


Diversity and Exploration of Endophytic Bacilli for the Management of Head Scab (Fusarium graminearum) of Wheat

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Sampling for the Isolation and Identification of Endophytic Bacillus
2.2. In-Vitro Determination of Antagonistic Activity
2.3. Field Trial of Antagonists against FHB
2.4. Strain Characterization for Siderophore Production and Hydrolytic Enzyme Activities
2.5. Detection of Antimicrobial Peptide Gene (s)
2.6. Statistical Analysis
3. Results
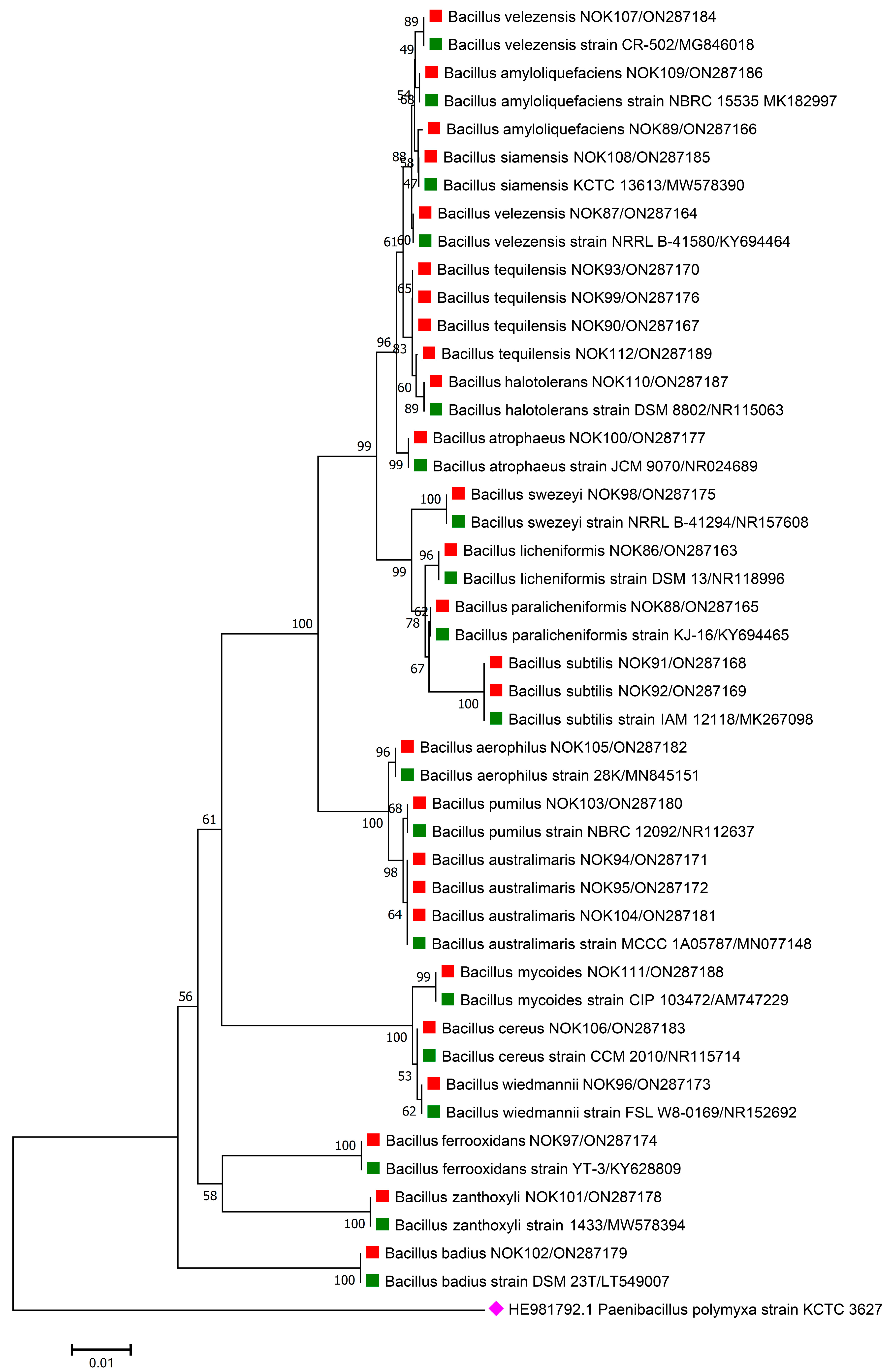
3.1. Species Diversity of Endophytic Bacillus in Tissues of Wheat of Different Genotypes
3.2. Identification and Selection of Antagonistic Endophytes
3.3. Strain Characterization for Potassium Solubilization, Siderophores Release and Hydrolytic Enzyme Activity
3.4. Detection of Antimicrobial Peptides (AMPs) Biosynthesis Associated Genes
3.5. Field Evaluation of Antagonistic Strains against F. graminearum
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaul, N.; Kashyap, P.L.; Kumar, S.; Singh, D.; Singh, G.P. Genetic Diversity and Population Structure of Head Blight Disease Causing Fungus Fusarium graminearum in Northern Wheat Belt of India. J. Fungi 2022, 8, 820. [Google Scholar] [CrossRef] [PubMed]
- Saharan, M.S.; Kumar, H.M.A.; Gurjar, M.S.; Aggarwal, R. Fusarium head blight of wheat in India-variability in pathogens associated and sources of resistance: An overview. Indian Phytopathol. 2021, 74, 345–353. [Google Scholar] [CrossRef]
- Wegulo, S.N.; Baenziger, P.S.; Nopsa, J.H.; Bockus, W.W.; Hallen-Adams, H. Management of Fusarium head blight of wheat and barley. Crop Prot. 2015, 73, 100–107. [Google Scholar] [CrossRef]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium Toxins in Cereals: Occurrence, Legislation, Factors Promoting the Appearance and Their Management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [PubMed]
- Degrassi, V.C.-P.G. Biological Control of Fusarium Head Blight by Bacterial Endophytes and Reduction of Deoxynivalenol in Wheat. Adv. Biochem. Biotechnol. 2020, 5, 10103. [Google Scholar] [CrossRef]
- Tini, F.; Beccari, G.; Onofri, A.; Ciavatta, E.; Gardiner, D.M.; Covarelli, L. Fungicides may have differential efficacies towards the main causal agents of Fusarium head blight of wheat. Pest Manag. Sci. 2020, 76, 3738–3748. [Google Scholar] [CrossRef] [PubMed]
- Mesterházy, Á.; Bartók, T.; Lamper, C. Influence of Wheat Cultivar, Species of Fusarium, and Isolate Aggressiveness on the Efficacy of Fungicides for Control of Fusarium Head Blight. Plant Dis. 2003, 87, 1107–1115. [Google Scholar] [CrossRef]
- Ramírez, M.L.; Chulze, S.; Magan, N. Impact of environmental factors and fungicides on growth and deoxynivalenol production by Fusarium graminearum isolates from Argentinian wheat. Crop Prot. 2004, 23, 117–125. [Google Scholar] [CrossRef]
- Xu, W.; Zhang, L.; Goodwin, P.H.; Xia, M.; Zhang, J.; Wang, Q.; Liang, J.; Sun, R.; Wu, C.; Yang, L. Isolation, Identification, and Complete Genome Assembly of an Endophytic Bacillus velezensis YB-130, Potential Biocontrol Agent against Fusarium graminearum. Front. Microbiol. 2020, 11, 598285. [Google Scholar] [CrossRef]
- Kashyap, P.L.; Kumar, S.; Aggarwal, S.K.; Kaul, N.; Jasrotia, P.; Gupta, A.; Singh, G.P. Resistance inducers and their role in reinforcing wheat defense system against fungal pathogens. J. Cereal Res. 2021, 13, 229–254. [Google Scholar] [CrossRef]
