1. Introduction
In the context of climate change, globalization, and enhanced human traveling, arboviruses continue to pose a threat to public health [
1]. Birds are important reservoirs for arboviruses such as the West Nile (WNV) and Usutu (USUV) viruses. These are mosquito-borne viruses belonging to the
Flaviviridae family (
Flavivirus genus) and members of the Japanese encephalitis virus serocomplex. The Sindbis virus (SINV), also vectored by mosquitoes, is included in the
Togaviridae family (
Alphavirus genus) [
2].
Since its first report in Romania in 1996, WNV circulation has been recorded yearly, with significant outbreaks among humans [
3,
4]. Serological evidence was also recorded during the following years in domestic and wild birds [
5,
6]. Although WNV causes disease and mortality every year, in Romania, it remains passively monitored solely in humans, birds, and horses, with acute neurological cases being tested.
The first introduction of USUV in Europe was described in Austria in 2001, associated with the high mortality of European blackbirds (
Turdus merula) and great gray owls (
Strix nebulosa), followed by a retrospective study in Italy on bird tissue samples [
7,
8]. To date, the virus circulation has been recorded in birds from several European countries, such as Hungary, Italy, and Germany [
9]. In Romania, USUV antibodies were recently documented for the first time in a domestic dog, but their presence in humans, birds, or other susceptible hosts has not yet been demonstrated [
10]. In addition, the co-circulation of WNV and USUV was also reported in Europe in 30 species of birds belonging to 11 orders [
11].
SINV was reported for the first time in Europe in 1965 when antibodies were detected in humans from Italy and Finland and in birds in the Volga Delta [
12,
13]. The only report from Romania dates back to 1975 when a low seroprevalence was being registered in humans [
14].
The medical importance of arboviruses lies in their zoonotic potential. In humans, WNV usually evolves asymptomatically or with flu-like symptoms. In 1% of the infections, humans develop the West Nile neuroinvasive disease, which is associated with meningoencephalitis and, in some cases, fatalities [
15,
16]. USUV was also found to be responsible for severe neuro-invasive infections in humans [
17]. SINV has mostly been associated with disease in humans, and so far, the infections were reported in the northern part of Europe (Sweden, Finland, and Russia) and South Africa [
18]. The disease is associated with fever, arthritis, and a skin rash [
19]. However, USUV and SINV are not considered in the surveillance strategy of Romania.
These viruses have common features, sharing aspects of ecology and epidemiology. Their life cycle is accomplished by ornithophilic mosquitoes that act as vectors and vertebrate hosts. The birds are susceptible to reservoir hosts due to the high and prolonged viremia they develop [
18].
Avian migration is considered to be of great medical importance because of the ability of birds to carry, spread, and transmit a wide range of pathogens, such as viruses, bacteria, fungi, and parasites [
20]. Migratory birds are considered responsible for the introduction and long-distance spread of the arboviruses, while resident birds maintain the viral amplification and local circulation [
21]. Located on the western route of the Eurasian-East African flyway, which connects the Black Sea with the African continent, Romania is an important stopover for migratory birds [
22]. Due to the habitat characteristics, which include wetlands and estuaries, previous studies have mentioned the Danube Delta as being a hotspot for WNV outbreaks in Romania. This habitat is not only favorable for birds but also for the reproduction of mosquitoes [
21]. However, no studies on the circulation of mosquito-borne avian-associated arboviruses in such areas or areas with multi-annual human cases have been published to date.
The aim of our study was to assess the seroprevalence of WNV, USUV, and SINV in blood samples from wild birds and in endemic areas for mosquito-borne viruses from Romania.
