
Proteomics of Gnathostomiasis: A Way Forward for Diagnosis and Treatment Development


Abstract
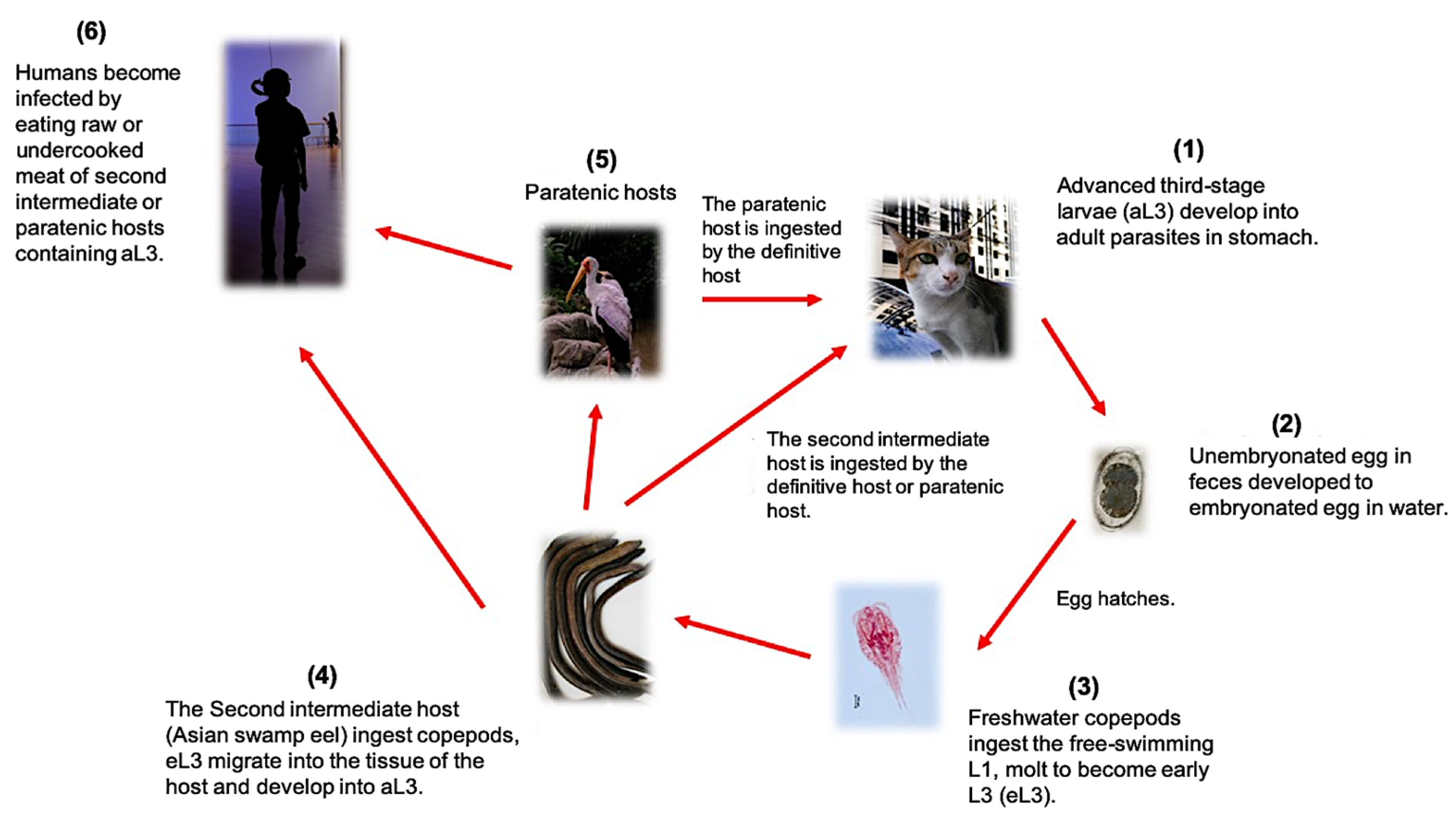
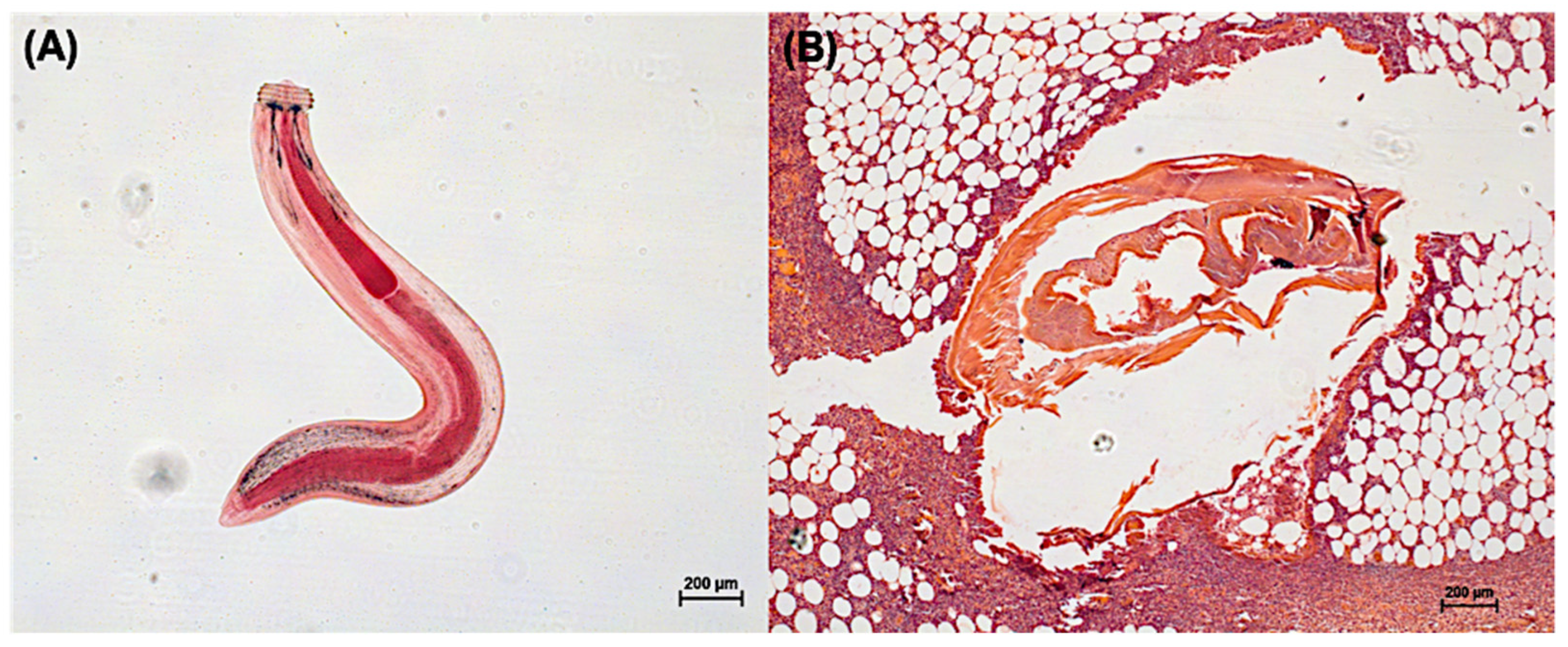
1. Biology and Distribution of Gnathostoma
2. Gnathostomiasis as Public Health and Travel Medicine Problem
3. Clinical Manifestations of Gnathostoma Larva Migration
4. Treatment, Prevention, and Diagnosis of Gnathostomiasis
5. Immunodiagnosis Is Appropriate Method for Diagnosis of Gnathostomiasis
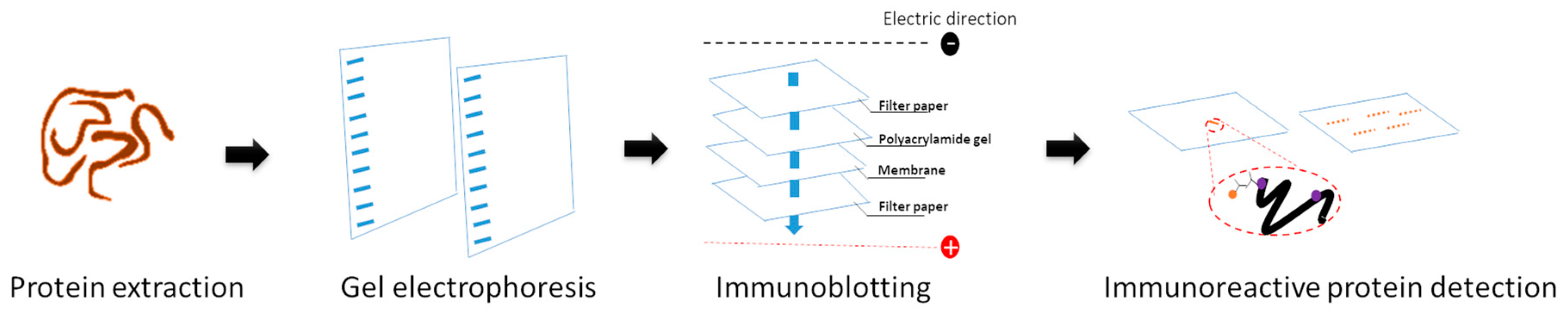
6. Proteomics for Identifying Immunodiagnostic Candidates and Drug Targets of G. spinigerum
7. Future Perspectives of Proteomics in Improving Diagnosis and Treatment of Gnathostomiasis
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Herman, J.S.; Chiodini, P.L. Gnathostomiasis, another emerging imported disease. Clin. Microbiol. Rev. 2009, 22, 484–492. [Google Scholar] [CrossRef]
- Moore, D.A.; McCroddan, J.; Dekumyoy, P.; Chiodni, P.L. Gnathostomiasis: An emerging imported disease. Emerg. Infect. Dis. 2003, 9, 647–650. [Google Scholar] [CrossRef] [PubMed]
- Chaicumpa, W. Immunodiagnosis of gnathostomiasis. Siriraj Med. J. 2010, 62, 79–83. [Google Scholar]
- Diaz, J.H. Gnathostomiasis: An emerging infection of raw fish consumers in Gnathostoma nematode-endemic and nonendemic countries. J. Travel Med. 2015, 22, 318–324. [Google Scholar] [CrossRef]
- Ontranto, D.; Deplazes, P. Zoonotic nematodes of wild carnivores. Int. J. Parasitol. Parasites Wildl. 2019, 9, 370–383. [Google Scholar] [CrossRef]
- Almeyda-Artigas, R.J.; Bargues, M.D.; Mas-Coma, S. ITS-2rDNA Sequencing of Gnathostoma species (Nematoda) and elucidation of the species causing human gnathostomiasis in the Americas. J. Parasitol. 2000, 86, 537–544. [Google Scholar] [CrossRef]
- Katchanov, J.; Sawanyawisuth, K.; Chotmongkol, V.; Nawa, Y. Neurognathostomiasis, a neglected parasitosis of the central nervous system. Emerg. Infect. Dis. 2011, 17, 1174–1180. [Google Scholar] [CrossRef]
- Visudhipan, P.; Chiemchanya, S.; Somburanasin, R.; Dheandhano, D. Causes of spontaneous subarachnoid hemorrhage in Thai infants and children. A study of 56 patients. J. Neurosurg. 1980, 53, 185–187. [Google Scholar] [CrossRef]
