Epidemiology of Leptospira Transmitted by Rodents in Southeast Asia. PLoS Negl. Trop. Dis. 2014, 8, e2902. [Google Scholar] [CrossRef]
- Loan, H.K.; Van Cuong, N.; Takhampunya, R.; Kiet, B.T.; Campbell, J.; Them, L.N.; Carrique-Mas, J.J. How important are rats as vectors of leptospirosis in the Mekong Delta of Vietnam? Vector-Borne Zoonotic Dis. 2015, 15, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Levett, P.N. Leptospirosis. Clin. Microbiol. Rev. 2001, 14, 296–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, B. Leptospira and Leptospirosis. Curr. Top. Microbiol. Immunol. 2015, 387, 1–293. [Google Scholar]
- Costa, F.; Wunder, E.A., Jr.; De Oliveira, D.; Bisht, V.; Rodrigues, G.; Reis, M.G.; Ko, A.I.; Begon, M.; Childs, J.E. Patterns in Leptospira Shedding in Norway Rats (Rattus norvegicus) from Brazilian Slum Communities at High Risk of Disease Transmission. PLoS Negl. Trop. Dis. 2015, 9, e0003819. [Google Scholar] [CrossRef] [Green Version]
- Casanovas-Massana, A.; Hamond, C.; Santos, L.A.; Oliveira, D.; Hacker, K.; Balassiano, I.; Costa, F.; Medeiros, M.A.; Reis, M.G.; Ko, A.; et al. Leptospira yasudae sp. nov. and Leptospira stimsonii sp. nov., two new species of the pathogenic group isolated from environmental sources. Int. J. Syst. Evol. Microbiol. 2020, 70, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Haake, D.A.; Levett, P.N. Leptospirosis in humans. Curr. Top. Microbiol. Immunol. 2015, 387, 65–97. [Google Scholar] [CrossRef] [Green Version]
- Felzemburgh, R.D.; Ribeiro, G.S.; Costa, F.; Reis, R.B.; Hagan, J.E.; Melendez, A.X.; Ko, A.I. Prospective study of leptospirosis transmission in an urban slum community: Role of poor environment in repeated exposures to the Leptospira agent. PLoS Negl. Trop. Dis. 2014, 8, e2927. [Google Scholar] [CrossRef] [PubMed]
- Benacer, D.; Zain, S.N.M.; Amran, F.; Galloway, R.L.; Thong, K.L. Isolation and molecular characterization of Leptospira interrogans and Leptospira borgpetersenii isolates from the urban rat populations of Kuala Lumpur, Malaysia. Am. J. Trop. Med. Hyg. 2013, 88, 704–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blasdell, K.R.; Morand, S.; Perera, D.; Firth, C. Association of rodent-borne Leptospira spp. with urban environments in Malaysian Borneo. PLoS Negl. Trop. Dis. 2019, 13, e0007141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed-Hassan, S.N.; Bahaman, A.R.; Mutalib, A.R.; Khairani-Bejo, S. Prevalence of pathogenic leptospires in rats from selected locations in Peninsular Malaysia. Res. J. Anim. Sci. 2012, 6, 12–25. [Google Scholar] [CrossRef] [Green Version]
- Suut, L.; Mazlan, A.; Arif, M.T.; Katip, T.; Nor Aliza, A.R.; Haironi, Y. Serovar diversity of Leptospira sp. infecting wild rodents in Sarawak, Malaysia. Trop. Biomed. 2018, 35, 252–258. [Google Scholar]
- Ismail, S.; Wahab NZ, A.; Badya, N.; Rahman NI, A.; Yeo, C.C.; Latif AZ, A.; Haque, M. A study on the presence of pathogenic Leptospira spp. in environmental water samples obtained from selected recreational areas in Terengganu, Malaysia. Res. J. Pharm. Technol. 2014, 7, 1153–1157. [Google Scholar]
- The Source of Malaysia’s Official Statistics. 2020 Sept 4 [cited 18 January 2021]. In: Department of Statistic Malaysia Official Portal—DOSM. Department of Statistic Malaysia-: Malaysia. [About 2 Screens]. Available online: https://www.dosm.gov.my/v1/index.php (accessed on 15 January 2021).