- Fatima, S.; Anjum, T. Identification of a Potential ISR Determinant from Pseudomonas aeruginosa PM12 against Fusarium Wilt in Tomato. Front. Plant Sci. 2017, 8, 848. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Cui, J.; Tian, B.; Cao, S.; Zhang, X.; Chen, H. Resistance risk assessment for Fusarium graminearum to pydiflumetofen, a new succinate dehydrogenase inhibitor. Pest Manag. Sci. 2020, 76, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Hou, Y.; Cai, Y.; Zhang, Y.; Li, Y.; Zhou, M. Whole-genome sequencing reveals that mutations in myosin-5 confer resistance to the fungicide phenamacril in Fusarium graminearum. Sci. Rep. 2015, 5, 8248. [Google Scholar] [CrossRef] [PubMed]
- de Chaves, M.A.; Reginatto, P.; da Costa, B.S.; de Paschoal, R.I.; Teixeira, M.L.; Fuentefria, A.M. Fungicide Resistance in Fusarium graminearum Species Complex. Curr. Microbiol. 2022, 79, 62. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, P.; Kashyap, P.L.; Srivastava, A.K.; Tiwari, R.K. Plant growth promoting and antifungal activity in endophytic Bacillus strains from pearl millet (Pennisetumglaucum). Braz. J. Microbiol. 2020, 51, 229–241. [Google Scholar] [CrossRef]
- Ahlawat, O.P.; Yadav, D.; Kashyap, P.L.; Khippal, A.; Singh, G. Wheat endophytes and their potential role in managing abiotic stress under changing climate. J. Appl. Microbiol. 2022, 132, 2501–2520. [Google Scholar] [CrossRef]
- Pan, D.; Mionetto, A.; Tiscornia, S.; Bettucci, L. Endophytic bacteria from wheat grain as biocontrol agents of Fusarium graminearum and deoxynivalenol production in wheat. Mycotoxin Res. 2015, 31, 137–143. [Google Scholar] [CrossRef]
- Hong, C.E.; Park, J.M. Endophytic bacteria as biocontrol agents against plant pathogens: Current state-of-the-art. Plant Biotechnol. Rep. 2016, 10, 353–357. [Google Scholar] [CrossRef]
- Frank, A.C.; Guzmán, J.P.S.; Shay, J.E. Transmission of Bacterial Endophytes. Microorganisms 2017, 5, 70. [Google Scholar] [CrossRef]
- Kushwaha, P.; Kashyap, P.L.; Bhardwaj, A.K.; Kuppusamy, P.; Srivastava, A.K.; Tiwari, R.K. Bacterial endophyte mediated plant tolerance to salinity: Growth responses and mechanisms of action. World J. Microbiol. Biotechnol. 2020, 36, 26. [Google Scholar] [CrossRef]
- Kushwaha, P.; Kashyap, P.L.; Kuppusamy, P.; Srivastava, A.K.; Tiwari, R.K. Functional characterization of endophytic bacilli from pearl millet (Pennisetumglaucum) and their possible role in multiple stress tolerance. Plant Biosyst.-Int. J. Deal. Asp. Plant Biol. 2020, 154, 503–514. [Google Scholar] [CrossRef]
- Shahid, I.; Han, J.; Hanooq, S.; Malik, K.A.; Borchers, C.H.; Mehnaz, S. Profiling of Metabolites of Bacillus spp. And Their Application in Sustainable Plant Growth Promotion and Biocontrol. Front. Sustain. Food Syst. 2021, 5, 605195. [Google Scholar] [CrossRef]
- Romero, D.; De Vicente, A.; Rakotoaly, R.H.; Dufour, S.E.; Veening, J.-W.; Arrebola, E.; Cazorla, F.M.; Kuipers, O.P.; Paquot, M.; Pérez-García, A. The Iturin and Fengycin Families of Lipopeptides Are Key Factors in Antagonism of Bacillus subtilis toward Podosphaerafusca. Mol. Plant Microbe Interact. 2007, 20, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Kong, H.; Buyer, J.S.; Lakshman, D.K.; Lydon, J.; Kim, S.D.; Roberts, D.P. Isolation and partial characterization of Bacillus subtilis ME488 for suppression of soil borne pathogens of cucumber and pepper. Appl. Microbiol. Biotechnol. 2008, 80, 115–123. [Google Scholar] [CrossRef]