- Chitanondh, H.; Rosen, L. Fatal Eosinophilic Encephalomyelitis Caused by the Nematode Gnathostoma Spinigerum. Am. J. Trop. Med. Hyg. 1967, 16, 638–645. [Google Scholar] [CrossRef]
- Boongird, P.; Phuapradit, P.; Siridej, N.; Chirachariyavej, T.; Chuachirun, S.; Vejjajiva, A. Neurological manifestations of gnathostomiasis. J. Neurol. Sci. 1977, 31, 279–291. [Google Scholar] [CrossRef]
- Nopparatana, C.; Chaicumpa, W.; Tapchaisri, P.; Setasuban, P.; Ruangkunaporn, Y. Towards a suitable antigen for diagnosis of Gnathostoma spinigerum infection. Int. J. Parasitol. 1992, 22, 1151–1156. [Google Scholar] [CrossRef]
- Laummaunwai, P.; Intapan, P.M.; Wongkham, C.; Lulitanond, V.; Tayapiwatana, C.; Maleewong, W. Gnathostoma spinigerum: Molecular cloning, expression and characterization of the cyclophilin protein. Exp. Parasitol. 2010, 126, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Daengsyang, S. Human gnathostomiasis in Siam with reference to the method of prevention. J. Parasitol. 1949, 35, 116–121. [Google Scholar] [CrossRef]
- Rusnak, J.M.; Lucey, D.R. Clinical Gnathostomiasis: Case Report and Review of the English-Language Literature. Clin. Infect. Dis. 1993, 16, 33–50. [Google Scholar] [CrossRef]
- Dow, C.; Chiodini, P.; Haines, A.; Michelson, S. Human gnathostomiasis. J. Infect. 1988, 17, 147–149. [Google Scholar] [CrossRef]
- Nagler, A.; Pollack, S.; Hassoun, G.; Kerner, H.; Barzilai, D.; Lengy, J. Human pleuropulmonary gnathosomiasis: A case report from Israel. Isr. J. Med. Sci. 1983, 19, 834–837. [Google Scholar]
- Parola, P.; Bordman, G.; Brouqui, P.; Delmont, J. Eosinopilic pleural effusion in gnathostomiasis. Emerg. Infect. Dis. 2004, 10, 1690–1691. [Google Scholar] [CrossRef] [PubMed]
- Biswas, J.; Gopal, L.; Sharma, T.; Badrinath, S.S. Intraocular Gntathostoma spinigerum. Clinicopathologic study of two cases with review of literature. Retina 1994, 14, 438–444. [Google Scholar] [CrossRef]
- Punyagupta, S.; Bunnag, T.; Juttijudata, P. Eosinophilic meningitis in Thailand. Clinical and epidemiological characteristics of 162 patients with myeloencaphalitis probably caused by Gnathostoma spinigerum. J. Neurol. Sci. 1990, 96, 241–256. [Google Scholar] [CrossRef]
