
Genetic Analysis, Population Structure, and Characterisation of Multidrug-Resistant Klebsiella pneumoniae from the Al-Hofuf Region of Saudi Arabia

 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Methods
2.1. Bacterial Isolates
2.2. The Extraction of Genomic DNA and PFGE Protocol for Klebsiella pneumoniae
2.3. Analysis of PFGE
2.4. 16s rRNA Sequencing of the Isolates and Analysis
2.5. Detection of Carriage of Antimicrobial Resistance Genes by Multiplex PCR
2.6. Statistical Analysis
3. Results
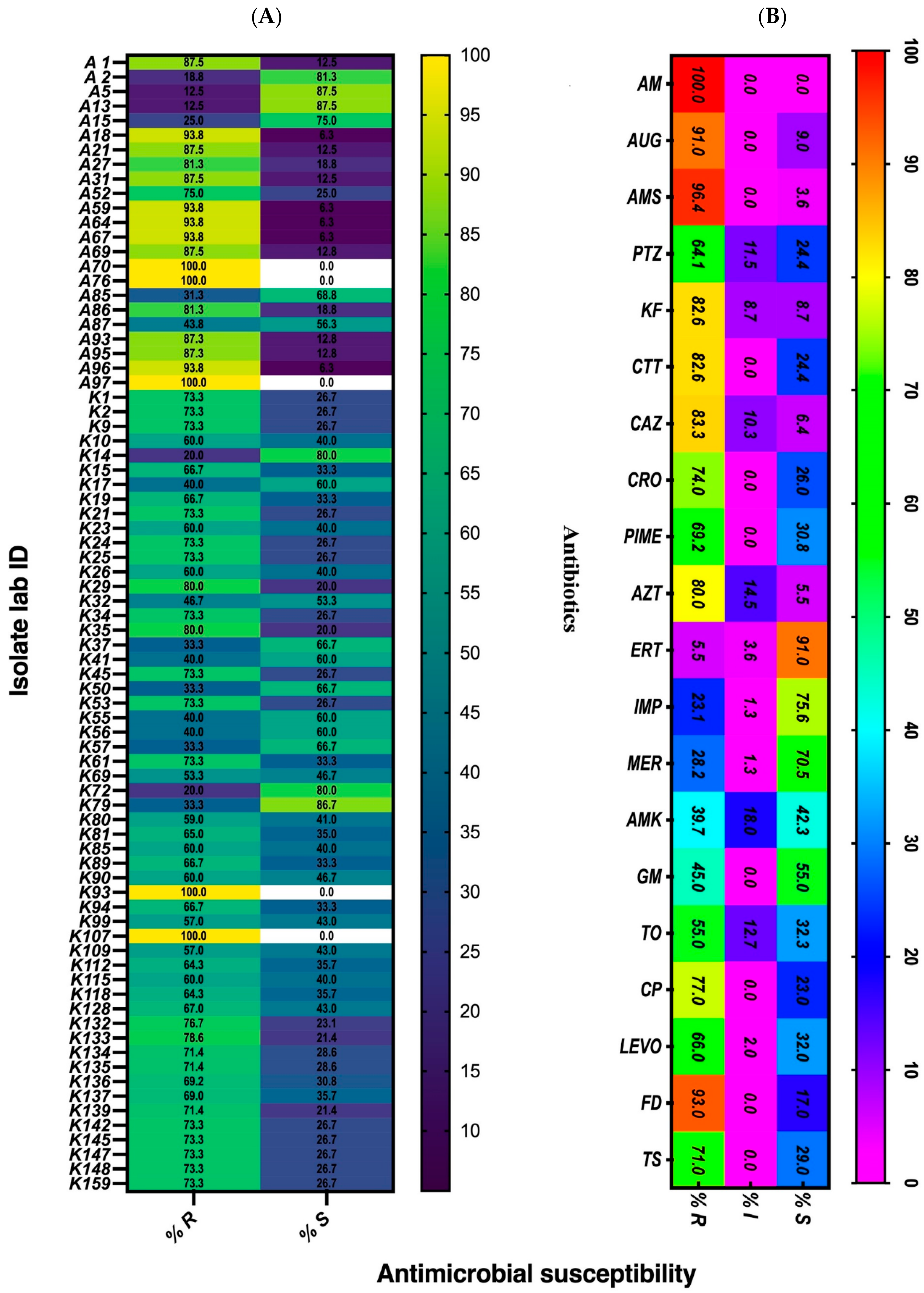
3.1. Antimicrobial Susceptibility Pattern of the Isolates
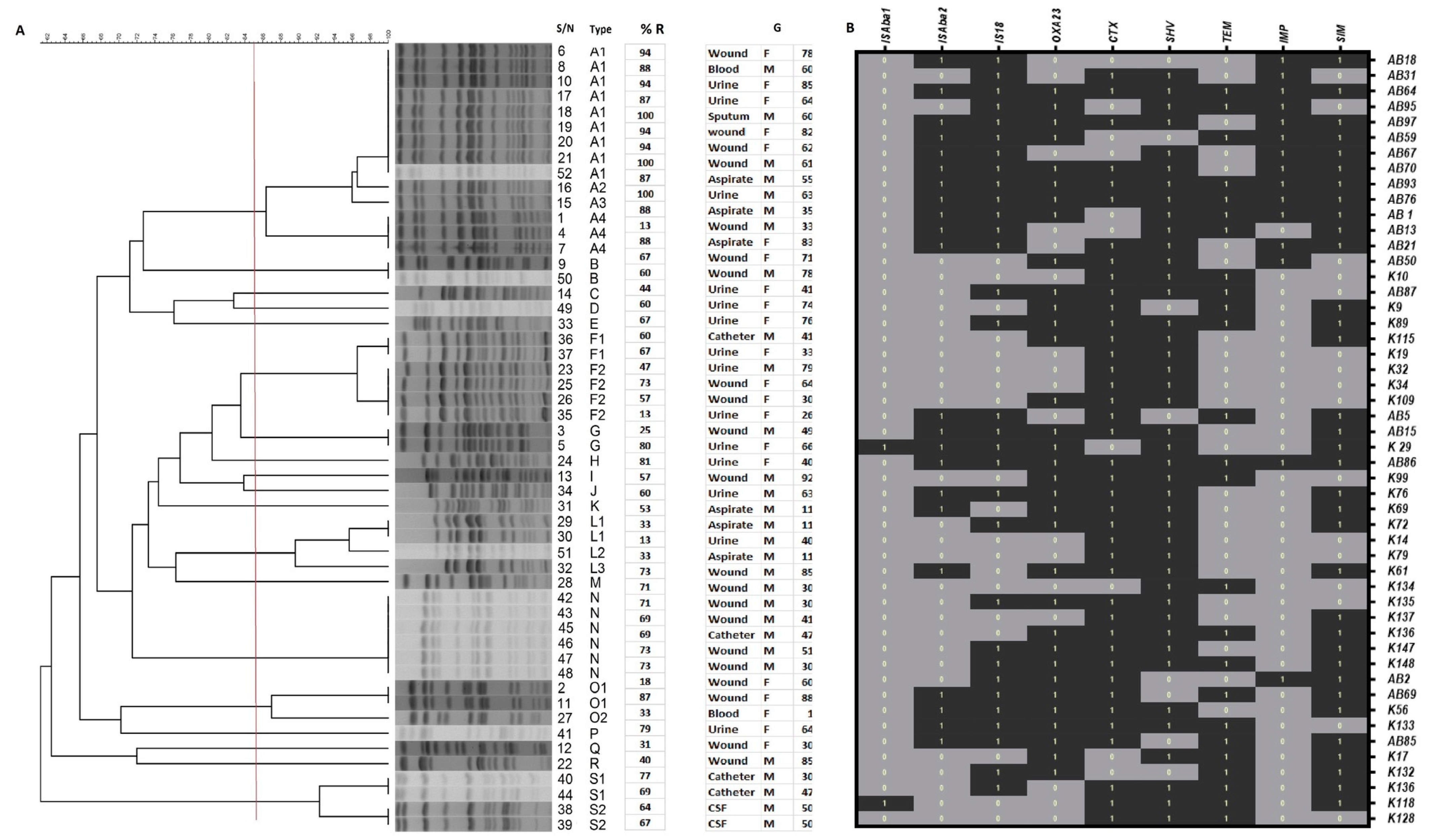
3.2. Genotyping of Klebsiella pneumoniae Isolates by PFGE
3.3. Associated Resistance Genes by Multiplex PCR
3.4. K. pneumoniae 16S rRNA Molecular Genotyping
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ashurst, J.V.; Dawson, A. Klebsiella Pneumonia. In StatPearls Treasure Island (FL); StatPearls Publishing, 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK519004/ (accessed on 8 July 2021).
- Wyres, K.L.; Holt, K.E. Klebsiella pneumoniae Population Genomics and Antimicrobial-Resistant Clones. Trends Microbiol. 2016, 24, 944–956. [Google Scholar] [CrossRef]
- Mędrzycka-Dąbrowska, W.; Lange, S.; Zorena, K.; Dąbrowski, S.; Ozga, D.; Tomaszek, L. Carbapenem-Resistant Klebsiella pneumoniae Infections in ICU COVID-19 Patients—A Scoping Review. J. Clin. Med. 2021, 10, 2067. [Google Scholar] [CrossRef]