- Mora, I.; Cabrefiga, J.; Montesinos, E. Cyclic Lipopeptide Biosynthetic Genes and Products, and Inhibitory Activity of Plant-Associated Bacillus against Phytopathogenic Bacteria. PLoS ONE 2015, 10, e0127738. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, C. Fengycins, Cyclic Lipopeptides from Marine Bacillus subtilis Strains, Kill the Plant-Pathogenic Fungus Magnaporthegrisea by Inducing Reactive Oxygen Species Production and Chromatin Condensation. Appl. Environ. Microbiol. 2018, 84, e00445-18. [Google Scholar] [CrossRef]
- Kalai-Grami, L.; Ben Slimane, I.; Mnari-Hattab, M.; Rezgui, S.; Aouani, M.A.; Hajlaoui, M.R.; Limam, F. Protective effect of Bacillus amyloliquefaciens against infections of Citrus aurantium seedlings by Phomatracheiphila. World J. Microbiol. Biotechnol. 2014, 30, 529–538. [Google Scholar] [CrossRef]
- Jiao, R.; Cai, Y.; He, P.; Munir, S.; Li, X.; Wu, Y.; Wang, J.; Xia, M.; He, P.; Wang, G.; et al. Bacillus amyloliquefaciens YN201732 Produces Lipopeptides with Promising Biocontrol Activity Against Fungal Pathogen Erysiphe cichoracearum. Front. Cell. Infect. Microbiol. 2021, 11, 598999. [Google Scholar] [CrossRef]
- Cheffi, M.; Bouket, A.C.; Alenezi, F.N.; Luptakova, L.; Belka, M.; Vallat, A.; Rateb, M.E.; Tounsi, S.; Triki, M.A.; Belbahri, L. Olea europaea L. Root Endophyte Bacillus velezensis OEE1 Counteracts Oomycete and Fungal Harmful Pathogens and Harbours a Large Repertoire of Secreted and Volatile Metabolites and Beneficial Functional Genes. Microorganisms 2019, 7, 314. [Google Scholar] [CrossRef]
- Tao, A.; Pang, F.; Huang, S.; Yu, G.; Li, B.; Wang, T. Characterisation of endophytic Bacillus thuringiensis strains isolated from wheat plants as biocontrol agents against wheat flag smut. Biocontrol Sci. Technol. 2014, 24, 901–924. [Google Scholar] [CrossRef]
- Rong, S.; Xu, H.; Li, L.; Chen, R.; Gao, X.; Xu, Z. Antifungal activity of endophytic Bacillus safensis B21 and its potential application as a biopesticide to control rice blast. Pestic. Biochem. Physiol. 2020, 162, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Munakata, Y.; Gavira, C.; Genestier, J.; Bourgaud, F.; Hehn, A.; Slezack-Deschaumes, S. Composition and functional comparison of vetiver root endophytic microbiota originating from different geographic locations that show antagonistic activity towards Fusarium graminearum. Microbiol. Res. 2021, 243, 126650. [Google Scholar] [CrossRef]
- da Luz, W.C.; Stockwell, C.A.; Bergstrom, G.C. Biological control of Fusarium graminearum. In Fusarium Head Blight of Wheat and Barley; Leonard, K.J., Bushnell, W.R., Eds.; APS Press: St. Paul, MN, USA, 2003; pp. 381–394. [Google Scholar]
- Sharma, A.; Kashyap, P.L.; Srivastava, A.K.; Bansal, Y.K.; Kaushik, R. Isolation and characterization of halotolerant bacilli from chickpea (Cicer arietinum L.) rhizosphere for plant growth promotion and biocontrol traits. Eur. J. Plant Pathol. 2018, 153, 787–800. [Google Scholar] [CrossRef]
- Cappuccino, J.G.; Sherman, N. Microbiology: A Laboratory Manual, 10th ed.; Benjamin/Cunnings Science Publishing: San Francisco, CA, USA, 2013; p. 560. [Google Scholar]
- Kumar, S.; Kashyap, P.L.; Singh, R.; Srivastava, A.K. Preservation and Maintenance of Microbial Cultures. In Analyzing Microbes. Springer Protocols Handbooks; Arora, D., Das, S., Sukumar, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 135–152. [Google Scholar] [CrossRef]