- Nontasut, P.; Bussaratid, V.; Chullawichit, S.; Charoensook, N.; Visetsuk, K. Comparison of ivermectin abd Albendazole treatment for Gnathostomiasis. Southeast Asian J. Trop. Med. Public Health 2006, 43, 433–440. [Google Scholar]
- Kraivichian, K.; Yentakam, S.; Nuchprayoon, S.; Sitichalernchai, P.; Chaicumpa, W. Treatment of cutaneous gnathostomiasis with ivermectin. Am. J. Trop. Med. Hyg. 2004, 71, 623–628. [Google Scholar] [CrossRef]
- Kraivichian, P.; Kulkumthorn, M.; Yingyourd, P.; Akarabovorn, P.; Paireepai, C.-C. Albendazole for the treatment of human gnathostomiasis. Trans. R. Soc. Trop. Med. Hyg. 1992, 86, 418–421. [Google Scholar] [CrossRef]
- Caumes, E.; Carriere, J.; Datry, A.; Gaxotte, P.; Danis, M.; Gentilini, M. A randomized trial of ivermectin versus albendazole for the treatment of cutaneous larva migrants. Am. J. Trop. Med. Hyg. 1993, 49, 641–644. [Google Scholar] [CrossRef]
- Sorvillo, F.J. Food-borne parasitic zoonoses: Fish and plant-borne parasites (World class parasites). Emerg. Infect. Dis. 2008, 14, 1503. [Google Scholar] [CrossRef]
- Hale, D.C.; Blumberg, L.; Frean, J. Case report: Gnathostomiasis in two travelers to Zambia. Am. J. Trop. Med. Hyg. 2003, 68, 707–709. [Google Scholar] [CrossRef]
- Wang, C.H.; Gao, S.F.; Guo, Y.P. Diagnostic significance of eosinophilia of the cerebrospinal fluid in cerebral cysticercosis. Chin. Med. J. 1993, 106, 282–284. [Google Scholar]
- Zambrano-Zaragoza, J.F.; Durán-Avelar, M.D.J.; Vibanco-Pérez, N.; Messina-Robles, M. Characterization of the Humoral Immune Response against Gnathostoma binucleatum in Patients Clinically Diagnosed with Gnathostomiasis. Am. J. Trop. Med. Hyg. 2012, 86, 988–992. [Google Scholar] [CrossRef]
- Diaz-Camacho, S.P.; Zazueta-Ramos, M.; Ponce-Torrecillas, E.; Osuna-Ramirez, I.; Castro-Velasquez, R.; Flores-Gaxiola, A.; Baquera-Heredia, J.; Willms, K.; Akahane, H.; Ogata, K.; et al. Clinical manifestations and immunodiagnosis of gnathostomiasis in Culiacan, Mexico. Am. J. Trop. Med. Hyg. 1998, 59, 908–915. [Google Scholar] [CrossRef]
