
Discrete Survival Model Analysis of Plasmodium falciparum Response to Artemisinin-Based Combination Therapies among Children in Regions of Varying Malaria Transmission in Cameroon

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Study Site and Setting
2.3. Study Population
2.4. Study Procedure
2.5. Statistical Analysis
3. Results
3.1. Description of the Study Participants and Parameters
3.2. Association between Study Sites and Parasite Density
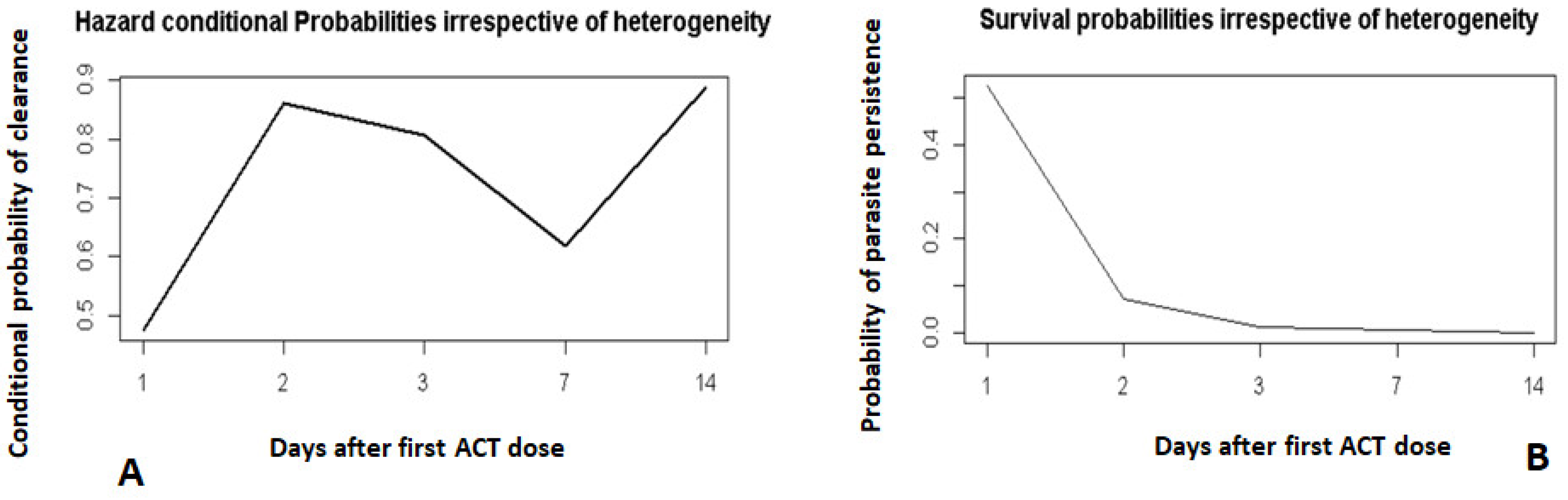
3.3. Discrete-time Survival Model Diagnostics
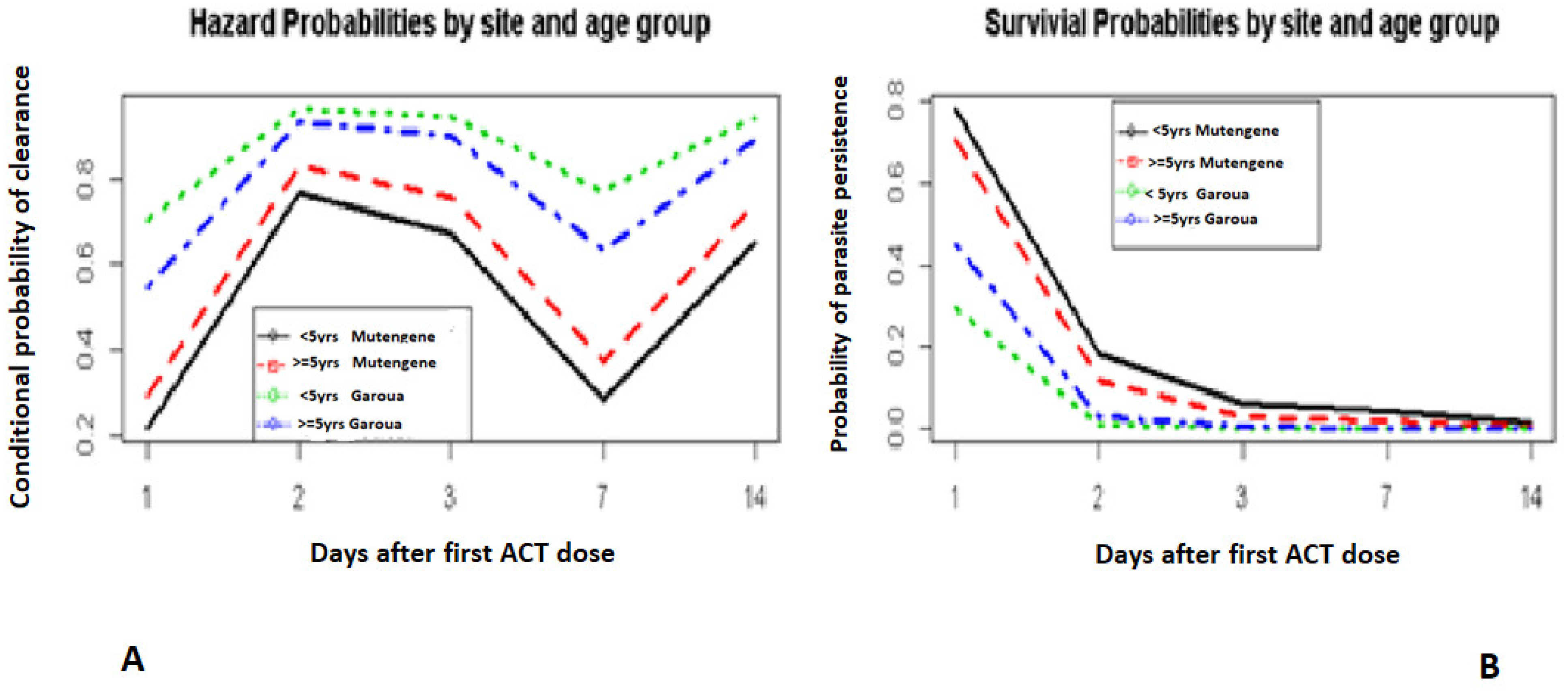
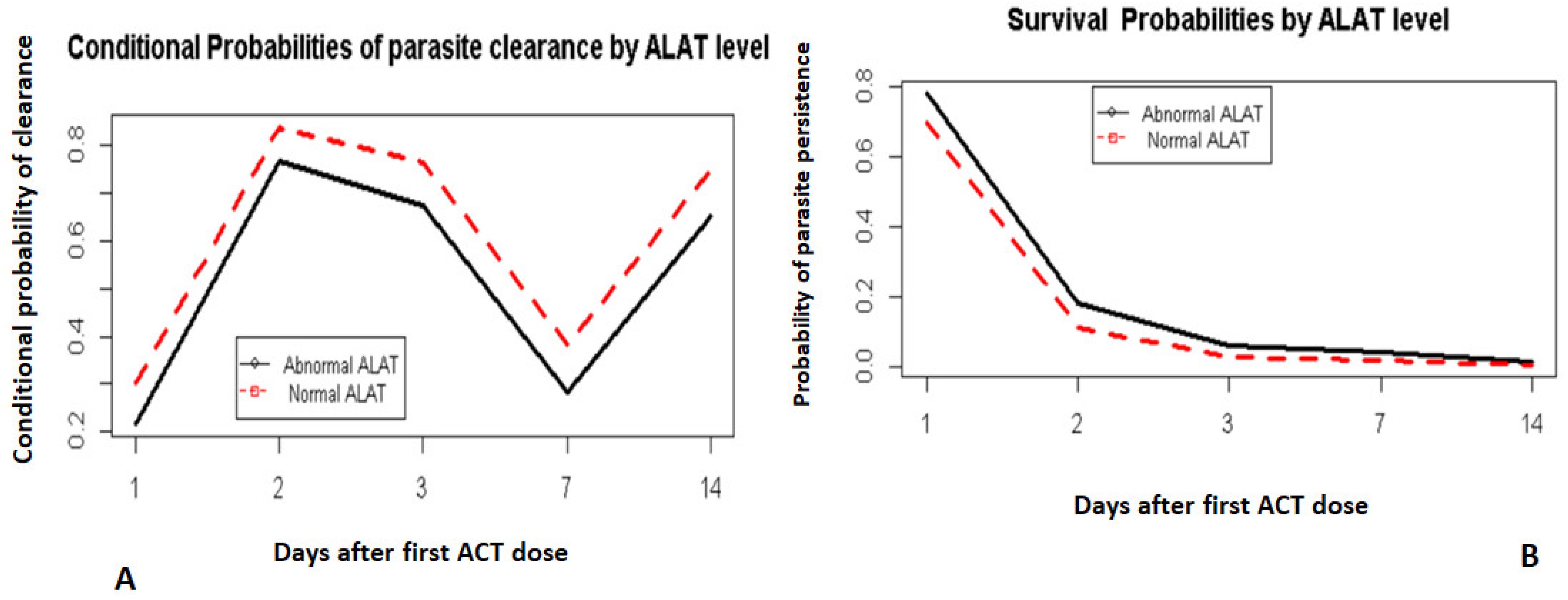
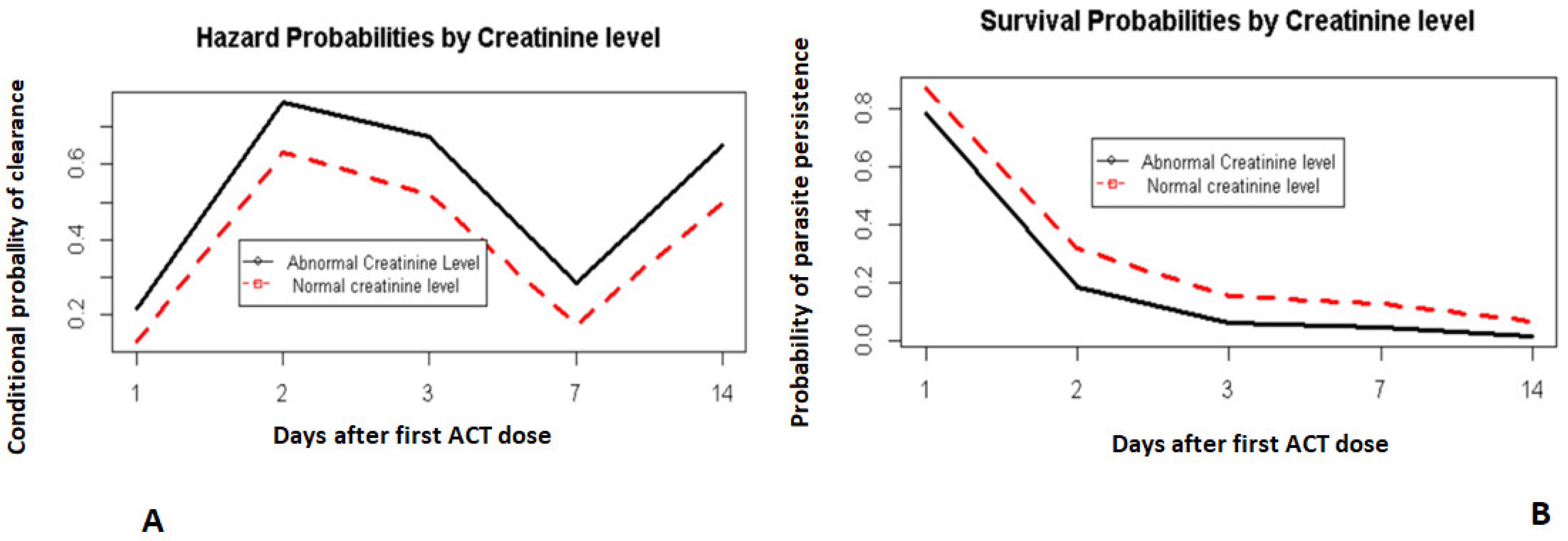
3.4. Survival Model Fitting

4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACT | Artemisinin-based combination therapy |
| ALAT | Alanine aminotransferase |
| AL | Artemether–lumefantrine |
| AS-AQ | Artesunate–amodiaquine |
| DHA-PPQ | Dihydroartemisinin–piperaquine |
| WHO | World Health Organization |
References
- Barnes, K.I.; Durrheim, D.N.; Little, F.; Jackson, A.; Mehta, U.; Allen, E.; Dlamini, S.S.; Tsoka, J.; Bredenkamp, B.; Mthembu, D.J.; et al. Effect of artemether-lumefantrine policy and improved vector control on malaria burden in KwaZulu–Natal, South Africa. PLoS Med. 2005, 2, e330. [Google Scholar] [CrossRef]
- Bhattarai, A.; Ali, A.S.; Kachur, S.P.; Mårtensson, A.; Abbas, A.K.; Khatib, R.; Al-Mafazy, A.-W.; Ramsan, M.; Rotllant, G.; Gerstenmaier, J.F.; et al. Impact of Artemisinin-Based Combination Therapy and Insecticide-Treated Nets on Malaria Burden in Zanzibar. PLoS Med. 2007, 4, e309. [Google Scholar] [CrossRef]
- Wongsrichanalai, C.; Meshnick, S.R. Declining artesunate-mefloquine efficacy against falciparum malaria on the Cambodia-Thailand border. Emerg. Infect. Dis. 2008, 14, 716–719. [Google Scholar] [CrossRef] [PubMed]