- Martin, R.M.; Bachman, M.A. Colonization, Infection, and the Accessory Genome of Klebsiella pneumoniae. Front. Cell. Infect. Microbiol. 2018, 8, 4. [Google Scholar] [CrossRef]
- Alves, A.; Sucena, I.; Ribeiro, C.; Silva, E.; Shiang, T.; Vanzeller, M. Factors associated with colonization and infection by Carbapenem Resistant Enterobacteriaceae. Eur. Respir. J. 2018, 52, PA4702. [Google Scholar]
- Sievert, D.M.; Ricks, P.; Edwards, J.R.; Schneider, A.; Patel, J.; Srinivasan, A.; Kallen, A.; Limbago, B.; Fridkin, S. Antimicrobial-resistant pathogens associated with healthcare-associated infections: Summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009–2010. Infect. Control. Hosp. Epidemiol. 2013, 34, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kotb, S.; Lyman, M.; Ismail, G.; Fattah, M.A.E.; Girgis, S.A.; Etman, A.; Hafez, S.; El-Kholy, J.; Zaki, M.E.S.; Rashed, H.-a.G.; et al. Epidemiology of Carbapenem-resistant Enterobacteriaceae in Egyptian intensive care units using National Healthcare-associated Infections Surveillance Data, 2011–2017. Antimicrob. Resist. Infect. Control. 2020, 9, 2. [Google Scholar] [CrossRef]
- Jones, R.N. Microbial etiologies of hospital-acquired bacterial pneumonia and ventilator-associated bacterial pneumonia. Clin. Infect. Dis. 2010, 51 (Suppl. 1), S81–S87. [Google Scholar] [CrossRef] [PubMed]
- Antibiotic Resistance Threats in the United States. Atlanta, GA: U.S. Department of Health and Human Services, CDC. 2019. Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report-508.pdf (accessed on 24 May 2021).