- Suntharasami, P.; Desakron, V.; Migasena, S.; Bunnag, D.; Harinasuta, T. ELISA for immunodiagnosis of human gnathosomiasis. Southeast Asian J. Trop. Med. Public Health 1985, 16, 274–279. [Google Scholar]
- Dharmkrong-At, A.; Mmigasena, S.; Suntharasamai, P.; Bunnag, D.; Priwan, R.; Sirisinha, S. Enzyme-Linked Immunosorbent Assay for detection of antibody to Gnathostoma antigen in patients with intermittent cutaneous migratory swelling. J. Clin. Microbiol. 1986, 23, 847–851. [Google Scholar] [CrossRef]
- Maleewong, W.; Morakote, N.; Thamasonthi, W.; Charuchinda, K.; Tesana, S.; Khamboonruang, C. Serodiagnosis of human gnathostomiasis. Southeast Asian J. Trop. Med. Public Health 1988, 19, 201–205. [Google Scholar] [PubMed]
- Tapchaisri, P.; Nopparatana, C.; Chaicumpa, W.; Setasuban, P. Specific antigen of Gnathostoma spinigerum for immunodiagnosis of human gnathostomiasis. Int. J. Parasitol. 1991, 21, 315–319. [Google Scholar] [CrossRef]
- Laummaunwai, P.; Sawanyawisuth, K.; Intapan, P.M.; Chotmongkol, V.; Wongkham, C.; Maleewong, W. Evaluation of human IgG class and subclass antibodies to a 24 kDA antigenic component of Gnathostoma spinigerum for the serodiagnosis of gnathostomiasis. Parasitol. Res. 2007, 101, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Saenseeha, S.; Penchom, J.; Yamasaki, H.; Laummaunwai, P.; Tayapiwatana, C.; Kitkhuandee, A.; Maleewong, W.; Intapan, P.M. A dot-ELISA test using a Gnathostoma spinigerum recombinant matrix metalloproteinase protein for the serodiagnosis of human gnathostomiasis. Southeast Asian J. Trop. Med. Public 2014, 45, 990–996. [Google Scholar]
- Janwan, P.; Intapan, P.M.; Yamasaki, H.; Laummaunwai, P.; Sawanyawisuth, K.; Wongkham, C.; Tayapiwatana, C.; Kitkhuandee, A.; Lulitanond, V.; Nawa, Y.; et al. A recombinant matrix metalloproteinase protein from Gnathostoma spinigerum for serodiagnosis of neurognathostomiasis. Korean J. Parasitol. 2013, 51, 751–754. [Google Scholar] [CrossRef]
- Uparanukraw, P.; Morakote, N.; Harnnoi, T.; Dantrakool, A. Molecular cloning of a gene encoding matrix metalloproteinase-like protein from Gnathostoma spinigerum. Parasitol. Res. 2001, 87, 751–757. [Google Scholar]
