
Introgression of the Powdery Mildew Resistance Genes Pm60 and Pm60b from Triticum urartu to Common Wheat Using Durum as a ‘Bridge’

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Introgression of Disease Resistance Genes
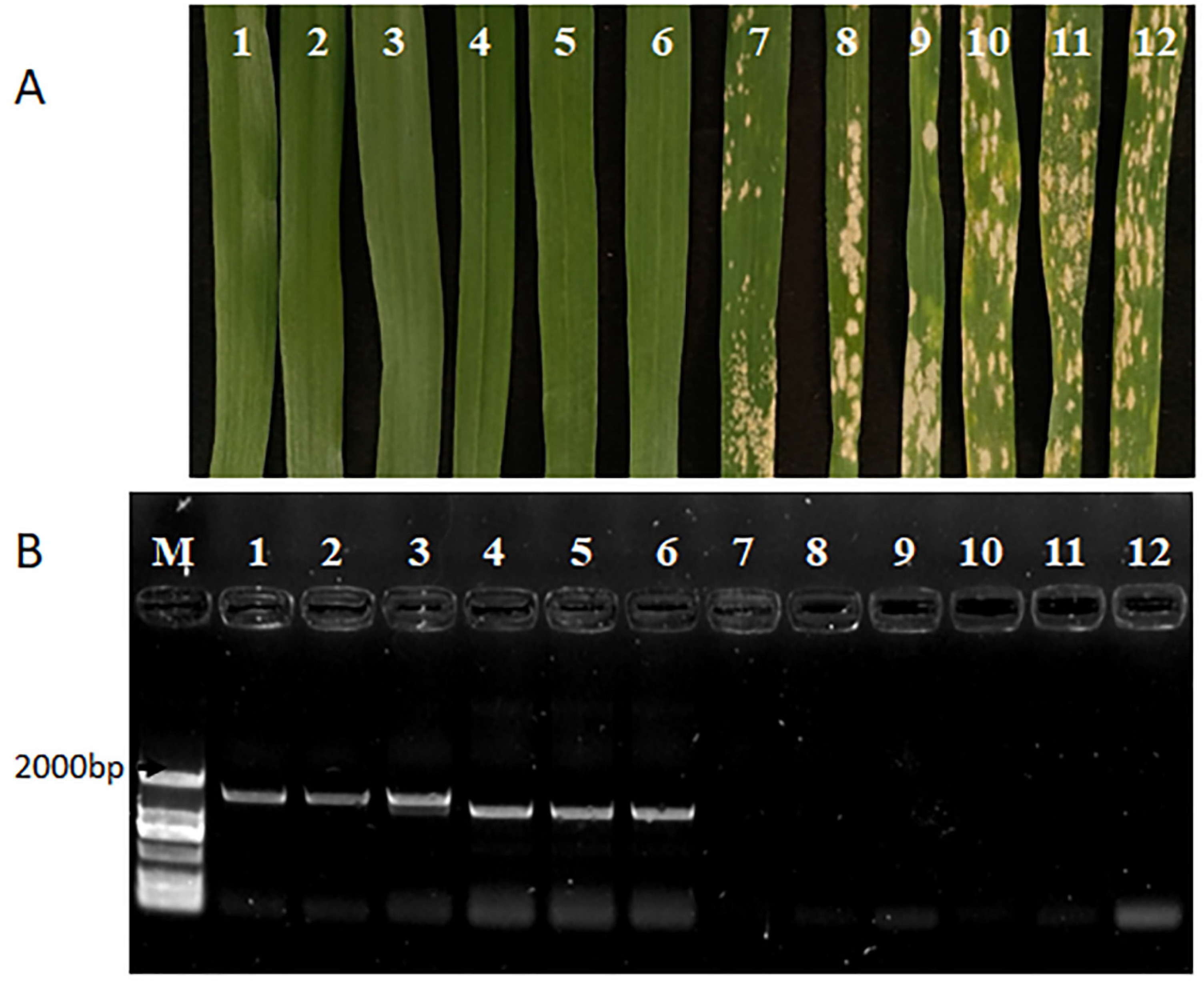
2.3. Bgt Inoculation and Disease Assessment
2.4. Tracing of Pm60 Allele by Molecular Markers
2.5. Analysis of the Introgressed Fragments Flanking Pm60 Locus
3. Results
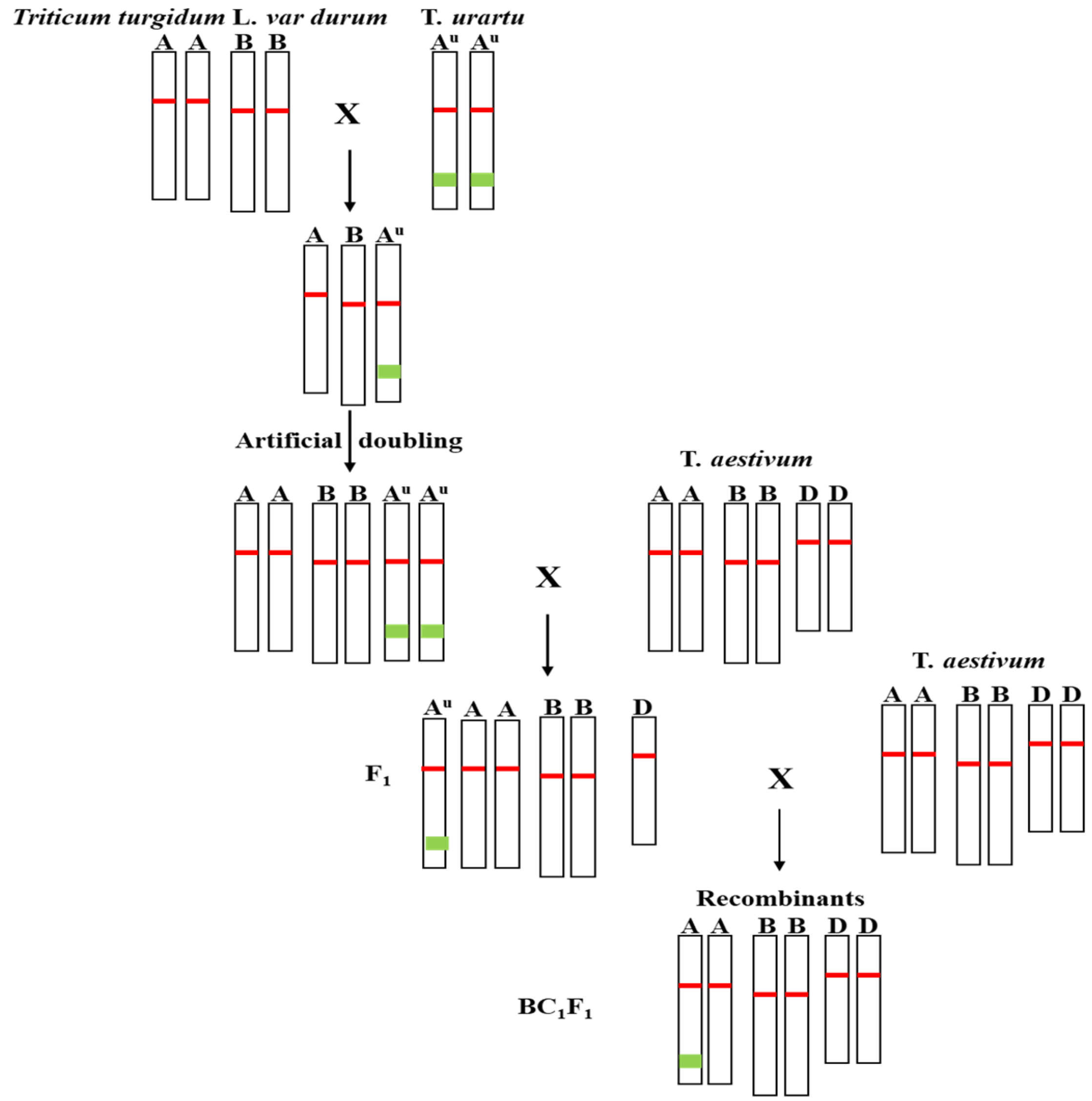
3.1. Synthesizing Hexaploid AABBAuAu Wheat by Crossing T. urartu (AuAu) with Durum (AABB)
3.2. Introgression of Pm60 Alleles into Common Wheat
3.3. Graphical Genotypes Analysis of the Introgressed Chromosome Segments
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Golzar, H.; Shankar, M.; D’Antuono, M. Responses of commercial wheat varieties and differential lines to western Australian powdery mildew (Blumeria graminis f. sp. tritici) populations. Australas. Plant Pathol. 2016, 45, 347–355. [Google Scholar] [CrossRef]
- Wan, W.; Xiao, J.; Li, M.; Tang, X.; Wen, M.; Cheruiyot, A.K.; Li, Y.; Wang, H.; Wang, X. Fine mapping of wheat powdery mildew resistance gene Pm6 using 2B/2G homoeologous recombinants induced by the ph1b mutant. Theor. Appl. Genet. 2020, 133, 1265–1275. [Google Scholar] [CrossRef]
- Li, Y.; Wei, Z.Z.; Fatiukha, A.; Jaiwar, S.; Wang, H.; Hasan, S.; Liu, Z.; Sela, H.; Krugman, T.; Fahima, T. TdPm60 identified in wild emmer wheat is an ortholog of Pm60 and constitutes a strong candidate for PmG16 powdery mildew resistance. Theor. Appl. Genet. 2021, 1–17. [Google Scholar] [CrossRef]
- Li, M.; Dong, L.; Li, B.; Wang, Z.; Xie, J.; Qiu, D.; Li, Y.; Shi, W.; Yang, L.; Wu, Q.; et al. A CNL protein in wild emmer wheat confers powdery mildew resistance. New Phytol. 2020, 228, 1027–1037. [Google Scholar] [CrossRef]
- Yin, H.; Fang, X.; Li, P.; Yang, Y.; Hao, Y.; Liang, X.; Bo, C.; Ni, F.; Ma, X.; Du, X.; et al. Genetic mapping of a novel powdery mildew resistance gene in wild emmer wheat from “Evolution Canyon” in Mt. Carmel Israel. Theor. Appl. Genet. 2021, 134, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Cenci, A.; D’Ovidio, R.; Tanzarella, O.A.; Ceoloni, C.; Porceddu, E. Identification of molecular markers linked to Pm13, an Aegilops longissima gene conferring resistance to powdery mildew in wheat. Theor. Appl. Genet. 1999, 98, 448–454. [Google Scholar] [CrossRef]