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; WHO: 1211 Geneva 27-Switzerland 2014. Available online: https://apps.who.int/iris/bitstream/handle/10665/112642/9789241564748_eng.pdf?sequence=1&isAllowed=y (accessed on 24 May 2021).
- Sleiman, A.; Awada, B.; Mocadie, M.; Sherri, N.; Haraoui, L.P.; Baby, V.; Araj, G.F.; Kanj, S.S.; Rizk, N.; Matar, G.M.; et al. An unequivocal superbug: PDR Klebsiella pneumoniae with an arsenal of resistance and virulence factor genes. J. Infect. Dev. Ctries. 2021, 15, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Peleg, A.Y.; Hooper, D.C. Hospital-acquired infections due to gram-negative bacteria. N. Engl. J. Med. 2010, 362, 1804–1813. [Google Scholar] [CrossRef]
- MacVane, S.H. Antimicrobial Resistance in the Intensive Care Unit: A Focus on Gram-Negative Bacterial Infections. J. Intensive Care Med. 2017, 32, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Mayr, F.B.; Yende, S.; Angus, D.C. Epidemiology of severe sepsis. Virulence 2014, 5, 4–11. [Google Scholar] [CrossRef]
- Effah, C.Y.; Sun, T.; Liu, S.; Wu, Y. Klebsiella pneumoniae: An increasing threat to public health. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 1. [Google Scholar] [CrossRef]
- Rønning, T.G.; Aas, C.G.; Støen, R.; Bergh, K.; Afset, J.E.; Holte, M.S.; Radtke, A. Investigation of an outbreak caused by antibiotic-susceptible Klebsiella oxytoca in a neonatal intensive care unit in Norway. Acta Paediatr. 2019, 108, 76–82. [Google Scholar] [CrossRef]
- Tsereteli, M.; Sidamonidze, K.; Tsereteli, D.; Malania, L.; Vashakidze, E. Epidemiology of Carbapenem-resistant Klebsiella pneumoniae in intensive care units of multiprofile hospitals in Tbilisi, Georgia. Georgian Med. News 2018, 280–281, 164–168. Available online: https://pubmed.ncbi.nlm.nih.gov/30204118/#:~:text=expand-,PMID%3A%2030204118,-Abstract (accessed on 16 June 2021).
- Al-Tawfiq, J.A.; Antony, A. Antimicrobial resistance of Klebsiella pneumoniae in a Saudi Arabian hospital: Results of a 6-year surveillance study, 1998–2003. J. Infect. Chemother. 2007, 13, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Al Bshabshe, A.; Al-Hakami, A.; Alshehri, B.; Al-Shahrani, K.A.; Alshehri, A.A.; Al Shahrani, M.B.; Assiry, I.; Joseph, M.R.; Alkahtani, A.M.; Hamid, M.E. Rising Klebsiella pneumoniae Infections and Its Expanding Drug Resistance in the Intensive Care Unit of a Tertiary Healthcare Hospital, Saudi Arabia. Cureus 2020, 12, e10060. [Google Scholar] [CrossRef]
- Alghoribi, M.F.; Binkhamis, K.; Alswaji, A.A.; Alhijji, A.; Alsharidi, A.; Balkhy, H.H.; Doumith, M.; Somily, A. Genomic analysis of the first KPC-producing Klebsiella pneumoniae isolated from a patient in Riyadh: A new public health concern in Saudi Arabia. J. Infect. Public Health 2020, 13, 647–650. [Google Scholar] [CrossRef]
- Al-Zalabani, A.; AlThobyane, O.A.; Alshehri, A.H.; Alrehaili, A.O.; Namankani, M.O.; Aljafri, O.H. Prevalence of Klebsiella pneumoniae Antibiotic Resistance in Medina, Saudi Arabia, 2014–2018. Cureus 2020, 12, e9714. [Google Scholar] [CrossRef] [PubMed]
- Al-Agamy, M.H.; Shibl, A.M.; Tawfik, A.F. Prevalence and molecular characterization of extended-spectrum beta-lactamase-producing Klebsiella pneumoniae in Riyadh, Saudi Arabia. Ann. Saudi Med. 2009, 29, 253–257. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kiratisin, P.; Apisarnthanarak, A.; Laesripa, C.; Saifon, P. Molecular characterization and epidemiology of extended-spectrum-beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae isolates causing health care-associated infection in Thailand, where the CTX-M family is endemic. Antimicrob. Agents Chemother. 2008, 52, 2818–2824. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Price, L.S.; Poirel, L.; Bonomo, R.A.; Schwaber, M.J.; Daikos, G.L.; Cormican, M.; Cornaglia, G.; Garau, J.; Gniadkowski, M.; Hayden, M.K.; et al. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect. Dis. 2013, 13, 785–796. [Google Scholar] [CrossRef]
- Hashemizadeh, Z.; Hosseinzadeh, Z.; Azimzadeh, N.; Motamedifar, M. Dissemination Pattern of Multidrug Resistant Carbapenemase Producing Klebsiella pneumoniae Isolates Using Pulsed-Field Gel Electrophoresis in Southwestern Iran. Infect. Drug Resist. 2020, 13, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Clinical Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Testing for Bacteria That Grows Aerobically, 9th ed.; (M07-A9); Clinical Laboratory Standards Institute: Pittsburgh, PA, USA, 2012. [Google Scholar]
- Kaur, J.; Chopra, S.; Sheevani, M.G. Modified Double Disc Synergy Test to Detect ESBL Production in Urinary Isolates of Escherichia coli and Klebsiella pneumoniae. J. Clin. Diagn. Res. 2013, 7, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Standard Operating Procedure for PulseNet PFGE of Escherichia Coli O157:H7, Escherichia Coli Non-O157 (STEC), Salmonella Serotypes, Shigella Sonnei and Shigella Flexneri; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2013. Available online: www.cdc.gov/pulsenet/pdf/ecoli-shigella-salmonella-pfge-protocol-508c.pdf (accessed on 24 May 2021).