- Pospiech, A. A Versatile Quick-Prep of Genomic DNA from Gram-Positive Bacteria. Trends Genet. 1995, 11, 217–218. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-F.; Xu, X.-M.; Zhu, Q.; Deng, M.-C.; Feng, L.; Peng, J.; Yuan, J.-P.; Wang, J.-H. An effective method for the detoxification of cyanide-rich wastewater by Bacillus sp. CN-22. Appl. Microbiol. Biotechnol. 2014, 98, 3801–3807. [Google Scholar] [CrossRef]
- Mora, I.; Cabrefiga, J. Antimicrobial peptide genes in Bacillus strains from plant environments. Int. Microbiol. 2011, 14, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Ramarathnam, R.; Bo, S.; Chen, Y.; Fernando, W.G.; Xuewen, G.; De Kievit, T. Molecular and biochemical detection of fengycin and bacillomycin D-producing Bacillus spp., antagonistic to fungal pathogens of canola and wheat. Can. J. Microbiol. 2007, 53, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Stack, R.W.; McMullen, M.P. A Visual Scale to Estimate Severity of Fusarium Head Blight in Wheat; North Dakota State University Extension Service: Fargo, ND, USA, 1995. [Google Scholar]
- Schwyn, B.; Neilands, J. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Hu, X.F.; Chen, J.; Guo, J.F. Two phosphate and potassium solubilizing bacterial isolated from Tiannu Mountain, Zhijiang, China. World J. Microbiol. Biotechnol. 2006, 22, 983–990. [Google Scholar] [CrossRef]
- Dutilloy, E.; Oni, F.E.; Esmaeel, Q.; Clément, C.; Barka, E.A. Plant Beneficial Bacteria as Bioprotectants against Wheat and Barley Diseases. J. Fungi 2022, 8, 632. [Google Scholar] [CrossRef]
- Becher, R.; Hettwer, U.; Karlovsky, P.; Deising, H.B.; Wirsel, S.G.R. Adaptation of Fusarium graminearum to Tebuconazole Yielded Descendants Diverging for Levels of Fitness, Fungicide Resistance, Virulence, and Mycotoxin Production. Phytopathology 2010, 100, 444–453. [Google Scholar] [CrossRef]
- Sessitsch, A.; Reiter, B.; Pfeifer, U.; Wilhelm, E. Cultivation-independent population analysis of bacterial endophytes in three potato varieties based on eubacterial and Actinomycetes-specific PCR of 16S rRNA genes. FEMS Microbiol. Ecol. 2002, 39, 23–32. [Google Scholar] [CrossRef]
- Pang, F.; Tao, A.; Ayra-Pardo, C.; Wang, T.; Yu, Z.; Huang, S. Plant organ- and growth stage-diversity of endophytic bacteria with potential as biofertilisers isolated from wheat (Triticum aestivum L.). BMC Plant Biol. 2022, 22, 276. [Google Scholar] [CrossRef]
- Robinson, R.J.; Fraaije, B.A.; Clark, I.M.; Jackson, R.W.; Hirsch, P.R.; Mauchline, T.H. Endophytic bacterial community composition in wheat (Triticum aestivum) is determined by plant tissue type, developmental stage and soil nutrient availability. Plant Soil 2016, 405, 381–396. [Google Scholar] [CrossRef]
- Marag, P.S.; Suman, A.; Gond, S. Prospecting endophytic bacterial colonisation and their potential plant growth promoting attributes in hybrid maize (Zea mays L.). Int. J. Curr. Microbiol. App. Sci. 2018, 7, 1292–1304. [Google Scholar] [CrossRef]
- Wang, G.; Meng, J.; Tian, T.; Xiao, X.; Zhang, B.; Xiao, Y. Endophytic Bacillus velezensis strain B-36 is a potential biocontrol agent against lotus rot caused by Fusarium oxysporum. J. Appl. Microbiol. 2020, 128, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Hazarika, D.J.; Goswami, G.; Gautom, T.; Parveen, A.; Das, P.; Barooah, M.; Boro, R.C. Lipopeptide mediated biocontrol activity of endophytic Bacillus subtilis against fungal phytopathogens. BMC Microbiol. 2019, 19, 71. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Guan, Y.; Dong, Y.; Zhao, L.; Rong, S.; Chen, W.; Lv, M.; Xu, H.; Gao, X.; Chen, R.; et al. Isolation and evaluation of endophytic Bacillus tequilensis GYLH001 with potential application for biological control of Magnaportheoryzae. PLoS ONE 2018, 13, e0203505. [Google Scholar] [CrossRef]