- Li, H.; Dong, Z.; Ma, C.; Xia, Q.; Tian, X.; Sehgal, S.; Koo, D.H.; Friebe, B.; Ma, P.; Liu, W. A spontaneous wheat-Aegilops longissima translocation carrying Pm66 confers resistance to powdery mildew. Theor. Appl. Genet. 2020, 133, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Devos, K.M.; Chao, S.; Miller, T.E.; Reader, S.M.; Gale, M.D. RFLP-based maps of the homoeologous group-6 chromosomes of wheat and their application in the tagging of Pm12, a powdery mildew resistance gene transferred from Aegilops speltoides to wheat. Theor. Appl. Genet. 1996, 92, 559. [Google Scholar] [CrossRef]
- Petersen, S.; Lyerly, J.H.; Worthington, M.L.; Parks, W.R.; Cowger, C.; Marshall, D.S.; Brown-Guedira, G.; Paul Murphy, J. Mapping of powdery mildew resistance gene Pm53 introgressed from Aegilops speltoides into soft red winter wheat. Theor. Appl. Genet. 2015, 128, 303–312. [Google Scholar] [CrossRef]
- He, H.; Zhu, S.; Zhao, R.; Jiang, Z.; Ji, Y.; Ji, J.; Qiu, D.; Li, H.; Bie, T. Pm21, encoding a typical CC-NBS-LRR protein, confers broad-spectrum resistance to wheat powdery mildew disease. Mol. Plant 2018, 11, 879–882. [Google Scholar] [CrossRef] [Green Version]
- Zou, S.H.; Wang, H.; Li, Y.W.; Kong, Z.S.; Tang, D.Z. The NB LRR gene Pm60 confers powdery mildew resistance in wheat. New Phytol. 2018, 218, 298–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntosh, R.A.; Zhang, P.; Cowger, C.; Parks, R.; Lagudah, E.S.; Hoxha, S. Rye-derived powdery mildew resistance gene Pm8 in wheat is suppressed by the Pm3 locus. Theor. Appl. Genet. 2011, 123, 359–367. [Google Scholar] [CrossRef]
- Brandolini, A.; Hidalgo, A.; Gabriele, S.; Heun, M. Chemical composition of wild and feral diploid wheats and their bearing on domesticated wheats. J. Cereal Sci. 2015, 63, 122–127. [Google Scholar] [CrossRef]
- Wang, X.; Luo, G.; Yang, W.; Li, Y.; Sun, J.; Zhan, K.; Liu, D.; Zhang, A. Genetic diversity, population structure and marker-trait associations for agronomic and grain traits in wild diploid wheat Triticum urartu. BMC Plant Biol. 2017, 17, 112. [Google Scholar] [CrossRef] [PubMed]