2. Materials and Methods
2.1. Sample Collection
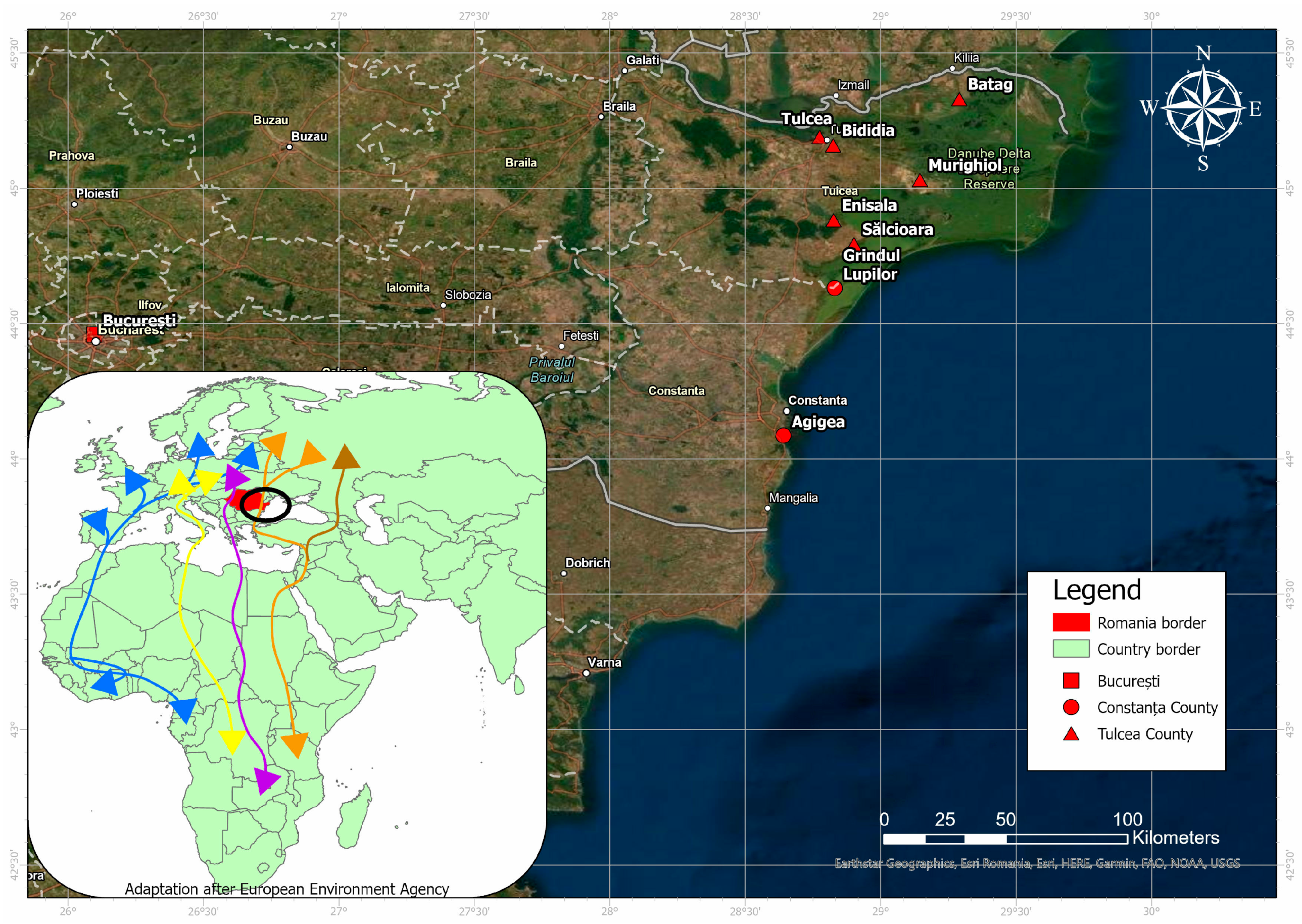
The study was conducted in the southeastern part of Romania between May 2018 and October 2019, as this is one of the major avian migration hotspots in Romania and also the area where most human cases are reported [
23]. The sampling protocol included nine locations in three counties: Constanța, Tulcea counties, and Bucharest.