- Francis, C.M. Identification key for mammal species. In A Field Guide to the Mammals of South-East Asia; Krystyna, M., Ed.; New Holland Publishers: London, UK, 2008; p. 392. [Google Scholar]
- Mills, J.N.; Childs, J.E.; Ksiazek, T.G.; Peters, C.J.; Velleca, W.M. Methods for Trapping and Sampling Small Mammals for Virologic Testing; U.S Dept. of Health & Human Service, Public Health Service, Centres for Disease Control and Prevention: Atlanta, GA, USA, 1995. [Google Scholar]
- Haake, D.A.; Chao, G.; Zuerner, R.L.; Barnett, J.K.; Barnett, D.; Mazel, M.; Matsunaga, J.; Levett, P.N.; Bolin, C.A. The leptospiral major outer membrane protein LipL32 is a lipoprotein expressed during mammalian infection. Infect. Immun. 2000, 68, 2276–2285. [Google Scholar] [CrossRef] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Backstedt, B.T.; Buyuktanir, O.; Lindow, J.C.; Wunder, E.A., Jr.; dos Reis, M.G.; Usmani-Brown, S.; Ledizet, M.; Ko, A.; Pal, U. Efficient Detection of Pathogenic Leptospires Using 16S Ribosomal RNA. PLoS ONE 2015, 10, e0128913. [Google Scholar] [CrossRef] [Green Version]
- Stoddard, R.A.; Gee, J.E.; Wilkins, P.P.; McCaustland, K.; Hoffmaster, A.R. Detection of pathogenic Leptospira spp. through Taq-Man polymerase chain reaction targeting the lipL32 gene. Diagn. Microbiol. Infect. Dis. 2009, 64, 247–255. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Version 4.0.5 [Software]. 2021. Available online: https://www.R-project.org (accessed on 15 May 2021).
- Aghová, T.; Kimura, Y.; Bryja, J.; Dobigny, G.; Granjon, L.; Kergoat, G.J. Fossils know it best: Using a new set of fossil calibrations to improve the temporal phylogenetic framework of murid rodents (Rodentia: Muridae). Mol. Phylogenetics Evol. 2018, 128, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Munian, K.; Azman, S.M.; Ruzman, N.A.; Fauzi, N.F.M.; Zakaria, A.N. Diversity and composition of volant and non-volant small mammals in northern Selangor State Park and adjacent forest of Peninsular Malaysia. Biodivers. Data J. 2020, 8, e50304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, K.; Bagchi, R. Eat in or take away—Seed predation and removal by rats (Muridae) during a fruiting event in a dipterocarp rainforest. Raffles Bull. Zool. 2005, 53, 281–286. [Google Scholar]
- Rahim NA, A.; Ahmad NI, I.; Zakaria, A.A.; Pesiu, E.; Salam, M.R.; Mamat, M.A.; Abdullah, M.T. Brief survey of non-volant small mammals on Pulau Perhentian Besar, Terengganu, Malaysia. J. Sustain. Sci. Manag. 2016, 2016, 19–25. [Google Scholar]
- Yusof, M.A.; Mohd-Taib, F.S.; Ishak, S.N.; Md-Nor, S.; Md-Sah, S.A.; Mohamed, N.Z.; Sekawi, Z. Microhabitat factors influenced the prevalence of pathogenic Leptospira spp. in small mammal host. EcoHealth 2019, 16, 260–274. [Google Scholar] [CrossRef]
- Ridzlan, F.R.; Bahaman, A.R.; Khairani-Bejo, S.; Mutalib, A.R. Detection of pathogenic Leptospira from selected environment in Kelantan and Terengganu, Malaysia. Trop. Biomed. 2010, 27, 632–638. [Google Scholar]
- Moreno, L.Z.; Miraglia, F.; Kremer, F.S.; Eslabao, M.R.; Dellagostin, O.A.; Lilenbaum, W.; Freitas, J.C.; Vasconcellos, S.A.; Heinemann, M.B.; Moreno, A.M. Comparative genomics of pathogenic Leptospira interrogans serovar Canicola isolated from swine and human in Brazil. Memórias Do Inst. Oswaldo Cruz 2018, 113, 126–129. [Google Scholar] [CrossRef] [Green Version]
- ElliEllis, W.A. Control of canine leptospirosis in Europe: Time for a change? Vet. Rec. 2010, 167, 602–605. [Google Scholar] [CrossRef] [Green Version]