- Larran, S.; Simon, M.R.; Moreno, M.V.; Siurana, M.S.; Perelló, A. Endophytes from wheat as biocontrol agents against tan spot disease. Biol. Control 2016, 92, 17–23. [Google Scholar] [CrossRef]
- Kashyap, P.L.; Srivastava, A.K.; Tiwari, S.P.; Kumar, S. Microbes for Climate Resilient Agriculture; John Wiley & Sons: Hoboken, NJ, USA, 2018; p. 358. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Kashyap, P.L.; Srivastava, M. The Plant Microbiome in Sustainable Agriculture; John Wiley & Sons: Hoboken, NJ, USA, 2021; p. 320. [Google Scholar] [CrossRef]
- Rana, K.L.; Kour, D.; Kaur, T.; Sheikh, I.; Yadav, A.N.; Kumar, V.; Suman, A.; Dhaliwal, H.S. Endophytic Microbes from Diverse Wheat Genotypes and Their Potential Biotechnological Applications in Plant Growth Promotion and Nutrient Uptake. Proc. Natl. Acad. Sci. India Sect. B Boil. Sci. 2020, 90, 969–979. [Google Scholar] [CrossRef]
- Solanki, M.K.; Kumar, S.; Pandey, A.K.; Srivastava, S.; Singh, R.K.; Kashyap, P.L.; Srivastava, A.K.; Arora, D.K. Diversity and antagonistic potential of Bacillus spp. associated to the rhizosphere of tomato for the management of Rhizoctoniasolani. Biocontrol Sci. Technol. 2012, 22, 203–217. [Google Scholar] [CrossRef]
- Besset-Manzoni, Y.; Joly, P.; Brutel, A.; Gerin, F.; Soudière, O.; Langin, T.; Prigent-Combaret, C. Does in vitro selection of biocontrol agents guarantee success in planta? A study case of wheat protection against Fusarium seedling blight by soil bacteria. PLoS ONE 2019, 14, e0225655. [Google Scholar] [CrossRef]
- Gao, X.; Gong, Y.; Huo, Y.; Han, Q.; Kang, Z.; Huang, L. Endophytic Bacillus subtilis Strain E1R-J Is a Promising Biocontrol Agent for Wheat Powdery Mildew. Biomed Res. Int. 2015, 2015, 462645. [Google Scholar] [CrossRef]
- Comby, M.; Gacoin, M.; Robineau, M.; Rabenoelina, F.; Ptas, S.; Dupont, J.; Profizi, C.; Baillieul, F. Screening of wheat endophytes as biological control agents against Fusarium head blight using two different in vitro tests. Microbiol. Res. 2017, 202, 11–20. [Google Scholar] [CrossRef]
- Jamali, H.; Sharma, A.; Kashyap, P.L.; Hi, R.; Srivastava, A.K. Exploitation of Multifarious Abiotic Stresses, Antagonistic Activity and Plant Growth Promoting Attributes of Bacillus amyloliquefaciens AH53 for Sustainable Agriculture Production. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 751–763. [Google Scholar] [CrossRef]
- Shukla, N.; Singh, D.; Tripathi, A.; Kumari, P.; Gupta, R.K.; Singh, S.; Shanker, K.; Singh, A. Synergism of endophytic Bacillus subtilis and Klebsiella aerogenes modulates plant growth and bacoside biosynthesis in Bacopa monnieri. Front. Plant Sci. 2022, 13, 896856. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Verma, A.; Pal, G.; Anubha; White, J.F.; Verma, S.K. Seed Endophytic Bacteria of Pearl Millet (Pennisetumglaucum L.) Promote Seedling Development and Defend Against a Fungal Phytopathogen. Front. Microbiol. 2021, 12, 774293. [Google Scholar] [CrossRef] [PubMed]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Kumar, K.; Pal, G.; Verma, A.; Verma, S.K. Seed inhabiting bacterial endophytes of finger millet (Eleusinecoracana L.) promote seedling growth and development, and protect from fungal disease. S. Afr. J. Bot. 2020, 134, 91–98. [Google Scholar] [CrossRef]