- Na-Bangchang, K.; Ruengweerayut, R.; Mahamad, P.; Ruengweerayut, K.; Chaijaroenkul, W. Declining in efficacy of a three-day combination regimen of mefloquine-artesunate in a multi-drug resistance area along the Thai-Myanmar border. Malar. J. 2010, 9, 273. [Google Scholar] [CrossRef] [PubMed]
- Ariey, F.; Witkowski, B.; Amaratunga, C.; Beghain, J.; Langlois, A.-C.; Khim, N.; Kim, S.; Duru, V.; Bouchier, C.; Ma, L.; et al. A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature 2014, 505, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Tacoli, C.; Gai, P.P.; Bayingana, C.; Sifft, K.; Geus, D.; Ndoli, J.; Sendegeya, A.; Gahutu, J.B.; Mockenhaupt, F.P. Artemisinin resistance-associated K13 Polymorphisms of Plasmodium falciparum in Southern Rwanda, 2010–2015. Am. J. Trop. Med. Hyg. 2016, 95, 1090–1093. [Google Scholar] [CrossRef] [PubMed]
- Bwire, G.M.; Ngasala, B.; Mikomangwa, W.P.; Kilonzi, M.; Kamuhabwa, A.A.R. Detection of mutations associated with artemisinin resistance at k13-propeller gene and a near complete return of chloroquine susceptible falciparum malaria in Southeast of Tanzania. Sci. Rep. 2020, 10, 3500. [Google Scholar] [CrossRef]
- Uwimana, A.; Legrand, E.; Stokes, B.H.; Ndikumana, J.-L.M.; Warsame, M.; Umulisa, N.; Ngamije, D.; Munyaneza, T.; Mazarati, J.-B.; Munguti, K.; et al. Emergence and clonal expansion of in vitro artemisinin-resistant Plasmodium falciparum kelch13 R561H mutant parasites in Rwanda. Nat. Med. 2020, 26, 1602–1608. [Google Scholar] [CrossRef]
- Uwimana, A.; Umulisa, N.; Venkatesan, M.; Svigel, S.S.; Zhou, Z.; Munyaneza, T.; Habimana, R.M.; Rucogoza, A.; Moriarty, L.F.; Sandford, R.; et al. Association of Plasmodium falciparum kelch13 R561H genotypes with delayed parasite clearance in Rwanda: An open-label, single-arm, multicentre, therapeutic efficacy study. Lancet Infect. Dis. 2021, 21, 1120–1128. [Google Scholar] [CrossRef]
