
Antigen-Presenting Cells in the Airways: Moderating Asymptomatic Bacterial Carriage



{kind=link}
{kind=link}
Abstract
1. Introduction
2. Infection of the Respiratory Tract and Carriage of Respiratory Pathogens
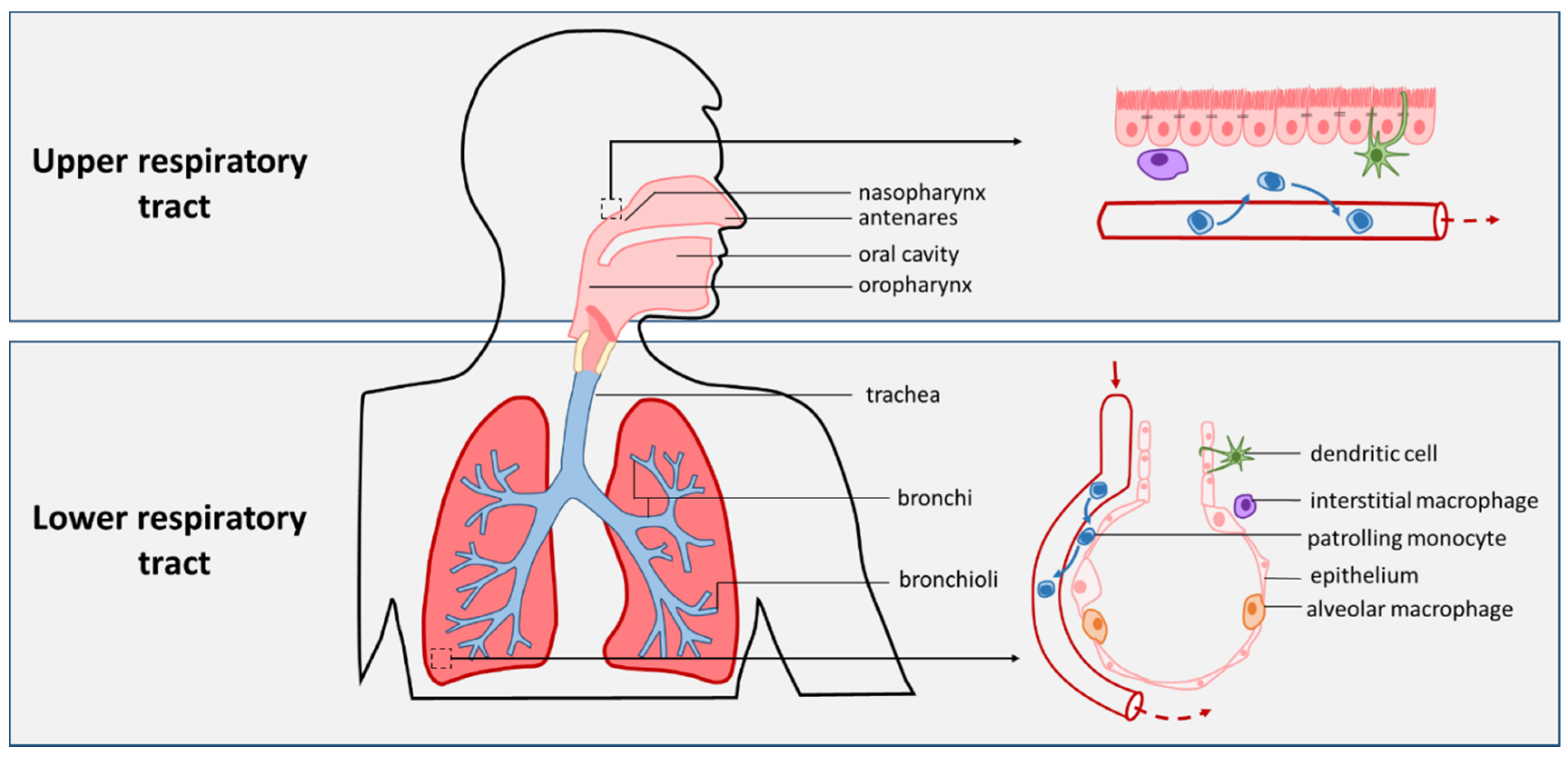
2.1. The Respiratory Tract and Pneumonia
2.2. Pneumonia-Causing Pathogens
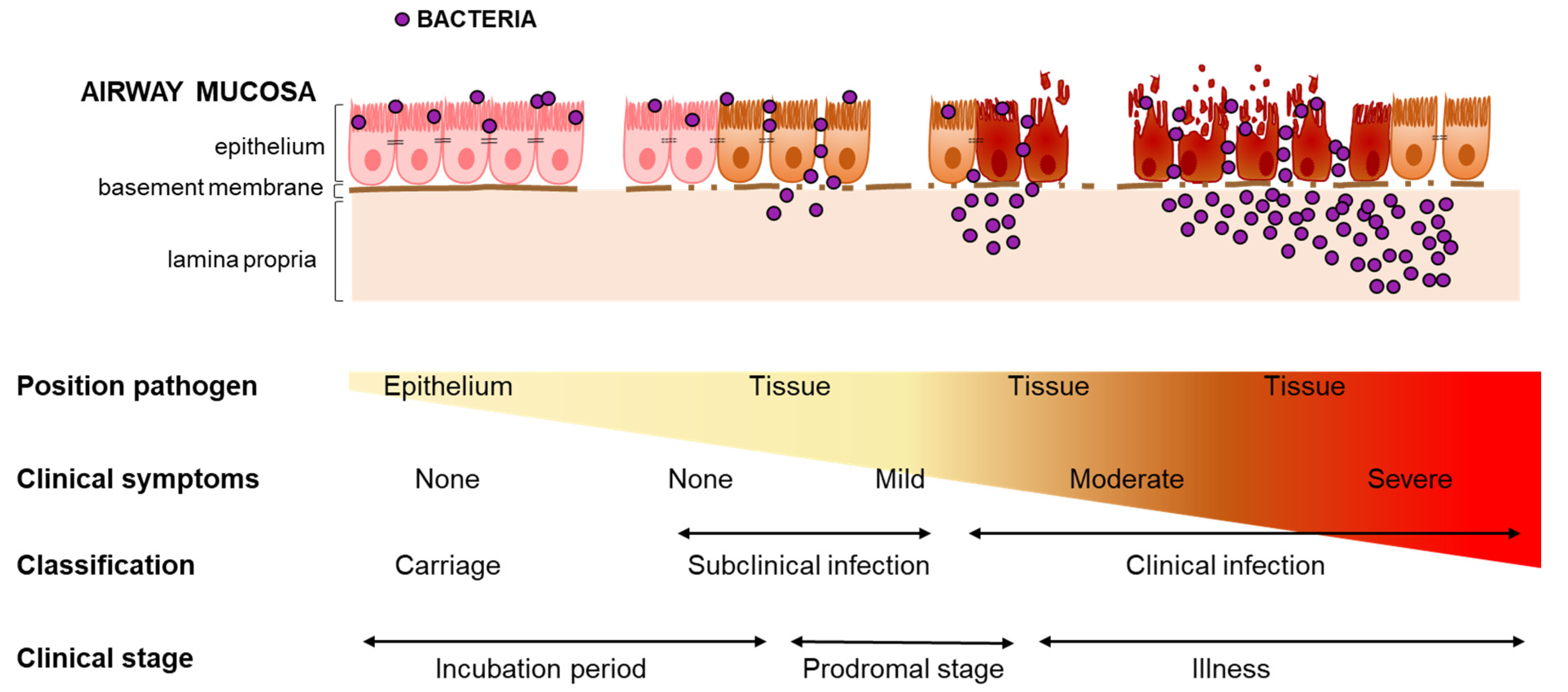
2.3. Defining Carriage and Infection
3. Progression from Carriage to Infection
4. Airway APCs in Infection and Carriage
4.1. APC-Pathogen Interactions during Infection
4.2. APC-Pathogen Interaction during Asymptomatic Carriage
4.3. APC Function and the Respiratory Microbiota
4.4. Commensal Bacteria Prime Airway APCs
5. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gehr, P. Annexe a. Anatomy and morphology of the respiratory tract. Ann. ICRP 1994, 24, 121–166. [Google Scholar] [CrossRef]
- Ball, M.; Hossain, M.; Padalia, D. Anatomy, Airway; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Patwa, A.; Shah, A. Anatomy and physiology of respiratory system relevant to anaesthesia. Indian J. Anaesth. 