- Khanna, A.; Raj, K.; Kumar, P.; Wati, L. Antagonistic and growth-promoting potential of multifarious bacterial endophytes against Fusarium wilt of chickpea. Egypt. J. Biol. Pest Control 2022, 32, 17. [Google Scholar] [CrossRef]
- Agarwal, H.; Dowarah, B.; Baruah, P.M.; Bordoloi, K.S.; Krishnatreya, D.B.; Agarwala, N. Endophytes from Gnetumgnemon L. can protect seedlings against the infection of phytopathogenic bacterium Ralstonia solanacearum as well as promote plant growth in tomato. Microbiol. Res. 2020, 238, 126503. [Google Scholar] [CrossRef]
- Ahmed, A.; Munir, S.; He, P.; Li, Y.; He, P.; Yixin, W.; He, Y. Biocontrol arsenals of bacterial endophyte: An imminent triumph against clubroot disease. Microbiol. Res. 2020, 241, 126565. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, J.; Quadt-Hallmann, A.; Mahaffee, W.F.; Kloepper, J.W. Bacterial endophytes in agricultural crops. Can. J. Microbiol. 1997, 43, 895–914. [Google Scholar] [CrossRef]
- Dogan, G.; Taskin, B. Hydrolytic Enzymes Producing Bacterial Endophytes of Some Poaceae Plants. Pol. J. Microbiol. 2021, 70, 297–304. [Google Scholar] [CrossRef]
- Shafi, J.; Tian, H.; Ji, M. Bacillus species as versatile weapons for plant pathogens: A review. Biotechnol. Biotechnol. Equip. 2017, 31, 446–459. [Google Scholar] [CrossRef]
- Singh, R.K.; Kumar, D.P.; Singh, P.; Solanki, M.K.; Srivastava, S.; Kashyap, P.L.; Kumar, S.; Srivastava, A.K.; Singhal, P.K.; Arora, D.K. Multifarious plant growth promoting characteristics of chickpea rhizosphere associated Bacilli help to suppress soil-borne pathogens. Plant Growth Regul. 2014, 73, 91–101. [Google Scholar] [CrossRef]
- Vinodkumar, S.; Nakkeeran, S.; Renukadevi, P.; Malathi, V.G. Biocontrol Potentials of Antimicrobial Peptide Producing Bacillus Species: Multifaceted Antagonists for the Management of Stem Rot of Carnation Caused by Sclerotiniasclerotiorum. Front. Microbiol. 2017, 8, 446. [Google Scholar] [CrossRef] [PubMed]
- Solanki, M.K.; Singh, R.K.; Srivastava, S.; Kumar, S.; Kashyap, P.L.; Srivastava, A.K. Characterization of antagonistic-potential of two Bacillus strains and their biocontrol activity against Rhizoctoniasolani in tomato. J. Basic Microbiol. 2015, 55, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef]
- Kim, P.I.; Ryu, J.; Kim, Y.H.; Chi, Y.-T. Production of Biosurfactant Lipopeptides Iturin A, Fengycin and Surfactin A from Bacillus subtilis CMB32 for Control of Colletotrichum gloeosporioides. J. Microbiol. Biotechnol. 2010, 20, 138–145. [Google Scholar] [CrossRef]
- Toral, L.; Rodríguez, M.; Béjar, V.; Sampedro, I. Antifungal Activity of Lipopeptides from Bacillus XT1 CECT 8661 against Botrytis cinerea. Front. Microbiol. 2018, 9, 1315. [Google Scholar] [CrossRef]
- Nandhini, S.; Sendhilvel, V.; Babu, S. Endophytic bacteria from tomato and their efficacy against Fusarium oxysporum f.sp. lycopersici, the wilt pathogen. J. Biopest. 2012, 5, 178–185. [Google Scholar]
- Sharma, C.K.; Vishnoi, V.K.; Dubey, R.C.; Maheshwari, D.K. A twin rhizospheric bacterial consortium induces systemic resistance to a phytopathogen Macrophominaphaseolina in mug bean. Rhizosphere 2018, 5, 71–75. [Google Scholar] [CrossRef]
- Hays, S.G.; Patrick, W.G.; Ziesack, M.; Oxman, N.; A Silver, P. Better together: Engineering and application of microbial symbioses. Curr. Opin. Biotechnol. 2015, 36, 40–49. [Google Scholar] [CrossRef]
- Minchev, Z.; Kostenko, O.; Soler, R.; Pozo, M.J. Microbial Consortia for Effective Biocontrol of Root and Foliar Diseases in Tomato. Front. Plant Sci. 2021, 12, 756368. [Google Scholar] [CrossRef] [PubMed]