- Anderson, T.J.C.; Nair, S.; Nkhoma, S.; Williams, J.T.; Imwong, M.; Yi, P.; Socheat, D.; Das, D.; Chotivanich, K.; Day, N.P.J.; et al. High Heritability of Malaria Parasite Clearance Rate Indicates a Genetic Basis for Artemisinin Resistance in Western Cambodia. J. Infect. Dis. 2010, 201, 1326–1330. [Google Scholar] [CrossRef]
- Boni, M.F.; Smith, D.; Laxminarayan, R. Benefits of using multiple first-line therapies against malaria. Proc. Natl. Acad. Sci. USA 2008, 105, 14216–14221. [Google Scholar] [CrossRef] [PubMed]
- Petersen, I.; Eastman, R.; Lanzer, M. Drug-resistant malaria: Molecular mechanisms and implications for public health. FEBS Lett. 2011, 585, 1551–1562. [Google Scholar] [CrossRef]
- Francis, D.; Nsobya, S.L.; Talisuna, A.; Yeka, A.; Kamya, M.R.; Machekano, R.; Dokomajilar, C.; Rosenthal, P.J.; Dorsey, G. Geographic differences in antimalarial drug efficacy in uganda are explained by differences in endemicity and not by known molecular markers of drug resistance. J. Infect. Dis. 2006, 193, 978–986. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Greenhouse, B.; Slater, M.; Njama-Meya, D.; Nzarubara, B.; Maiteki-Sebuguzi, C.; Clark, T.D.; Staedke, S.G.; Kamya, M.R.; Hubbard, A.; Rosenthal, P.J.; et al. Decreasing efficacy of antimalarial combination therapy in Uganda is explained by decreasing host immunity rather than increasing drug resistance. J. Infect. Dis. 2009, 199, 758–765. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Smith, D.L.; Klein, E.Y.; McKenzie, F.E.; Laxminarayan, R. Prospective strategies to delay the evolution of anti-malarial drug resistance: Weighing the uncertainty. Malar. J. 2010, 9, 217. [Google Scholar] [CrossRef]
- Maude, R.J.; Pontavornpinyo, W.; Saralamba, S.; Aguas, R.; Yeung, S.; Dondorp, A.M.; Day, N.P.J.; White, N.J.; White, L.J. The last man standing is the most resistant: Eliminating artemisinin-resistant malaria in Cambodia. Malar. J. 2009, 8, 31. [Google Scholar] [CrossRef]