- Janwan, P.; Intapan, P.M.; Yamasaki, H.; Laummaunwai, P.; Sawanyawisuth, K.; Wongkham, C.; Tayapiwatana, C.; Kitkhuandee, A.; Lulitanond, V.; Nawa, Y.; et al. Application of recombinatnt Gnathostoma spinigerum matrix mettaloproteinase-like protein for serodiagnosis of human gnathostomiasis by immunoblotting. Am. J. Trop. Hyg. 2013, 89, 63–67. [Google Scholar] [CrossRef]
- Kongkerd, N.; Uparanukraw, P.; Morakote, N.; Sajid, M.; McKerrow, J.H. Identification and characterization of a cathepsin L-like cysteine protease from Gnathostoma spinigerum. Mol. Biochem. Parasitol. 2008, 160, 129–137. [Google Scholar] [CrossRef]
- Stachyra, A.; Zawistowska-Deniziak, A.; Basałaj, K.; Grzelak, S.; Gondek, M.; Bień-Kalinowska, J. The Immunological Properties of Recombinant Multi-Cystatin-Like Domain Protein From Trichinella Britovi Produced in Yeast. Front. Immunol. 2019, 11, 2420. [Google Scholar] [CrossRef]
- Nuamtanong, S.; Reamtong, O.; Phuphisut, O.; Chotsiri, P.; Malaithong, P.; Dekumyoy, P.; Adisakwattana, P. Transcriptome and excretory-sectroy proteome of infective-stage larvae of the nematode Gnathostoma spinigerum reveal potential immunodiagnostic targets for development. Parasite 2019, 26, 34. [Google Scholar] [CrossRef]
- Aebersold, R.; Mann, M. Mass spectrometry-based proteomics. Nature 2003, 422, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.; Jefferies, J.R.; Brophy, P.M. Parasite proteomics. Parasitol. Today 2000, 16, 400–403. [Google Scholar] [CrossRef]
- Mutapi, F. Helminth parasite proteomics: From experimental models to human infections. Parasitology 2012, 139, 1195–1204. [Google Scholar] [CrossRef][Green Version]
- Chen, W.; Wang, X.; Li, X.; Lv, X.; Zhou, C.; Deng, C.; Lei, H.; Men, J.; Fan, Y.; Liang, C.; et al. Molecular characterization of cathepsin B from Clonorchis sinensis excretory/secretory products and assessment of its potential for serodiagnosis of clonorchiasis. Parasit Vectors 2014, 4, 149. [Google Scholar] [CrossRef]
- Sun, G.G.; Wang, Z.O.; Liu, C.Y.; Jiang, P.; Liu, R.D.; Wen, H.; Qi, X.; Wang, L.; Cui, J. Early serodiangosis of trichinellosis by ELISA using excretory-secretory antigens of Trichinella spiralis adult worms. Parasit Vectors 2015, 8, 1–8. [Google Scholar] [CrossRef]