- Brunazzi, A.; Scaglione, D.; Talini, R.F.; Miculan, M.; Magni, F.; Poland, J.; Enrico Pe, M.; Brandolini, A.; Dell’Acqua, M. Molecular diversity and landscape genomics of the crop wild relative Triticum urartu across the Fertile Crescent. Plant J. 2018, 94, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Singh, R.P.; Mujeeb-Kazi, A. Resistance to stripe rust in durum wheats, A-genome diploids, and their amphiploids. Euphytica 1997, 94, 279–286. [Google Scholar] [CrossRef]
- Qiu, Y.C.; Zhou, R.H.; Kong, X.Y.; Zhang, S.S.; Jia, J.Z. Microsatellite mapping of a Triticum urartu Tum. Derived powdery mildew resistance gene transferred to common wheat (Triticum aestivum L.). Theor. Appl. Genet. 2005, 111, 1524–1531. [Google Scholar] [CrossRef] [PubMed]
- Rouse, M.N.; Jin, Y. Stem rust resistance in A-genome diploid relatives of wheat. Plant Dis. 2011, 95, 941–944. [Google Scholar] [CrossRef] [Green Version]
- Grewal, S.; Hubbart-Edwards, S.; Yang, C.; Scholefield, D.; Ashling, S.; Burridge, A.; Wilkinson, P.A.; King, I.P.; King, J. Detection of T. urartu introgressions in wheat and development of a panel of interspecific introgression lines. Front. Plant Sci. 2018, 9, 1565. [Google Scholar] [CrossRef] [PubMed]
- Fricano, A.; Brandolini, A.; Rossini, L.; Sourdille, P.; Wunder, J.; Effgen, S.; Hidalgo, A.; Erba, D.; Piffanelli, P.; Salamini, F. Crossability of Triticum urartu and Triticum monococcum wheats, homoeologous recombination, and description of a panel of interspecific introgression lines. G3 Genes Genomes Genet. 2014, 4, 1931–1941. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Chen, B.; Jiao, J.; Jia, L.; Liu, C. Two novel vesicle-inducing proteins in plastids 1 genes cloned and characterized in Triticum urartu. PLoS ONE 2017, 12, e0170439. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, H.; Zhang, L.; Yuan, Z.; Hao, M.; Zheng, Y. Distant hybridization: A tool for interspecific manipulation of chromosomes. In Alien Gene Transfer in Crop Plants; Springer: New York, NY, USA, 2014; Volume 1, pp. 25–42. [Google Scholar] [CrossRef]
- Ogbonnaya, F.C.; Abdalla, O.; Mujeeb-Kazi, A.; Kazi, A.G.; Xu, S.S.; Gosman, N.; Tsujimoto, H. Synthetic hexaploids: Harnessing species of the primary gene pool for wheat improvement. Plant Breed. Rev. 2013, 37, 35–122. [Google Scholar] [CrossRef]