- Ribot, E.M.; Fair, M.A.; Gautom, R.; Cameron, D.N.; Hunter, S.B.; Swaminathan, B.; Barrett, T.J. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 2006, 3, 59–67. [Google Scholar] [CrossRef]
- Koroglu, M.; Ozbek, A.; Demiray, T.; Hafizoglu, T.; Guclu, E.; Altindis, M.; Karabay, O.; Durmaz, R. Investigation of clonal relationships of K. pneumoniae isolates from neonatal intensive care units by PFGE and rep-PCR. J. Infect. Dev. Ctries. 2015, 9, 829–836. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Durmaz, R.; Otlu, B.; Koksal, F.; Hosoglu, S.; Ozturk, R.; Ersoy, Y.; Aktas, E.; Gursoy, N.C.; Caliskan, A. The optimization of a rapid pulsed-field gel electrophoresis protocol for the typing of Acinetobacter baumannii, Escherichia coli and Klebsiella spp. Jpn. J. Infect. Dis. 2009, 62, 372–377. [Google Scholar] [PubMed]
- Zakaria, A.M.; Hassuna, N.A. Modified PFGE protocol for improving type ability of DNA degradation susceptible nosocomial Klebsiella pneumoniae. J. Med. Microbiol. 2019, 68, 1787–1792. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: Criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef] [PubMed]
- Woodford, N.; Ellington, M.J.; Coelho, J.M.; Turton, J.F.; Ward, M.E.; Brown, S.; Amyes, S.G.; Livermore, D.M. Multiplex PCR for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int. J. Antimicrob. Agents 2006, 27, 351–353. [Google Scholar] [CrossRef] [PubMed]
- Ellington, M.J.; Kistler, J.; Livermore, D.M.; Woodford, N. Multiplex PCR for rapid detection of genes encoding acquired metallo-beta-lactamases. J. Antimicrob. Chemother. 2007, 59, 321–322. [Google Scholar] [CrossRef] [PubMed]
- Yum, J.H.; Yi, K.; Lee, H.; Yong, D.; Lee, K.; Kim, J.M.; Rossolini, G.M.; Chong, Y. Molecular characterization of metallo-beta-lactamase-producing Acinetobacter baumannii and Acinetobacter genomospecies 3 from Korea: Identification of two new integrons carrying the bla(VIM-2) gene cassettes. J. Antimicrob. Chemother. 2002, 49, 837–840. [Google Scholar] [CrossRef]
- Poirel, L.; Nordmann, P. Carbapenem resistance in Acinetobacter baumannii: Mechanisms and epidemiology. Clin. Microbiol. Infect. 2006, 12, 826–836. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.; Sharma, M.; Ray, P. Detection of TEM & SHV genes in Escherichia coli & Klebsiella pneumoniae isolates in a tertiary care hospital from India. Indian J. Med. Res. 2010, 132, 332–336. [Google Scholar]
- Sharma, M.; Pathak, S.; Srivastava, P. Prevalence and antibiogram of Extended Spectrum β-Lactamase (ESBL) producing Gram negative bacilli and further molecular characterization of ESBL producing Escherichia coli and Klebsiella spp. J. Clin. Diagn. Res. 2013, 7, 2173–2177. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI supplement M100; Clinical and Laboratory Standards Institute: ittsburgh, PA, USA, 2020; Available online: https://www.nih.org.pk/wp-content/uploads/2021/02/CLSI-2020.pdf (accessed on 1 August 2021).