- Okell, L.C.; Drakeley, C.J.; Bousema, T.; Whitty, C.; Ghani, A. Modelling the Impact of Artemisinin Combination Therapy and Long-Acting Treatments on Malaria Transmission Intensity. PLoS Med. 2008, 5, e226. [Google Scholar] [CrossRef]
- Pongtavornpinyo, W.; Yeung, S.; Hastings, I.M.; Dondorp, A.M.; Day, N.P.; White, N.J. Spread of anti-malarial drug resistance: Mathematical model with implications for ACT drug policies. Malar. J. 2008, 7, 229. [Google Scholar] [CrossRef]
- Sayang, C.; Gausseres, M.; Vernazza-Licht, N.; Malvy, D.; Bley, D.; Millet, P. Treatment of malaria from monotherapy to artemisinin-based combination therapy by health professionals in urban health facilities in Yaoundé, central province, Cameroon. Malar. J. 2009, 8, 176. [Google Scholar] [CrossRef]
- Nji, A.M.; Ali, I.M.; Moyeh, M.N.; Ngongang, E.-O.; Ekollo, A.M.; Chedjou, J.-P.; Ndikum, V.N.; Evehe, M.S.; Froeschl, G.; Heumann, C.; et al. Randomized non-inferiority and safety trial of dihydroartemisin-piperaquine and artesunate-amodiaquine versus artemether-lumefantrine in the treatment of uncomplicated Plasmodium falciparum malaria in Cameroonian children. Malar. J. 2015, 14, 27. [Google Scholar] [CrossRef]
- World Health Organization. Methods for Surveillance of Antimalarial Drug Efficacy; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- World Health Organization. UNICEF/UNDP/World Bank/WHO Special programme for research and training in tropical diseases. In Microscopy for the Detection, Identification and Quantification of Malaria Parasites on Stained Thick and Thin Blood Films in Research Settings (Version 1.0): Procedure: Methods Manual; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Singer, J.D.; Willett, J.B. It’s About Time: Using discrete-time survival analysis to study duration and the timing of events. J. Educ. Stat. 1993, 18, 155–195. [Google Scholar]