- Ranganathan, S.; Garg, G. Secretome: Clues into pathogen infection and clinical applications. Genome Med. 2009, 113, 1–7. [Google Scholar] [CrossRef]
- Vibanco-Pérez, N.; Durán-Avelar, M.D.J.; Zambrano-Zaragoza, J.F.; Ventura-Ramón, G.H. Proteases secreted by Gnathostoma binucleatum degrade fibronectin and antibodies from mammals. Helminthologia 2015, 52, 6–10. [Google Scholar] [CrossRef]
- Sorsa, T.; Tervahartiala, T.; Leppilahti, J.; Hernandez, M.; Gamonal, J.; Tuomainen, M.A.; Lauhio, A.; Pussinen, P.J.; Mäntylä, P. Collagenase-2 (MMP-8) as a point-of-care biomarker in periodontitis and cardiovascular diseases. Therapeutic response to non-antimicrobial properties of tetracyclines. Pharmacol. Res. 2011, 63, 108–113. [Google Scholar] [CrossRef]
- Law, R.H.P.; Zhang, Q.; McGowan, S.; Buckle, A.M.; A Silverman, G.; Wong, W.; Rosado, C.J.; Langendorf, C.G.; Pike, R.N.; I Bird, P.; et al. An overview of the serpin superfamily. Genome Biol. 2006, 7, 216. [Google Scholar] [CrossRef][Green Version]
- Song, Y.Y.; Zhang, Y.; Ren, H.N.; Sun, G.G.; Qi, X.; Yang, F.; Jiang, P.; Zhang, X.; Cui, J.; Wang, Z.Q. Characterization of a serine protease inhibitor from Trichinella spiralis and its participation in larval invasion of host’s intestinal epithelial cells. Parasites Vectors 2018, 11, 499. [Google Scholar] [CrossRef]
- Wongkamchai, S.; Chiangjong, W.; Sinchaikul, S.; Chen, S.-T.; Choochote, W.; Thongboonkerd, V. Identification of Brugia malayi immunogens by an immunoproteomics approach. J. Proteom. 2011, 74, 1607–1613. [Google Scholar] [CrossRef] [PubMed]
- Campista-León, S.; Delgado-Vargas, F.; Ríos-Sicairos, J.; Landa, A.; Willms, K.; Bojórquez-Contreras, N.; Diaz-Camacho, S.P.; López-Moreno, H.S.; Mendoza-Hernández, G. Identification of Immunodominant Peptides from Gnathostoma binucleatum. Am. J. Trop. Med. Hyg. 2012, 87, 888–896. [Google Scholar] [CrossRef]
- Laummaunwai, P.; Intapan, P.M.; Wongkham, C.; Lulitanond, V.; Maleewong, W. Identification of antigenic components of Gnathostoma spinigerum advanced third stage larvae by two-dimensional gel electrophoresis and mass spectrometry. Southeast Asian J. Trop. Med. Public Health 2008, 39, 19–25. [Google Scholar]