- Nirwati, H.; Sinanjung, K.; Fahrunissa, F.; Wijaya, F.; Napitupulu, S.; Hati, V.P.; Hakim, M.S.; Meliala, A.; Aman, A.T.; Nuryastuti, T. Biofilm formation and antibiotic resistance of Klebsiella pneumoniae isolated from clinical samples in a tertiary care hospital, Klaten, Indonesia. BMC Proc. 2019, 13, 20. [Google Scholar] [CrossRef] [PubMed]
- Lagha, R.; Ben Abdallah, F.; ALKhammash, A.A.H.; Amor, N.; Hassan, M.M.; Mabrouk, I.; Alhomrani, M.; Gaber, A. Molecular characterization of multidrug resistant Klebsiella pneumoniae clinical isolates recovered from King Abdulaziz Specialist Hospital at Taif City, Saudi Arabia. J. Infect. Public Health 2021, 14, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Azim, N.S.A.; Nofal, M.Y.; AlHarbi, M.A.; Al-Zaban, M.I.; Somily, A.M. Molecular-diversity, Prevalence and Antibiotic Susceptibility of Pathogenic Klebsiella Pneumoniae under Saudi Condition. Pak. J. Biol. Sci. 2019, 22, 174–179. [Google Scholar] [CrossRef]
- Rammaert, B.; Goyet, S.; Beauté, J.; Hem, S.; Te, V.; Try, P.L.; Mayaud, C.; Borand, L.; Buchy, P.; Guillard, B.; et al. Klebsiella pneumoniae related community-acquired acute lower respiratory infections in Cambodia: Clinical characteristics and treatment. BMC Infect. Dis. 2012, 12, 3. [Google Scholar] [CrossRef]
- Pitout, J.D.D.; Nordmann, P.; Poirel, L. Carbapenemase-Producing Klebsiella pneumoniae, a Key Pathogen Set for Global Nosocomial Dominance. Antimicrob. Agents Chemother. 2015, 59, 5873–5884. [Google Scholar] [CrossRef]
- Wasfi, R.; Elkhatib, W.F.; Ashour, H.M. Molecular typing and virulence analysis of multidrug resistant Klebsiella pneumoniae clinical isolates recovered from Egyptian hospitals. Sci. Rep. 2016, 6, 38929. [Google Scholar] [CrossRef]
- Manjula, N.G.; Math, G.C.; Nagshetty, K.; Patil, S.A.; Gaddad, S.M.; Shivannavar, C.T. Antibiotic Susceptibility Pattern of ESβL Producing Klebsiella pneumoniae Isolated from Urine Samples of Pregnant Women in Karnataka. J. Clin. Diagn. Res. 2014, 8, DC08–DC11. [Google Scholar] [CrossRef]
- Cepas, V.; López, V.C.; Muñoz, E.; Rolo, D.; Ardanuy, C.; Martí, S.; Xercavins, M.; Horcajada, J.P.; Bosch, J.; Soto, S.M. Relationship Between Biofilm Formation and Antimicrobial Resistance in Gram-Negative Bacteria. Microb. Drug Resist. 2019, 25, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Akhavan, B.J.; Khanna, N.R.; Vijhani, P. Amoxicillin. In StatPearls [Internet], Treasure Island (FL); StatPearls Publishing: USA, 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK482250/ (accessed on 8 July 2021).
- Ahmad, S.; Al-Juaid, N.F.; Alenzi, F.Q.; Mattar, E.H.; Bakheet, O.E.-S. Prevalence, antibiotic susceptibility pattern and production of extended-spectrum beta-lactamases amongst clinical isolates of Klebsiella pneumoniae at Armed Forces Hospital in Saudi Arabia. J. Coll. Physicians Surg. Pak. 2009, 19, 264–265. [Google Scholar]
- Almaghrabi, R.; Clancy, C.J.; Doi, Y.; Hao, B.; Chen, L.; Shields, R.K.; Press, E.G.; Iovine, N.M.; Townsend, B.M.; Wagener, M.M.; et al. Carbapenem-resistant Klebsiella pneumoniae strains exhibit diversity in aminoglycoside-modifying enzymes, which exert differing effects on plazomicin and other agents. Antimicrob. Agents Chemother. 2014, 58, 4443–4451. [Google Scholar] [CrossRef] [PubMed]
- Doi, Y.; Paterson, D.L. Carbapenemase-producing Enterobacteriaceae. Semin Respir. Crit. Care Med. 2015, 36, 74–84. [Google Scholar] [CrossRef]
- Shahcheraghi, F.; Aslani, M.M.; Mahmoudi, H.; Karimitabar, Z.; Solgi, H.; Bahador, A.; Alikhani, M.Y. Molecular study of carbapenemase genes in clinical isolates of Enterobacteriaceae resistant to carbapenems and determining their clonal relationship using pulsed-field gel electrophoresis. J. Med. Microbiol. 2017, 66, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, E.R.; Aly, S.A.; Halby, H.M.; Ahmed, S.H.; Zakaria, A.M.; El-Asheer, O.M. Epidemiological typing of multidrug-resistant Klebsiella pneumoniae, which causes paediatric ventilator-associated pneumonia in Egypt. J. Med. Microbiol. 2017, 66, 628–634. [Google Scholar] [CrossRef]
- Zheng, R.; Zhang, Q.; Guo, Y.; Feng, Y.; Liu, L.; Zhang, A.; Zhao, Y.; Yang, X.; Xia, X. Outbreak of plasmid-mediated NDM-1-producing Klebsiella pneumoniae ST105 among neonatal patients in Yunnan, China. Ann. Clin. Microbiol. Antimicrob. 2016, 15, 10. [Google Scholar] [CrossRef] [PubMed]