- Daud, A.; Fuzi, N.M.; Arshad, M.M.; Kamarudin, S.; Mohammad, W.M.; Amran, F.; Ismail, N. Leptospirosis seropositivity and its serovars among cattle in Northeastern Malaysia. Vet. World 2018, 11, 840. [Google Scholar] [CrossRef] [Green Version]
- Goh, S.H.; Khor, K.H.; Radzi, R.; Lau, S.F.; Khairani-Bejo, S.; Rahman, M.S.; Roslan, M.A. Shedding and Genetic Diversity of Leptospira spp. From Urban Stray Dogs in Klang Valley, Malaysia. Top. Companion Anim. Med. 2021, 45, 100562. [Google Scholar] [CrossRef]
- Wahab, Z.A. Epidemiology and Current Situation of Leptospirosis in Malaysia. In Proceedings of the Local Authority Environmental Health Conference, Labuan, Malaysia, 8–9 September 2015. [Google Scholar]
- Ganoza, C.; A Matthias, M.; Collins-Richards, D.; Brouwer, K.C.; Cunningham, C.B.; Segura, E.R.; Gilman, R.H.; Gotuzzo, E.; Vinetz, J.M. Determining Risk for Severe Leptospirosis by Molecular Analysis of Environmental Surface Waters for Pathogenic Leptospira. PLoS Med. 2006, 3, e308. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Order and Family | Scientific Name | Common Name | Total Individuals Captured | Total Positive Individuals (%) |
|---|---|---|---|---|
| Scandentia Tupaiidae | Tupaia glis | Common Treeshrew | 10 | 2 (20) |
| Rodentia Muridae | Rattus tiomanicus | Malaysian Wood Rat | 22 | 4 (18.2) |
| Rattus rattus | Black Rat | 1 | 0 (0) | |
| Maxomys rajah | Rajah Spiny Rat | 4 | 1 (25) | |
| Maxomys whiteheadi | Whitehead’s Spiny Rat | 1 | 1 (100) | |
| Sundamys muelleri | Muller’s Giant Sunda Rat | 2 | 1 (50) | |
| Rodentia Sciuridae | Collasciurus notatus | Plaintain Squirrel | 4 | 0 (0) |
| Sundasciurus tenuis | Slender Squirrel | 1 | 0 (0) | |
| Total | 45 | 9 (20) |
| Lane | Species | Percent ID | Query Cover | Accession No. |
|---|---|---|---|---|
| 1 (Batch 1) (R. tiomanicus) | Leptospira interrogans | 98.9% | 99% | CP044513.1 |
| 1 (Batch 2) M. whiteheadi | Leptospira interrogans | 97.6% | 99% | KY356922.1 |
| 2 (R. tiomanicus) | Leptospira interrogans | 98% | 99% | KY356922.1 |
| 3 (S. muelleri) | Uncultured Leptospira sp. | 97.5 | 98% | MG831575.1 |
| 5 (R. tiomanicus) | Uncultured Leptospira sp. | 98.3% | 97% | MG831575.1 |
| 6 (R. tiomanicus) | Leptospira interrogans | 95.9% | 99% | KY356922.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shafie, N.J.; Halim, N.S.A.; Awoniyi, A.M.; Zalipah, M.N.; Md-Nor, S.; Nazri, M.U.I.A.; Costa, F. Prevalence of Pathogenic Leptospira spp. in Non-Volant Small Mammals of Hutan Lipur Sekayu, Terengganu, Malaysia. Pathogens 2022, 11, 1300. https://doi.org/10.3390/pathogens11111300
Shafie NJ, Halim NSA, Awoniyi AM, Zalipah MN, Md-Nor S, Nazri MUIA, Costa F. Prevalence of Pathogenic Leptospira spp. in Non-Volant Small Mammals of Hutan Lipur Sekayu, Terengganu, Malaysia. Pathogens. 2022; 11(11):1300. https://doi.org/10.3390/pathogens11111300
Chicago/Turabian StyleShafie, Nur Juliani, Najma Syahmin Abdul Halim, Adedayo Michael Awoniyi, Mohamed Nor Zalipah, Shukor Md-Nor, Mohd Ulul Ilmie Ahmad Nazri, and Federico Costa. 2022. "Prevalence of Pathogenic Leptospira spp. in Non-Volant Small Mammals of Hutan Lipur Sekayu, Terengganu, Malaysia" Pathogens 11, no. 11: 1300. https://doi.org/10.3390/pathogens11111300
APA StyleShafie, N. J., Halim, N. S. A., Awoniyi, A. M., Zalipah, M. N., Md-Nor, S., Nazri, M. U. I. A., & Costa, F. (2022). Prevalence of Pathogenic Leptospira spp. in Non-Volant Small Mammals of Hutan Lipur Sekayu, Terengganu, Malaysia. Pathogens, 11(11), 1300. https://doi.org/10.3390/pathogens11111300