- Janwan, P.; Intapan, P.M.; Laummaunwai, P.; Rodpai, R.; Wongkham, C.; Insawang, T.; Thanchomnang, T.; Sanpool, O.; Maleewong, W. Proteomic analysis identification of antigenic proteins in Gnathostoma spinigerum larvae. Exp. Parasitol. 2015, 159, 53–58. [Google Scholar] [CrossRef]
- Sugaroon, S.; Wiwanitkit, V. Gnathostoma infective stage larvae in swamp eels (Fluta alba) at a metropolitan market in Bangkok, Thailand. Ann. Clin. Lab. Sci. 2003, 33, 94–96. [Google Scholar]
- Sieu, T.; Dung, T.; Nga, N.; Hien, T.; Dalsgaard, A.; Waikagul, J.; Murrell, D. Prevalence of Gnathostoma spinigerum infection in wild and cultured swamp eels in Vietnam. J. Parasitol. 2009, 95, 246–248. [Google Scholar] [CrossRef]
- Veríssimo, C.M.; Graeff-Teixeira, C.; Jones, M.K.; Morassutti, A.L. Glycans in the roles of parasitological diagnosis and host-parasite interplay. Parasitology 2019, 146, 1217–1232. [Google Scholar] [CrossRef]
- Singh, A. Glycoproteomics. Nat. Methods 2021, 18, 28. [Google Scholar] [CrossRef]
- Tissot, B.; North, S.J.; Ceroni, A.; Pang, P.; Panico, M.; Rosati, M.; Capone, A.; Haslam, S.M.; Dell, A.; Morris, H.R. Glycoproteomics: Past, present and future. FEBS Lett. 2009, 583, 1728–1735. [Google Scholar] [CrossRef] [PubMed]
- Mitcheson, D.F.; Tobin, A.B.; Alam, M.M. Applying chemical genetic tools to the study of phospho-signalling pathways in malaria parasites. Biochim. Biophys. Acta 2015, 1854, 1650–1656. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Antigen | Separation- | Database for Protein Identification | Protein Identification | Ref. |
|---|---|---|---|---|
| G. binucleatum CWA | pH 5–8, 12% gel | Metazoa of NCBInr | Galectins | [52] |
| G. binucleatum ESP | pH 5–8, 12% gel | Metazoa of NCBInr | - | [52] |
| G. spinigerum CWA | pH 3–11, 12% gel | nr.fasta database | Cyclophilin Hypothetical protein Actin Matrix metalloproteinase-like Intermediate filament protein B | [53] |