- Medini, D.; Donati, C.; Tettelin, H.; Masignani, V.; Rappuoli, R. The microbial pan-genome. Curr. Opin. Genet. Dev. 2005, 15, 589–594. [Google Scholar] [CrossRef]
- Karlowsky, J.A.; Jones, M.E.; Draghi, D.C.; Thornsberry, C.; Sahm, D.F.; Volturo, G.A. Prevalence and antimicrobial susceptibilities of bacteria isolated from blood cultures of hospitalized patients in the United States in 2002. Ann. Clin. Microbiol. Antimicrob. 2004, 3, 7. [Google Scholar] [CrossRef]
- Xercavins, M.; Jiménez, E.; Padilla, E.; Riera, M.; Freixas, N.; Boix-Palop, L.; Pérez, J.; Calbo, E. High clonal diversity of ESBL-producing Klebsiella pneumoniae isolates from clinical samples in a non-outbreak situation. A cohort study. Antimicrob. Resist. Infect. Control 2020, 9, 5. [Google Scholar] [CrossRef]
- Azab, K.S.M.; Abdel-Rahman, M.A.; El-Sheikh, H.H.; Azab, E.; Gobouri, A.A.; Farag, M.M.S. Distribution of Extended-Spectrum β-Lactamase (ESBL)-Encoding Genes among Multidrug-Resistant Gram-Negative Pathogens Collected from Three Different Countries. Antibiotics 2021, 10, 247. [Google Scholar] [CrossRef]
- Sheppard, A.E.; Stoesser, N.; Wilson, D.J.; Sebra, R.; Kasarskis, A.; Anson, L.W.; Giess, A.; Pankhurst, L.J.; Vaughan, A.; Grim, C.J.; et al. Nested Russian Doll-Like Genetic Mobility Drives Rapid Dissemination of the Carbapenem Resistance Gene bla KPC. Antimicrob. Agents Chemother. 2016, 60, 3767–3778. [Google Scholar] [CrossRef]
- Mshana, S.E.; Fritzenwanker, M.; Falgenhauer, L.; Domann, E.; Hain, T.; Chakraborty, T.; Imirzalioglu, C. Molecular epidemiology and characterization of an outbreak causing Klebsiella pneumoniae clone carrying chromosomally located bla(CTX-M-15) at a German University-Hospital. BMC Microbiol. 2015, 15, 122. [Google Scholar] [CrossRef] [PubMed]
- Zowawi, H.M.; Forde, B.M.; Alfaresi, M.; Alzarouni, A.; Farahat, Y.; Chong, T.-M.; Yin, W.-F.; Chan, K.G.; Li, J.; Schembri, M.A.; et al. Stepwise evolution of pandrug-resistance in Klebsiella pneumoniae. Sci. Rep. 2015, 5, 15082. [Google Scholar] [CrossRef]
- Villalón, P.; Valdezate, S.; Medina-Pascual, M.J.; Carrasco, G.; Vindel, A.; Saez-Nieto, J.A. Epidemiology of the Acinetobacter-derived cephalosporinase, carbapenem-hydrolysing oxacillinase and metallo-β-lactamase genes, and of common insertion sequences, in epidemic clones of Acinetobacter baumannii from Spain. J. Antimicrob. Chemother. 2013, 68, 550–553. [Google Scholar] [CrossRef] [PubMed]
- Sabbagh, P.; Ferdosi-Shahandashti, A.; Rajabnia, M.; Maali, A.; Shahandashti, F.E. Investigating Class I Integron and Antimicrobial Resistance Profile of Klebsiella pneumonia isolates in Babol, North of Iran. J. Med Microbiol. Infect. Dis. 2020, 8, 24–28. Available online: http://jommid.pasteur.ac.ir/article-1-242-en.html (accessed on 23 June 2021). [CrossRef]
- Adams, M.D.; Bishop, B.; Wright, M.S. Quantitative assessment of insertion sequence impact on bacterial genome architecture. Microb. Genom. 2016, 2, e000062. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Bonnin, R.A.; Boulanger, A.; Schrenzel, J.; Kaase, M.; Nordmann, P. Tn125-related acquisition of blaNDM-like genes in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2012, 56, 1087–1089. [Google Scholar] [CrossRef]
- Poirel, L.; Bonnin, R.A.; Nordmann, P. Analysis of the resistome of a multidrug-resistant NDM-1-producing Escherichia coli strain by high-throughput genome sequencing. Antimicrob. Agents Chemother. 2011, 55, 4224–4229. [Google Scholar] [CrossRef]
- Bonnin, R.A.; Poirel, L.; Nordmann, P. A novel and hybrid composite transposon at the origin of acquisition of bla(RTG-5) in Acinetobacter baumannii. Int. J. Antimicrob. Agents 2012, 40, 257–259. [Google Scholar] [CrossRef]
- Akya, A.; Elahi, A.; Chegenelorestani, R.; Rezaee, M. Dissemination of Multidrug-Resistant, Class I and II Integrons and Molecular Typing of CTX-M-producing Klebsiella pneumoniae. Int. J. Appl. Basic Med. Res. 2018, 8, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Ashayeri-Panah, M.; Feizabadi, M.M.; Eftekhar, F. Correlation of Multi-drug Resistance, Integron and blaESBL Gene Carriage with Genetic Fingerprints of Extended-Spectrum β-Lactamase Producing Klebsiella pneumoniae. Jundishapur. J. Microbiol. 2014, 7, e8747. [Google Scholar] [CrossRef] [PubMed]