- Klymiuk, V.; Fatiukha, A.; Huang, L.; Wei, Z.Z.; Kis-Papo, T.; Saranga, Y.; Krugman, T.; Fahima, T. Durum wheat as a bridge between wild emmer wheat genetic resources and bread wheat. In Applications of Genetic and Genomic Research in Cereals; Woodhead Publishing: Sawston, UK, 2019; pp. 201–230. [Google Scholar] [CrossRef]
- Chen, S.; Rouse, M.N.; Zhang, W.; Zhang, X.; Guo, Y.; Briggs, J.; Dubcovsky, J. Wheat gene Sr60 encodes a protein with two putative kinase domains that confers resistance to stem rust. New Phytol. 2020, 225, 948–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, F.; Li, Y.; Yang, B.; Yuan, H.; Jin, C.; Zhou, L.; Pei, H.; Zhao, L.; Li, Y.; Zhou, Y.; et al. Powdery mildew disease resistance and marker-assisted screening at the Pm60 locus in wild diploid wheat Triticum urartu. Crop J. 2020, 8, 252–259. [Google Scholar] [CrossRef]
- Acanda, Y.; Martínez, Ó.; González, M.V.; Prado, M.J.; Rey, M. Highly efficient in vitro tetraploid plant production via colchicine treatment using embryogenic suspension cultures in grapevine (Vitis vinifera cv. Mencía). Plant Cell Tiss. Org. 2015, 123, 547–555. [Google Scholar] [CrossRef]
- Xiao, M.; Song, F.; Jiao, J.; Wang, X.; Xu, H.; Li, H. Identification of the gene Pm47 on chromosome 7BS conferring resistance to powdery mildew in the Chinese wheat landrace Hongyanglazi. Theor. Appl. Genet. 2013, 126, 1397–1403. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, Q.; Ni, Z.; Yang, T.; McIntosh, R.A. Development of SCAR markers linked to the Pm21 gene conferring resistance to powdery mildew in common wheat. Plant Breed. 1999, 118, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Saghai Maroof, M.A.; Biyashev, R.M.; Yang, G.P.; Zhang, Q.; Allard, R.W. Extraordinarily polymorphic microsatellite DNA in barley: Species diversity, chromosomal locations, and population dynamics. Proc. Natl. Acad. Sci. USA 1994, 91, 5466–5470. [Google Scholar] [CrossRef] [Green Version]
- Bassam, B.J.; Caetano-Anollés, G.; Gresshoff, P.M. Fast and sensitive silver staining of DNA in polyacrylamide gels. Anal. Biochem. 1991, 196, 80–83. [Google Scholar] [CrossRef]
- Gao, L.; Koo, D.H.; Juliana, P.; Rife, T.; Singh, D.; da Silva, C.L.; Lux, T.; Dorn, K.M.; Clinesmith, M.; Silva, P.; et al. The Aegilops ventricosa 2N v S segment in bread wheat: Cytology, genomics and breeding. Theor. Appl. Genet. 2021, 134, 529–542. [Google Scholar] [CrossRef]