- Samain, E.; Ernenwein, C.; Aussenac, T.; Selim, S. Effective and durable systemic wheat-induced resistance by a plant-growth-promoting rhizobacteria consortium of Paenibacillus sp. strain B2 and Arthrobacter spp. strain AA against Zymoseptoriatritici and drought stress. Physiol. Mol. Plant Pathol. 2022, 119, 101830. [Google Scholar] [CrossRef]
- Abdeljalil, N.O.-B.; Renault, D.; Gerbore, J.; Vallance, J.; Daami-Remadi, P.R.A.M. Comparative Efficacy of Three Tomato-Associated Rhizobacteria used Singly or in Combination in Suppressing Rhizoctonia Root Rot and Enhancing Tomato Growth. J. Microb. Biochem. Technol. 2016, 8, 110–119. [Google Scholar] [CrossRef]
- Palazzini, J.M.; Alberione, E.; Torres, A.; Donat, C.; Köhl, J.; Chulze, S. Biological control of Fusarium graminearum sensu stricto, causal agent of Fusarium head blight of wheat, using formulated antagonists under field conditions in Argentina. Biol. Control. 2016, 94, 56–61. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Muhae-Ud-Din, G.; Ali, M.A.; Naveed, M.; Naveed, K.; Abbas, A.; Anwar, J.; Tanveer, M.H. Consortium application of endophytic bacteria and fungi improves grain yield and physiological attributes in advanced lines of bread wheat. Turk. J. Agric.-Food Sci. Technol. 2018, 6, 136–144. [Google Scholar] [CrossRef]
- Resti, Z.; Warnita; Liswarni, Y. Endophytic bacterial consortia as biocontrol of purple blotch and plant growth promoters of shallots. IOP Conf. Ser. Earth Environ. Sci. 2021, 741, 012009. [Google Scholar] [CrossRef]
- Sundaramoorthy, S.; Raguchander, T.; Ragupathi, N.; Samiyappan, R. Combinatorial effect of endophytic and plant growth promoting rhizobacteria against wilt disease of Capsicum annum L. caused by Fusarium solani. Biol. Control 2012, 60, 59–67. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Region | Gene | Primer Sequences (5′–3′) | PCR Profile | Product Size (bp) | References |
|---|---|---|---|---|---|
| 16S rRNA | 27F | AGAGTTTGATCMTGGCTCAG | Initial denaturation at 95 °C for 4 min, followed by 35 cycles of denaturation, annealing, and elongation at 95 °C for 1 min, 56 °C for 1 min and 72 °C for 90 s, respectively. The final extension step was done at 72 °C for 10 min | [43] | |
| 1525R | AAGGAGGTGWTCCARCC | ~1500 | |||
| Surfactin | srfA-F | TCGGGACAGGAAGACATCAT CCACTCAAACGGATAATCCTGA | Initial denaturation at 95 °C for 4 min, 35 cycles of 95 °C for 35 s, 58 °C for 30 s, and 72 °C for 45 s, final extension 72 °C for 10 min | 201 | [44] |
| srfA-R | |||||
| Bacillomycin | bmyC-F | TGAAACAAAGGCATATGCTC AAAAATGCATCTGCCGTTCC | Initial denaturation at 94 °C for 4 min, 40 cycles of 95 °C for 60 s, 56 °C for 30 s, and 72 °C for 45 s, final extension 72 °C for 10 min | 395 | [45] |
| bmyC-R | |||||
| Iturin | ItuD-F | GATGCGATCTCCTTGGATGT ATCGTCATGTGCTGCTTGAG | Initial denaturation at 95 °C for 4 min, 35 cycles of 95 °C for 35 s, 60 °C for 30 s, and 72 °C for 45 s, final extension 72 °C for 10 min | 647 | [44] |
| Strain | Wheat Genotype | # PDC (%) | Diameter of Clear Zone (mm) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Siderophore | K-SolubilIzation | Protease | Lipase | Amylase | Chitinase | Pectinase | |||
| NOK09 | DBW187 | *77.3 ± 0.14 b | 20.95 ± 1.01 a | 17.45 ± 1.84 a | 13.50 ± 0.55 a | 21.47 ± 1.71 a | 19.12 ± 1.12 a | 28.43 ± 0.65 a | 22.53 ± 1.01 a |
| NOK16 | DBW187 | 36.2 ± 0.62 h | 10.26 ± 0.14 e | 10.34 ± 2.43 b | 10.08 ± 0.81 c | - | 13.42 ± 1.02 b | 20.12 ± 1.32 b | 18.03 ± 1.05 b |