- Al-Marzooq, F.; Yusof, M.Y.M.; Tay, S.T. Molecular analysis of ciprofloxacin resistance mechanisms in Malaysian ESBL-producing Klebsiella pneumoniae isolates and development of mismatch amplification mutation assays (MAMA) for rapid detection of gyrA and parC mutations. BioMed Res. Int. 2014, 2014, 601630. [Google Scholar] [CrossRef] [PubMed]
- Mshana, S.E.; Hain, T.; Domann, E.; Lyamuya, E.F.; Chakraborty, T.; Imirzalioglu, C. Predominance of Klebsiella pneumoniae ST14 carrying CTX-M-15 causing neonatal sepsis in Tanzania. BMC Infect. Dis. 2013, 13, 466. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Name of Primer | Nucleotide Sequence | Amplicon Size (bp) | Reference |
|---|---|---|---|
| OXA 23-F | GATCGGATTGGAGAACCAGA | 501 | [34] |
| OXA 23-R | ATTCTTGACCGCATTTCCAT | ||
| SIM-F | TACAAGGGATTCGGCATCG | 570 | [35] |
| SIM-R | TAATGGCCTGTTCCCATGTG | ||
| IMP-F | CATGGTTTGGTGGTTCTTGT | 616 | [36] |
| IMP-R | ATAATTTGGCGGACTTTGGC | ||
| ISAba1-F | GTGCTTTGCGCTCATCATGC | 430 | [37] |
| ISAba1-R | CATGTAAACCAATGCTCACC | ||
| ISAba2-F | AATCCGAGATAGAGCGGTTC | 1100 | |
| ISAba2-R | TGACACATAACCTAGTGCAC | ||
| ISAba3-F | CAATCAAATGTCCAACCTGC | 403 | |
| ISAba3-R | CGTTTACCCCAAACATAAGC | ||
| IS18-F | CACCCAACTTTCTCAAGATG | 925 | |
| IS18-R | ACCAGCCATAACTTCACTCG | ||
| CTX-F | GACGATGTCACTGGCTGAGC | 500 | [38] |
| CTX-R | AGCCGCCGACGCTAATACA | ||
| SHV-F | AGGATTGACTGCCTTTTTG | 392 | [39] |
| SHV-R | ATTTGCTGATTTCGCTCG | ||
| TEM-F | ATCAGCAATAAACCAGC | 516 | |
| TEM-R | CCCCGAAGAACGTTTTC |
| Lab. ID | Year | Ward Isolation | Antimicrobial Resistance Pattern | ||
|---|---|---|---|---|---|
| AB1 | 2019 | WD | XDR | CRE | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Amk, Cp, Fd, Ts. |
| AB2 | 2019 | OncW | MDR | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Amk, Fd. | |
| AB5 | 2019 | ER | SS | Am, Aug. | |
| AB13 | 2019 | ENT | SS | Am, Fd. | |
| AB15 | 2019 | OncW | MDR | Am, Cp, Ts. | |
| AB18 | 2019 | WD | XDR | CRE | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Amk, Gm, Cp, Fd, Ts. |
| AB21 | 2019 | ER | XDR | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Amk, Gm, Cp, Fd, Ts. | |
| AB31 | 2019 | WD | XDR | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Amk, Gm, Cp, Fd, Ts. | |
| AB52 | 2019 | WD | XDR | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Cp, Fd, Ts. | |
| AB64 | 2019 | WD | XDR | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Amk, Gm, Cp, Fd, Ts. | |
| AB69 | 2019 | OW | XDR | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Amk, Gm, Cp, Fd, Ts. | |
| AB85 | 2019 | WD | MDR | MDR | Am, Aug, Kf, Ctt, Fd. |
| AB86 | 2019 | OncW | XDR | CRE | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Amk, Cp, Fd, Ts |
| AB87 | 2019 | WD | MDR | MDR | Am, Aug, Kf, Caz, Ctt, Fd, Fs. |
| AB76 | 2019 | WD | XDR | CRE | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Amk, Gm, Cp, Fd, Ts. |
| AB93 | 2019 | ICU | XDR | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Amk, Gm, Cp, Fd, Ts. | |
| AB95 | 2019 | Neuro | XDR | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Mer, Amk, Cp, Fd, Ts. | |
| AB97 | 2019 | ICU | XDR | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Amk, Gm, Cp, Tig, Fd, Ts. | |
| AB59 | 2019 | ICU | XDR | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Amk, Gm, Cp, Fd, Ts. | |
| AB67 | 2019 | WD | XDR | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Amk, Gm, Cp, Fd, Ts. | |
| AB70 | 2019 | WD | XDR | Am, Aug, Ptz, Kf, Ctt, Caz, Cro, Pime, Imp, Mer, Amk, Gm, Cp, Tig, Fd, Ts. | |
| K17 | 2017 | WD | MDR | ESBL ESBL | Ams, Ptz, Caz, Pime, Min. |
| K19 | 2017 | WD | MDR | Ams, Ptz, Caz, Pime, Min, Levo, Ts. | |
| K29 | 2017 | ICU | MDR | Ams, Ptz, Caz, Pime, Azt, Gm, To, Cp, Levo, Ts, Min. | |
| K32 | 2017 | OPD | MDR | Ams, Ptz, Caz, Pime, Azt, Cp, Ts. | |
| K34 | 2017 | ICU | MDR | Ams, Ptz, Caz, Pime, Azt, Gm, To, Cp, Levo, Ts, Min. | |
| K56 | 2017 | WD | MDR | Ams, Ptz, Caz, Pime, Azt, Gm, To, Cp, Levo, Ts, Min. | |
| K61 | 2018 | ER | MDR | Ams, Ptz, Caz, Pime, Azt, Gm, To, Cp, Levo, Ts. | |