| G. spinigerum CWA | pH 3–11, 12% gel | All entries of NCBI protein database | As37 Actin 2 Heat shock protein 90 Heat shock protein 70 Chaperonine protein HSP60 Chaperone protein DnaK Phosphoenolpyruvate Carboxykinase domain-containing Protein Carboxyl transferase domain protein Enolase Glyceraldehyde-3-phosphate dehydrogenase Peptidyl-prolylcis-trans isomerase Cyclophilin Cytoplasmic intermediate filament protein Peroxiredoxin Matrix metalloproteinase-like protein Fructose-bisphosphate aldolase 53 kDa Excretory/secretory protein Glu/Leu/Phe/Val dehydrogenase, dimerization domain protein Galectin methylmalonyl 4-Hydroxybutyrate coenzyme a transferase Phosphoglycerate mutase Eukaryotic translation elongation factor 1A Phosphoglycerate kinase Proteasome subunit alpha type 7-1 Myosin heavy chain Cytoplasmic Cu/Zn-superoxide dismutase CBN-MCE-1 protein ATPase and cell division protein 48 and Vps4 oligomer ization Kinesin-2 | [54] |
| G. spinigerum ESP | pH 3–10, 12% gel | In-house transcriptome | Serine_rich_NEDD9 Catalase heme-binding enzyme Pyrroline-5-carboxylate reductase Phospho-2-dehydro-3-deoxyheptonate aldolase Serpin AJAP1/PANP C-terminus | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thiangtrongjit, T.; Nogrado, K.; Ketboonlue, T.; Malaitong, P.; Adisakwattana, P.; Reamtong, O. Proteomics of Gnathostomiasis: A Way Forward for Diagnosis and Treatment Development. Pathogens 2021, 10, 1080. https://doi.org/10.3390/pathogens10091080
Thiangtrongjit T, Nogrado K, Ketboonlue T, Malaitong P, Adisakwattana P, Reamtong O. Proteomics of Gnathostomiasis: A Way Forward for Diagnosis and Treatment Development. Pathogens. 2021; 10(9):1080. https://doi.org/10.3390/pathogens10091080
Chicago/Turabian StyleThiangtrongjit, Tipparat, Kathyleen Nogrado, Thawatchai Ketboonlue, Preeyarat Malaitong, Poom Adisakwattana, and Onrapak Reamtong. 2021. "Proteomics of Gnathostomiasis: A Way Forward for Diagnosis and Treatment Development" Pathogens 10, no. 9: 1080. https://doi.org/10.3390/pathogens10091080
APA StyleThiangtrongjit, T., Nogrado, K., Ketboonlue, T., Malaitong, P., Adisakwattana, P., & Reamtong, O. (2021). Proteomics of Gnathostomiasis: A Way Forward for Diagnosis and Treatment Development. Pathogens, 10(9), 1080. https://doi.org/10.3390/pathogens10091080