Blood samples (max 1% of body weight) were collected from wild birds, which were captured by using standard mist nets. The following data were recorded for each bird: the date and location of capture, species, age, and gender. Depending on the migratory behavior of the wild birds, they were divided into long-distance and short-distance migrants. Blood samples were collected from the jugular vein and centrifuged at 8000 rpm for 10 min. The serum was collected and stored at −80 °C until further analysis.
2.2. Serological Analysis
2.2.1. ELISA
All serum samples were screened at the Department of Parasitology and Parasitic Diseases of the University of Agricultural Sciences and Veterinary Medicine of Cluj-Napoca, using the ELISA method for WNV. We used the commercial kit INGEZIM West Nile COMPAC (Eurofins Technologies, Madrid, Spain), targeting specific antibodies against protein E. The method was performed according to the manufacturer’s instructions.
2.2.2. Viruses and Cells
Vero cells (L0015 Collection of Cell Lines in Veterinary Medicine (CCLV), Friedrich- Loeffler-Institut, Greifswald—Insel Riems, Germany) were routinely cultivated in a minimal essential medium (MEM: MEM Hank’s Salts/MEM Earles’ Salts, New York, NY, USA), with 10% fetal calf serum (FCS) at 37 °C with 5% CO2. The viruses used were WNV lineage II Germany/2019/T167-20 (LR743455.1, MN921233), Usutu Europe 3 (HE599647), and Sindbis Edsbyn.
2.2.3. Serum Neutralization Test (SNT)
Depending on the volume of sera available from the birds with positive or equivocal IgG ELISA index values for WNV, serum samples were further analyzed by SNT for WNV, USUV, and SINV. All sera were heat-inactivated before the analyses (30 min at 56 °C) at the Friedrich-Loeffler-Institut, Germany.
SNT was performed in microtiter plates as described in the OIE terrestrial manual. Briefly, two-fold dilution series starting with a 1:5 dilution was made on all the serum samples. Serum dilutions were incubated with 100 Tcid50 of the respective virus for 1 h at 37 °C. After incubation, 1 × 104 Vero cells were added to each well, and samples were incubated for 7 days. Cells were fixed and stained with crystal violet to help with cytopathic effect detection. If the volume allowed, serum samples were tested in duplicates with a WNV lineage II, USUV lineage Europe 3, and SINV. For the low-quantity samples, a chosen subset of the viruses was tested. Neutralizing titres were calculated as the geometric mean of the duplicates.
Sera were considered positive if they showed neutralizing titres equal to 1:10 or higher. Between the WNV and USUV sera, they were considered clearly positive for one virus but not the other if the neutralization titre was four-fold higher for that virus compared to the other.
2.3. Statistical Analysis
The statistical analysis was performed using EpiInfoTM 2000 software (version 7.2.0.1., Atlanta, Georgia). The prevalence of WNV and USUV was estimated from the ratio of positive to the total number of samples, with the exact binomial confidence intervals of 95%. We excluded from this analysis the bird species with less than 10 samples.
A Generalized Linear Model (GLM) with a log link function approach was used to determine the directly relative risk of infection of the studied birds with WNV and USUV. For the analysis, we used the positive cases and the total number of samples collected in each location. GLM analysis was used to find which species was the most suitable to be infected among the assessed species with WNV or with USUV.
2.4. Ethical Statement
Live, apparently healthy birds were captured using mist nets and released immediately after sampling. These birds were captured according to the OM 1380/13.07.2019 by specially trained personnel, with ringing permits (No. 740622) from the Romanian Ornithological Centre, Academy of Agriculture and Forestry.