- Adonina, I.G.; Timonova, E.M.; Salina, E.A. Introgressive Hybridization of Common Wheat: Results and Prospects. Russ. J. Genet. 2021, 57, 390–407. [Google Scholar] [CrossRef]
- Huang, L.; Raats, D.; Sela, H.; Klymiuk, V.; Lidzbarsky, G.; Feng, L.; Krugman, T.; Fahima, T. Evolution and adaptation of wild emmer wheat populations to biotic and abiotic stresses. Annu. Rev. Phytopathol. 2016, 54, 279–301. [Google Scholar] [CrossRef]
- Jiang, J.; Friebe, B.; Gill, B.S. Recent advances in alien gene transfer in wheat. Euphytica 1993, 73, 199–212. [Google Scholar] [CrossRef]
- Younis, A.; Ramzan, F.; Ramzan, Y.; Zulfiqar, F.; Ahsan, M.; Lim, K.B. Molecular markers improve abiotic stress tolerance in crops: A review. Plants 2020, 9, 1374. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zhao, F.; Chen, Y.; Zhang, P.; Zhang, H.; Guo, G.; Xie, J.; Dong, L.; Lu, P.; Li, M.; et al. Bulked segregant CGT-Seq-facilitated map-based cloning of a powdery mildew resistance gene originating from wild emmer wheat (Triticum dicoccoides). Plant Biotechnol. J. 2021, 9, 1288–1290. [Google Scholar] [CrossRef] [PubMed]
- Börner, A.; Ogbonnaya, F.C.; Röder, M.S.; Rasheed, A.; Periyannan, S.; Lagudah, E.S. Aegilops tauschii introgressions in wheat. In Alien Introgression in Wheat; Springer: Cham, Switzerland, 2015; pp. 245–271. [Google Scholar] [CrossRef]
- Naz, A.A.; Bungartz, A.; Serfling, A.; Kamruzzaman, M.; Schneider, M.; Wulff, B.B.; Pillen, K.; Ballvora, A.; Oerke, E.C.; Ordon, F.; et al. Lr21 diversity unveils footprints of wheat evolution and its new role in broad-spectrum leaf rust resistance. Plant Cell Environ. 2021, 44, 3445–3458. [Google Scholar] [CrossRef] [PubMed]
- Periyannan, S.; Moore, J.; Ayliffe, M.; Bansal, U.; Wang, X.; Huang, L.; Deal, K.; Luo, M.; Kong, X.; Bariana, H.; et al. The gene Sr33, an ortholog of barley Mla genes, encodes resistance to wheat stem rust race Ug99. Science 2013, 341, 786–788. [Google Scholar] [CrossRef]
- Li, J.; Wan, H.S.; Yang, W.Y. Synthetic hexaploid wheat enhances variation and adaptive evolution of bread wheat in breeding processes. J. Syst. Evol. 2014, 52, 735–742. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Primer Sequence (5′–3′) | Purpose |
|---|---|---|