- White, N.J. Malaria parasite clearance. Malar. J. 2017, 16, 88. [Google Scholar] [CrossRef]
- Rodriguez-Barraquer, I.; Arinaitwe, E.; Jagannathan, P.; Kamya, M.R.; Rosenthal, P.J.; Rek, J.; Dorsey, G.; Nankabirwa, J.; Staedke, S.G.; Kilama, M.; et al. Quantification of anti-parasite and anti-disease immunity to malaria as a function of age and exposure. Elife 2018, 7, e35832. [Google Scholar] [CrossRef]
- Lopera-Mesa, T.M.; Doumbia, S.; Chiang, S.; Zeituni, A.E.; Konate, D.S.; Doumbouya, M.; Keita, A.S.; Stepniewska, K.; Traore, K.; Diakite, S.A.S.; et al. Plasmodium falciparum Clearance Rates in Response to Artesunate in Malian Children with Malaria: Effect of Acquired Immunity. J. Infect. Dis. 2013, 207, 1655–1663. [Google Scholar] [CrossRef]
- Nhabomba, A.J.; Guinovart, C.; Jiménez, A.; Manaca, M.N.; Quintó, L.; Cisteró, P.; Aguilar, R.; Barbosa, A.; Rodríguez, M.H.; Bassat, Q.; et al. Impact of age of first exposure to Plasmodium falciparum on antibody responses to malaria in children: A randomized, controlled trial in Mozambique. Malar. J. 2014, 13, 121. [Google Scholar] [CrossRef]
- Barua, P.; Beeson, J.G.; Maleta, K.; Ashorn, P.; Rogerson, S.J. The impact of early life exposure to Plasmodium falciparum on the development of naturally acquired immunity to malaria in young Malawian children. Malar. J. 2019, 18, 11. [Google Scholar] [CrossRef]
- Bolad, A.; Farouk, S.E.; Israelsson, E.; Dolo, A.; Doumbo, O.K.; Nebie, I.; Maiga, B.; Kouriba, B.; Luoni, G.; Sirima, B.S.; et al. Distinct Interethnic Differences in immunoglobulin G class/subclass and immunoglobulin m antibody responses to malaria antigens but not in immunoglobulin g responses to non-malarial antigens in sympatric tribes living in West Africa. Scand. J. Immunol. 2005, 61, 380–386. [Google Scholar] [CrossRef]