4. Discussion
WNV, USUV, and SINV are mosquito-borne viruses with continuous circulation in Europe. Given the heterogeneity of surveillance methods used in Europe and the cross-reactions that may influence the diagnosis, these viruses remain a threat to public health [
24,
25]. As previously stated, the plaque-reduction neutralization test (PRNT) is recognized as the gold standard method for the serological diagnostics of flaviviruses [
26]. However, Gennaro et al. concluded that serum neutralization tests could successfully replace the PRNT [
27]. Even in serum neutralization tests that relate flaviviruses from the same sero-complex, they cannot be easily distinguished. Thus, infection with one virus leads to cross-reactive antibodies against other viruses in the same sero-complex. In the present study, we aimed to analyze the presence of antibodies for WNV, USUV, and SINV in migratory and resident birds from the southeastern region of Romania. The study was conducted in an area overlapping the main migration routes of wild birds. We also considered Bucharest, the capital city and an endemic area with the highest predicted probability for WNV infections in humans in Romania [
23].
The SNT for WNV in this study was comparable with those from other studies on wild birds conducted in Romania. Previous studies conducted on birds in Romania have shown seroprevalence rates of 3.5% and 8.8% [
5,
28]. Another study from Romania reported a seroprevalence of 32.1% by ELISA, but no further confirmation tests were performed [
6]. In addition to the previous data, the present study included a much larger number of samples belonging to 46 species of wild birds.
This is the first report of seropositivity to USUV in wild birds from Romania. This is supported by the fact that the sera reacted much more strongly against USUV than WNV. Similar to WNV, USUV has the same life cycle, using birds as amplifying hosts for the introduction and spread of the virus [
29]. Recently, USUV antibodies were confirmed in a dog from Romania [
10]. Several sera (2.4%) in this study could not be clearly distinguished as WNV- or USUV-positive as they reacted to both viruses with similar titers in neutralization assays. These could either be due to an infection with yet another virus in the same sero-complex or due to infections with both viruses. Co-circulation is likely as birds whose sera react only to USUV, or much better to USUV, were sampled in locations that also yielded sera reacting only to WNV. This emphasizes their co-circulation, which was already mentioned in previous studies conducted in Europe [
11,
30]. Several studies have previously attempted to find the correlation between coinfections with the aforementioned viruses and the immune response [
31]. Given that both viruses can determine/induce lesions incompatible with life on their own, it would prove quite a challenge to correlate their symptomatology. Moreover, Zhou et al., and Wang et al., discussed the appearance of a superinfection exclusion phenomenon which would determine the inhibition of one of the viruses (in this case, WNV would inhibit the productive replication of USUV within the cell) [
32,
33]. However, additional studies are needed to confirm these theories.
Previous studies suggested a possible sentinel role of the common blackbird in outbreaks of USUV infections, as the virus introduction in Europe was associated with increased mortality in this bird species in all the countries where outbreaks were recorded [
7,
30,
34,
35]. A possible explanation could be the weaker immune system of this species [
36]. The results we obtained in this study suggest the same in the case of WNV, as previously observed in a study conducted in Germany [
37]. Given the recent clinical cases in humans, we suggest testing cases of viral encephalitis for USUV and SINV as well [
17]. Moreover, the confirmation of WNV in blood donors in several European countries should be taken into consideration by the public institutions responsible for the blood test methodology in transfusions [
4,
38].
The presence of SINV in wildlife has already been reported in Europe [
39]. Moreover, the co-circulation of SINV with WNV and USUV was also registered in birds influenced by similar aspects of the life cycle, suggesting similar epidemiological mechanisms [
2]. Since 1975 only one study concerning SINV in Romania was conducted, revealing a prevalence of 0.6% in humans [
14]. The complete absence of a serological response in our study could be related to the relatively small sample size.
To our best knowledge, this is the first large-scale comprehensive study to assess the WNV seropositivity and the first serological confirmation of USUV in wild birds in Romania. Moreover, this is the only study assessing the current seroprevalence of SINV in Romania since 1975.

 ,
, 


{kind=link}
{kind=link}