2015, 59, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, L.; Ochs, M. The micromechanics of lung alveoli: Structure and function of surfactant and tissue components. Histochem. Cell Biol. 2018, 150, 661–676. [Google Scholar] [CrossRef] [PubMed]
- Hakansson, A.P.; Orihuela, C.J.; Bogaert, D. Bacterial-host interactions: Physiology and pathophysiology of respiratory infection. Physiol. Rev. 2018, 98, 781–811. [Google Scholar] [CrossRef] [PubMed]
- Grief, S.N.; Loza, J.K. Guidelines for the evaluation and treatment of pneumonia. Prim. Care 2018, 45, 485–503. [Google Scholar] [CrossRef]
- Liu, L.; Oza, S.; Hogan, D.; Chu, Y.; Perin, J.; Zhu, J.; Lawn, J.E.; Cousens, S.; Mathers, C.; Black, R.E. Global, regional, and national causes of under-5 mortality in 2000-15: An updated systematic analysis with implications for the sustainable development goals. Lancet 2016, 388, 3027–3035. [Google Scholar] [CrossRef]
- United Nations Inter-Agency Group for Child Mortality Estimation. Levels & Trends in Child Mortality: Report 2020—Estimates Developed by the United Nations Inter-Agency Group for Child Mortality; United Nations Children’s Fund: New York, NY, USA, 2020.