- Flegg, J.A.; Guérin, P.J.; Nosten, F.; Ashley, E.A.; Phyo, A.P.; Dondorp, A.M.; Fairhurst, R.M.; Socheat, D.; Borrmann, S.; Björkman, A.; et al. Optimal sampling designs for estimation of Plasmodium falciparum clearance rates in patients treated with artemisinin derivatives. Malar. J. 2013, 12, 411. [Google Scholar] [CrossRef]
- Nkhoma, S.C.; Stepniewska, K.; Nair, S.; Phyo, A.P.; McGready, R.; Nosten, F.; Anderson, T.J.C. Genetic evaluation of the performance of malaria parasite clearance rate metrics. J. Infect. Dis. 2013, 208, 346–350. [Google Scholar] [CrossRef]
- Takala-Harrison, S.; Jacob, C.G.; Arze, C.; Cummings, M.P.; Silva, J.C.; Dondorp, A.M.; Fukuda, M.M.; Hien, T.T.; Mayxay, M.; Noedl, H.; et al. Independent emergence of artemisinin resistance mutations among Plasmodium falciparum in Southeast Asia. J. Infect. Dis. 2015, 211, 670–679. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predictors | Model Parameters β (Standard Error) |
|---|---|
| Day 1 | 4.63 (0.43) *** |
| Day 2 | 7.14 (0.48) *** |
| Day 3 | 6.75 (0.65) *** |
| Day 7 | 5.04 (0.63) *** |
| Day 14 | 6.60 (1.16) *** |
| Day 21 | −9.77 (533.41) |
| Sex (male) | −0.10 (0.15) |
| Age group > 5 years | −0.76 (0.40) |
| Site (Mutengene) | 4.05 (1.29) ** |
| AS-AQ | −0.14 (0.36) |
| DHA-PPQ | −0.17 (0.36) |
| Fast metabolizer | 0.03 (0.16) |
| Normal neutrophils level day 0 (%) | −0.13 (0.16) |
| Normal ALAT level day 0 (U/L) | 0.46 (0.22) * |
| Ecotype (Forest) | 1.49 (1.23) |
| Ecotype (Sahel) | −1.26 (1.70) |
| Urbanicity (Urban) | −0.03 (0.26) |
| Weight (kg) | 0.01 (0.02) |
| Temperature (°C) | −0.09 (0.11) |
| Hemoglobin level > 10 g/dL | 0.26 (0.16) |
| Normal creatinine level at day 0 (mg/dL) | −0.64 (0.21) ** |
| Age group > 5 years × AS-AQ | 0.12 (0.41) |
| Age group > 5 years × DHA-PPQ | −0.01 (0.41) |
| Site (Mutengene) × AS-AQ | 0.36 (0.41) |
| Site (Mutengene) × DHA-PPQ | 0.69 (0.42) |