| NOK33 | PBW343 | 71.9 ± 0.23 c | 20.62 ± 1.41 a | 16.58 ± 1.41 a | 13.14 ± 2.20 a | 20.10 ± 1.04 a | 18.62 ± 1.21 a | 28.22 ± 1.08 a | 22.42 ± 1.20 a |
| NOK47 | Agra Local | 69.04 ± 0.42 d | 13.24 ± 1.43 bc | 9.42 ± 1.31 bc | 10.42 ± 0.55 c | 13.71 ± 1.43 c | 12.02 ± 1.01 bc | 10.12 ± 1.19 f | 13.62 ± 1.08 e |
| NOK52 | Agra Local | 66.66 ± 0.52 e | 11.01 ± 1.12 d | 10.85 ± 1.08 b | 10.21 ± 1.01 c | 11.07 ± 1.76 cd | 11.32 ± 1.72 cd | 14.30 ± 1.33 d | 15.74 ± 1.09 cd |
| NOK59 | Agra Local | 69.04 ± 0.52 d | 10.11 ± 1.08 d | 10.86 ± 0.95 b | 10.41 ± 0.64 c | 12.27 ± 1.08 c | 13.12 ± 1.07 b | 11.40 ± 1.07 f | 13.12 ± 1.02 e |
| NOK68 | HD3086 | **ND | ND | ND | ND | ND | ND | ND | ND |
| NOK72 | HD3086 | 53.4 ± 0.42 g | 14.32 ± 0.88 b | - | 10.70 ± 0.52 c | - | 11.02 ± 1.0 cd | 16.46 ± 0.26 d | 12.20 ± 1.12 ef |
| NOK78 | HD3086 | 61.9 ± 0.56 f | 10.51 ± 1.75 de | 8.95 ± 0.98 c | 9.14 ± 2.08 cd | 12.54 ± 1.23 c | 10.32 ± 1.06 d | 18.73 ± 0.35 c | 11.85 ± 1.21 ef |
| NOK82 | HD3086 | 69.04 ± 0.52 d | 12.35 ± 2.01c | 10.45 ± 1.84 b | 10.20 ± 0.81 c | - | 13.25 ± 1.02 b | 13.43 ± 0.65 e | 11.04 ± 1.01 ef |
| NOK85 | HD3086 | 64.28 ± 0.45 e | 14.35 ± 2.01 b | - | 11.08 ± 0.92 bcd | 15.08 ± 1.27 bc | 12.42 ± 1.52 c | 18.12 ± 1.42 c | 14.04 ± 1.15 cd |
| NOK89 | HD2967 | 61.9 ± 0.58 f | 10.62 ± 2.31 de | 11.08 ± 1.41 b | 10.14 ± 2.05 c | 12.10 ± 1.04 c | 11.22 ± 2.21 cd | 13.42 ± 1.18 e | 16.42 ± 1.22 c |
| NOK95 | HD2967 | 36.2 ± 0.59 h | 13.24 ± 1.20 bc | - | 12.42 ± 0.95 b | 10.71 ± 1.43 d | 12.02 ± 1.07 bc | 13.12 ± 1.09 e | 14.62 ± 1.18 cd |
| NOK103 | HD2967 | 60.3 ± 0.82 fg | 12.01 ± 1.40 bcd | 11.85 ± 1.90 b | 11.21 ± 1.31 bcd | 11.07 ± 1.46 cd | 10.52 ± 1.52 de | 14.30 ± 1.43 e | 15.74 ± 1.09 cd |
| NOK109 | HD2967 | 79.4 ± 0.52 a | 21.11 ± 1.08 a | 17.86 ± 0.95 a | 14.21 ± 1.11 a | 22.07 ± 1.26 a | 19.32 ± 1.72 a | 29.30 ± 1.44 a | 23.74 ± 1.29 a |
| Treatment(s) | *Disease Index (%) | |
|---|---|---|
| 2020–2021 | 2021–2022 | |
| T1 (NOK9 + NFG1) | 39.19 | 43.28 |
| T2 (NOK33 + NFG1) | 34.05 | 37.62 |
| T3 (NOK109 + NFG1) | 30.19 | 32.70 |
| T4 (NOK9 + NOK33 + NFG1) | 26.50 | 29.68 |
| T5 (NOK9 + NOK109 + NFG1) | 21.99 | 26.63 |
| T6 (NOK33+ NOK109 + NFG1) | 16.94 | 23.33 |
| T7 (NOK9 +NOK33 + NOK109 + NFG1) | 11.82 | 11.46 |
| T8 (NFG1 only) | 81.47 | 77.85 |
| T9 (Propiconazole @ 0.1% + NFG1) | 2.75 | 3.31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaul, N.; Kashyap, P.L.; Kumar, S.; Singh, D.; Singh, G.P. Diversity and Exploration of Endophytic Bacilli for the Management of Head Scab (Fusarium graminearum) of Wheat. Pathogens 2022, 11, 1088. https://doi.org/10.3390/pathogens11101088
Kaul N, Kashyap PL, Kumar S, Singh D, Singh GP. Diversity and Exploration of Endophytic Bacilli for the Management of Head Scab (Fusarium graminearum) of Wheat. Pathogens. 2022; 11(10):1088. https://doi.org/10.3390/pathogens11101088
Chicago/Turabian StyleKaul, Noyonika, Prem Lal Kashyap, Sudheer Kumar, Deepti Singh, and Gyanendra Pratap Singh. 2022. "Diversity and Exploration of Endophytic Bacilli for the Management of Head Scab (Fusarium graminearum) of Wheat" Pathogens 11, no. 10: 1088. https://doi.org/10.3390/pathogens11101088
APA StyleKaul, N., Kashyap, P. L., Kumar, S., Singh, D., & Singh, G. P. (2022). Diversity and Exploration of Endophytic Bacilli for the Management of Head Scab (Fusarium graminearum) of Wheat. Pathogens, 11(10), 1088. https://doi.org/10.3390/pathogens11101088