| M-Pm60 F M-Pm60 R | CATTAACTTTGAGTTGTTGGA CGGTGATCATACCAGAATTC | Screening of the Pm60 alleles |
| scaf15–5.24 F scaf15–5.24 R | CATCATCACAAAGATACCGAT TCGTCTTTATCTCTGCTCCT | Linkage marker with Pm60 alleles |
| scaf22–5.13 F scaf22–5.13 R | AATGTAGATCGTCTCTTCGC ACACCTTGTGTTTTTGCTCT | Linkage marker with Pm60 alleles |
| scaf7–3.3 F scaf7–3.3 R | CGGTTCCATCTCACATTTTG ATTGAGAGTTCGGGATTTGG | Linkage marker with Pm60 alleles |
| scaf5–4.12 F scaf5–4.12 R | AGATGAAATTGAGCGAAGTT ATAATCAATGTCCACCGAAG | Linkage marker with Pm60 alleles |
| scaf32–5.24 F scaf32–5.24 R | CCACCTCATGAACAACTACC AGGAAACACAACACAACAGG | Linkage marker with Pm60 alleles |
| No. | Male (♂) | Female (♀) | Number of Seedlings | |||
|---|---|---|---|---|---|---|
| Resistant | Susceptible | χ2 (1:1) | p-Value | |||
| Z1 | Fielder | (Mo75/ PI428215)/Xuezao | 17 | 23 | 0.40 | 0.53 |
| Z2 | Xuezao | (Mo75/ PI428215)/Xuezao | 19 | 9 | 6.3 | 0.01 |
| Z5 | Fielder | (Mo75/ PI428315)/Xuezao | 9 | 13 | 0.18 | 0.67 |
| Z7 | Shiluan 02–1 | (Mo75/ PI428215)/Xuezao | 17 | 15 | 0.51 | 0.48 |
| Z10 | Shiluan 02–1 | (Mo75/ PI428315)/Xuezao | 23 | 21 | 0.37 | 0.54 |
| Z14 | Xuezao | (Mo75/ CITR17664)/Xuezao | 22 | 6 | 29.17 | 0.07 × 10–5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Li, Y.; Li, Y.; Fahima, T.; Shen, Q.; Xie, C. Introgression of the Powdery Mildew Resistance Genes Pm60 and Pm60b from Triticum urartu to Common Wheat Using Durum as a ‘Bridge’. Pathogens 2022, 11, 25. https://doi.org/10.3390/pathogens11010025
Zhang Q, Li Y, Li Y, Fahima T, Shen Q, Xie C. Introgression of the Powdery Mildew Resistance Genes Pm60 and Pm60b from Triticum urartu to Common Wheat Using Durum as a ‘Bridge’. Pathogens. 2022; 11(1):25. https://doi.org/10.3390/pathogens11010025
Chicago/Turabian StyleZhang, Qiang, Yinghui Li, Yiwen Li, Tzion Fahima, Qianhua Shen, and Chaojie Xie. 2022. "Introgression of the Powdery Mildew Resistance Genes Pm60 and Pm60b from Triticum urartu to Common Wheat Using Durum as a ‘Bridge’" Pathogens 11, no. 1: 25. https://doi.org/10.3390/pathogens11010025
APA StyleZhang, Q., Li, Y., Li, Y., Fahima, T., Shen, Q., & Xie, C. (2022). Introgression of the Powdery Mildew Resistance Genes Pm60 and Pm60b from Triticum urartu to Common Wheat Using Durum as a ‘Bridge’. Pathogens, 11(1), 25. https://doi.org/10.3390/pathogens11010025