- World Health Organisation. The Top 10 Causes of Death [Fact Sheet]. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 10 June 2021).
- Van de Maat, J.; van de Voort, E.; Mintegi, S.; Gervaix, A.; Nieboer, D.; Moll, H.; Oostenbrink, R. Research in European Pediatric Emergency Medicine Study Group. Antibiotic prescription for febrile children in european emergency departments: A cross-sectional, observational study. Lancet Infect. Dis. 2019, 19, 382–391. [Google Scholar] [CrossRef]
- Petersen, I.; Hayward, A.C.; Subgroup, S.S. Antibacterial prescribing in primary care. J. Antimicrob. Chemother. 2007, 60 (Suppl. 1), i43–i47. [Google Scholar] [CrossRef]
- Goodman, D.; Crocker, M.E.; Pervaiz, F.; McCollum, E.D.; Steenland, K.; Simkovich, S.M.; Miele, C.H.; Hammitt, L.L.; Herrera, P.; Zar, H.J.; et al. Challenges in the diagnosis of paediatric pneumonia in intervention field trials: Recommendations from a pneumonia field trial working group. Lancet Respir. Med. 2019, 7, 1068–1083. [Google Scholar] [CrossRef]
- Bénet, T.; Sánchez Picot, V.; Messaoudi, M.; Chou, M.; Eap, T.; Wang, J.; Shen, K.; Pape, J.W.; Rouzier, V.; Awasthi, S.; et al. Microorganisms associated with pneumonia in children <5 years of age in developing and emerging countries: The gabriel pneumonia multicenter, prospective, case-control study. Clin. Infect. Dis. 2017, 65, 604–612. [Google Scholar]
- Jain, S.; Self, W.H.; Wunderink, R.G.; Fakhran, S.; Balk, R.; Bramley, A.M.; Reed, C.; Grijalva, C.G.; Anderson, E.J.; Courtney, D.M.; et al. Community-acquired pneumonia requiring hospitalization among u.S. Adults. N. Engl. J. Med. 2015, 373, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Williams, D.J.; Arnold, S.R.; Ampofo, K.; Bramley, A.M.; Reed, C.; Stockmann, C.; Anderson, E.J.; Grijalva, C.G.; Self, W.H.; et al. Community-acquired pneumonia requiring hospitalization among u.S. Children. N. Engl. J. Med. 2015, 372, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Cillóniz, C.; Torres, A.; Niederman, M.; van der Eerden, M.; Chalmers, J.; Welte, T.; Blasi, F. Community-acquired pneumonia related to intracellular pathogens. Intensive Care Med. 2016, 42, 1374–1386. [Google Scholar] [CrossRef] [PubMed]
- Cilloniz, C.; Martin-Loeches, I.; Garcia-Vidal, C.; San Jose, A.; Torres, A. Microbial etiology of pneumonia: Epidemiology, diagnosis and resistance patterns. Int. J. Mol. Sci. 2016, 17, 2120. [Google Scholar] [CrossRef]
- Lanks, C.W.; Musani, A.I.; Hsia, D.W. Community-acquired pneumonia and hospital-acquired pneumonia. Med. Clin. N. Am. 2019, 103, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Cinetto, F.; Scarpa, R.; Rattazzi, M.; Agostini, C. The broad spectrum of lung diseases in primary antibody deficiencies. Eur. Respir. Rev. 2018, 27, 180019. [Google Scholar] [CrossRef]
- Letourneau, A.R.; Issa, N.C.; Baden, L.R. Pneumonia in the immunocompromised host. Curr. Opin. Pulm. Med. 2014, 20, 272–279. [Google Scholar] [CrossRef]
- Bhagirath, A.Y.; Li, Y.; Somayajula, D.; Dadashi, M.; Badr, S.; Duan, K. Cystic fibrosis lung environment and pseudomonas aeruginosa infection. BMC Pulm. Med. 2016, 16, 174. [Google Scholar] [CrossRef]