| Age group > 5 years × Site (Mutengene) | 1.00 (0.31) ** |
| Predictors | Odd Ratio Estimate | 95% Confidence Interval |
|---|---|---|
| Site (Mutengene) | 0.12 | (0.07, 0.18) * |
| Normal ALAT level day 0 | 1.58 | (1.15, 2.01) * |
| Normal creatinine level at day 0 | 0.53 | (0.12, 0.67) * |
| Age group > 5 years × Site (Mutengene) | 2.72 | (2.11, 3.33) * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nji, A.M.; Ali, I.M.; Niba, P.T.N.; Marie-Solange, E.; Heumann, C.; Froeschl, G.; Mbacham, W.F. Discrete Survival Model Analysis of Plasmodium falciparum Response to Artemisinin-Based Combination Therapies among Children in Regions of Varying Malaria Transmission in Cameroon. Pathogens 2021, 10, 1106. https://doi.org/10.3390/pathogens10091106
Nji AM, Ali IM, Niba PTN, Marie-Solange E, Heumann C, Froeschl G, Mbacham WF. Discrete Survival Model Analysis of Plasmodium falciparum Response to Artemisinin-Based Combination Therapies among Children in Regions of Varying Malaria Transmission in Cameroon. Pathogens. 2021; 10(9):1106. https://doi.org/10.3390/pathogens10091106
Chicago/Turabian StyleNji, Akindeh M., Innocent M. Ali, Peter Thelma Ngwa Niba, Evehe Marie-Solange, Christian Heumann, Guenter Froeschl, and Wilfred F. Mbacham. 2021. "Discrete Survival Model Analysis of Plasmodium falciparum Response to Artemisinin-Based Combination Therapies among Children in Regions of Varying Malaria Transmission in Cameroon" Pathogens 10, no. 9: 1106. https://doi.org/10.3390/pathogens10091106
APA StyleNji, A. M., Ali, I. M., Niba, P. T. N., Marie-Solange, E., Heumann, C., Froeschl, G., & Mbacham, W. F. (2021). Discrete Survival Model Analysis of Plasmodium falciparum Response to Artemisinin-Based Combination Therapies among Children in Regions of Varying Malaria Transmission in Cameroon. Pathogens, 10(9), 1106. https://doi.org/10.3390/pathogens10091106