| K69 | 2018 | WD | MDR | Ams, Ptz, Caz, Pime, Azt, Cp, Levo, Ts, Min. | |
| K72 | 2018 | ICU | MDR | Caz, Pime, Azt, Cp, Levo, Ts. | |
| K76 | 2018 | OPD | MDR | Caz, Pime, Azt, Cp, Ts. | |
| K79 | 2018 | ICU | MDR | Kf, Caz, Cxm, Fd, Pime, | |
| K89 | 2018 | OPD | MDR | Ams, Ptz, Caz, Pime, Cxm, Fd, Ts, Aug, Cp. | |
| K99 | 2018 | ICU | MDR | Ams, Ptz, Caz, Pime, Cxm, Fd, Ts, Aug, Cp, Gm. | |
| K109 | 2018 | WD | MDR | Ams, Ptz, Caz, Pime, Cxm, Fd, Ts, Aug, Cp, | |
| K112 | 2018 | ICU | MDR | Ams, Ptz, Caz, Pime, Cxm, Fd, Ts, Aug, Cp, | |
| K115 | 2018 | ICU | MDR | Ams, Ptz, Caz, Pime, Cxm, Fd, Ts, Aug, Cp, | |
| K118 | 2018 | ICU | MDR | Ams, Ptz, Caz, Pime, Cxm, Fd, Aug, Cp. | |
| K128 | 2018 | ICU | MDR | Ams, Ptz, Caz, Pime, Cxm, Fd, Aug, Cp. | |
| K132 | 2018 | ICU | MDR | Ams, Ptz, Caz, Pime, Cxm, Fd, Aug, Cp, Ts, Gm. | |
| K133 | 2018 | WD | MDR | Ams, Ptz, Caz, Pime, Cxm, Fd, Aug, Cp, Ts, Gm. | |
| K134 | 2018 | WD | MDR | Ams, Ptz, Caz, Pime, Cxm, Fd, Aug, Cp, Ts, Gm. | |
| K135 | 2018 | WD | MDR | Ams, Ptz, Caz, Pime, Cxm, Fd, Aug, Cp, Ts, Gm. | |
| K136 | 2018 | ICU | MDR | Ams, Nor, Caz, Cp, Gm, Ts, Pime. | |
| K137 | 2018 | ICU | MDR | Ams, Ptz, Cxm, Caz, Pime, Cxm, Fd, Aug, Cp, Ts. | |
| K139 | 2018 | WD | MDR | Ams, Ptz, Cxm, Caz, Pime, Cxm, Fd, Aug, Cp, Ts, Gm. | |
| K147 | 2018 | ICU | MDR | Ams, Ptz, Caz, Pime, Azt, Gm, To, Cp, Levo, Min. | |
| K148 | 2018 | WD | MDR | Ams, Ptz, Caz, Pime, Azt, Gm, To, Cp, Levo, Min. | |
| K9 | 2017 | OPD | MDR | Ams, Ptz, Caz, Pime, Azt, Gm, To, Cp, Levo, Ts. | |
| K10 | 2017 | WD | MDR | Ams, Ptz, Caz, Pime, Azt, Gm, To, Cp, Levo, Min. | |
| K14 | 2017 | WD | MDR | Ams, Ptz, Caz, Pime, Min. | |
| K1 | 2017 | OPD | XDR | Ams, Ptz, Caz, Pime, Azt, To, Cp, Levo, Cst, Ts. | |
| Resistance-Determining Agents | Genotype | Number of Isolates (%) |
|---|---|---|
| OXA carbapenemases | OXA-23 | 34 (67) |
| Absence of OXA-23 and OXA-40 | 17 (33) | |
| Metallo-beta-lactamase (MBLs) | SIM | 22 (43) |
| IMP | 3 (6) | |
| Co-existence of SIM and IMP | 12 (23.5) | |
| Absence of SIM and IMP | 14 (27.5) | |
| Overall carriage/absence of MBLs | 37 (72.5)/14 (27.5) | |
| Insertion sequences | ISAba1 | 1 (2) |
| ISAba2 | 1 (2) | |
| IS18 | 11 (21.6) | |
| Co-existence of ISAba1 and ISAba2 | 1 (2) | |
| Co-existence of ISAba1 and IS18 | 0 (0) | |
| Co-existence of ISAba2 and IS18 | 20 (39) | |
| Co-existence of ISAba1, ISAba2, and IS18 | 1 (2) | |
| Absence of ISAba1, ISAba2, and IS18 | 16 (31.4) | |
| Overall carriage/absence of IS | 35 (69)/16 (31) | |
| Extended-spectrum beta lactamase (ESBL) | TEM | 2 (4) |
| CTX | 1 (2) | |
| SHV | 2 (4) | |
| Co-existence of TEM and SHV | 5 (9.8) | |
| Co-existence of TEM and CTX | 4 (7.8) | |
| Co-existence of SHV and CTX | 22 (43) | |
| Co-existence of TEM, SHV, and CTX | 14 (27) | |
| Absence of TEM, SHV, and CTX | 1 (2) | |
| Overall carriage/absence of ESBL | 50 (98)/1 (2) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badger-Emeka, L.I.; Al-Sultan, A.A.; Bohol, M.F.F.; Al-Anazi, M.R.; Al-Qahtani, A.A. Genetic Analysis, Population Structure, and Characterisation of Multidrug-Resistant Klebsiella pneumoniae from the Al-Hofuf Region of Saudi Arabia. Pathogens 2021, 10, 1097. https://doi.org/10.3390/pathogens10091097
Badger-Emeka LI, Al-Sultan AA, Bohol MFF, Al-Anazi MR, Al-Qahtani AA. Genetic Analysis, Population Structure, and Characterisation of Multidrug-Resistant Klebsiella pneumoniae from the Al-Hofuf Region of Saudi Arabia. Pathogens. 2021; 10(9):1097. https://doi.org/10.3390/pathogens10091097
Chicago/Turabian StyleBadger-Emeka, Lorina I., Abdulrahman A. Al-Sultan, Marie Fe F. Bohol, Mashael R. Al-Anazi, and Ahmed A. Al-Qahtani. 2021. "Genetic Analysis, Population Structure, and Characterisation of Multidrug-Resistant Klebsiella pneumoniae from the Al-Hofuf Region of Saudi Arabia" Pathogens 10, no. 9: 1097. https://doi.org/10.3390/pathogens10091097
APA StyleBadger-Emeka, L. I., Al-Sultan, A. A., Bohol, M. F. F., Al-Anazi, M. R., & Al-Qahtani, A. A. (2021). Genetic Analysis, Population Structure, and Characterisation of Multidrug-Resistant Klebsiella pneumoniae from the Al-Hofuf Region of Saudi Arabia. Pathogens, 10(9), 1097. https://doi.org/10.3390/pathogens10091097