- Hassoun, A.; Huff, M.D.; Weisman, D.; Chahal, K.; Asis, E.; Stalons, D.; Grigorenko, E.; Green, J.; Malone, L.L.; Clemmons, S.; et al. Seasonal variation of respiratory pathogen colonization in asymptomatic health care professionals: A single-center, cross-sectional, 2-season observational study. Am. J. Infect. Control 2015, 43, 865–870. [Google Scholar] [CrossRef][Green Version]
- Rawlings, B.A.; Higgins, T.S.; Han, J.K. Bacterial pathogens in the nasopharynx, nasal cavity, and osteomeatal complex during wellness and viral infection. Am. J. Rhinol. Allergy 2013, 27, 39–42. [Google Scholar] [CrossRef]
- Krone, C.L.; Wyllie, A.L.; van Beek, J.; Rots, N.Y.; Oja, A.E.; Chu, M.L.; Bruin, J.P.; Bogaert, D.; Sanders, E.A.; Trzciński, K. Carriage of streptococcus pneumoniae in aged adults with influenza-like-illness. PLoS ONE 2015, 10, e0119875. [Google Scholar] [CrossRef]
- Wyllie, A.L.; Rümke, L.W.; Arp, K.; Bosch, A.; Bruin, J.P.; Rots, N.Y.; Wijmenga-Monsuur, A.J.; Sanders, E.A.M.; Trzciński, K. Molecular surveillance on streptococcus pneumoniae carriage in non-elderly adults; little evidence for pneumococcal circulation independent from the reservoir in children. Sci. Rep. 2016, 6, 34888. [Google Scholar] [CrossRef]
- Chi, D.H.; Hendley, J.O.; French, P.; Arango, P.; Hayden, F.G.; Winther, B. Nasopharyngeal reservoir of bacterial otitis media and sinusitis pathogens in adults during wellness and viral respiratory illness. Am. J. Rhinol. 2003, 17, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Gray, B.M.; Converse, G.M., 3rd; Dillon, H.C., Jr. Epidemiologic studies of streptococcus pneumoniae in infants: Acquisition, carriage, and infection during the first 24 months of life. J. Infect. Dis. 1980, 142, 923–933. [Google Scholar] [CrossRef]
- DeMuri, G.P.; Gern, J.E.; Eickhoff, J.C.; Lynch, S.V.; Wald, E.R. Dynamics of bacterial colonization with streptococcus pneumoniae, haemophilus influenzae, and moraxella catarrhalis during symptomatic and asymptomatic viral upper respiratory tract infection. Clin. Infect. Dis. 2018, 66, 1045–1053. [Google Scholar] [CrossRef]
- Spuesens, E.B.; Fraaij, P.L.; Visser, E.G.; Hoogenboezem, T.; Hop, W.C.; van Adrichem, L.N.; Weber, F.; Moll, H.A.; Broekman, B.; Berger, M.Y.; et al. Carriage of mycoplasma pneumoniae in the upper respiratory tract of symptomatic and asymptomatic children: An observational study. PLoS Med. 2013, 10, e1001444. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Coimbra, J.; Sarda, C.; Rello, J. Burden of community-acquired pneumonia and unmet clinical needs. Adv. Ther. 2020, 37, 1302–1318. [Google Scholar] [CrossRef]
- Hill, P.C.; Cheung, Y.B.; Akisanya, A.; Sankareh, K.; Lahai, G.; Greenwood, B.M.; Adegbola, R.A. Nasopharyngeal carriage of streptococcus pneumoniae in gambian infants: A longitudinal study. Clin. Infect. Dis. 2008, 46, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.C.; Björkman, P.; Persson, K. Polymerase chain reaction is superior to serology for the diagnosis of acute mycoplasma pneumoniae infection and reveals a high rate of persistent infection. BMC Microbiol. 2008, 8, 93. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.R.; Hill, V.L.; Burks, M.L.; Peters, J.I.; Singh, H.; Kannan, T.R.; Vale, S.; Cagle, M.P.; Principe, M.F.; Baseman, J.B.; et al. Mycoplasma pneumoniae in children with acute and refractory asthma. Ann. Allergy Asthma Immunol. 2013, 110, 328–334.e321. [Google Scholar] [CrossRef]
- Foy, H.M. Infections caused by mycoplasma pneumoniae and possible carrier state in different populations of patients. Clin. Infect. Dis. 1993, 17 (Suppl. 1), S37–S46. [Google Scholar] [CrossRef]
- Dorigo-Zetsma, J.W.; Wilbrink, B.; van der Nat, H.; Bartelds, A.I.; Heijnen, M.L.; Dankert, J. Results of molecular detection of mycoplasma pneumoniae among patients with acute respiratory infection and in their household contacts reveals children as human reservoirs. J. Infect. Dis. 2001, 183, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Siegel, S.J.; Weiser, J.N. Mechanisms of bacterial colonization of the respiratory tract. Annu. Rev. Microbiol. 2015, 69, 425–444. [Google Scholar] [CrossRef]
- Simell, B.; Auranen, K.; Käyhty, H.; Goldblatt, D.; Dagan, R.; O’Brien, K.L.; Pneumococcal Carriage, G. The fundamental link between pneumococcal carriage and disease. Expert Rev. Vaccines 2012, 11, 841–855. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nat. Immunol. 2004, 5, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Syrjänen, R.K.; Herva, E.E.; Mäkelä, P.H.; Puhakka, H.J.; Auranen, K.J.; Takala, A.K.; Kilpi, T.M. The value of nasopharyngeal culture in predicting the etiology of acute otitis media in children less than two years of age. Pediatr. Infect. Dis. J. 2006, 25, 1032–1036. [Google Scholar] [CrossRef]
- Tang, J.; Hui, J.; Ma, J.; Mingquan, C. Nasal decolonization of staphylococcus aureus and the risk of surgical site infection after surgery: A meta-analysis. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 33. [Google Scholar] [CrossRef]
- Williams, R.E. Healthy carriage of staphylococcus aureus: Its prevalence and importance. Bacteriol. Rev. 1963, 27, 56–71. [Google Scholar] [CrossRef]
- Goyal, N.; Miller, A.; Tripathi, M.; Parvizi, J. Methicillin-resistant staphylococcus aureus (mrsa): Colonisation and pre-operative screening. Bone Jt. J. 2013, 95-B, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Bode, L.G.M.; Kluytmans, J.A.J.W.; Wertheim, H.F.L.; Bogaers, D.; Vandenbroucke-Grauls, C.M.J.E.; Roosendaal, R.; Troelstra, A.; Box, A.T.A.; Voss, A.; van der Tweel, I.; et al. Preventing surgical-site infections in nasal carriers of staphylococcus aureus. N. Engl. J. Med. 2010, 362, 9–17. [Google Scholar] [CrossRef]
- Wertheim, H.F.; Melles, D.C.; Vos, M.C.; van Leeuwen, W.; van Belkum, A.; Verbrugh, H.A.; Nouwen, J.L. The role of nasal carriage in staphylococcus aureus infections. Lancet Infect. Dis. 2005, 5, 751–762. [Google Scholar] [CrossRef]
- Albrich, W.C.; Madhi, S.A.; Adrian, P.V.; van Niekerk, N.; Mareletsi, T.; Cutland, C.; Wong, M.; Khoosal, M.; Karstaedt, A.; Zhao, P.; et al. Use of a rapid test of pneumococcal colonization density to diagnose pneumococcal pneumonia. Clin. Infect. Dis. 2012, 54, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Thors, V.; Christensen, H.; Morales-Aza, B.; Oliver, E.; Sikora, P.; Vipond, I.; Muir, P.; Finn, A. High-density bacterial nasal carriage in children is transient and associated with respiratory viral infections—Implications for transmission dynamics. Pediatr. Infect. Dis. J. 2019, 38, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Thors, V.; Morales-Aza, B.; Pidwill, G.; Vipond, I.; Muir, P.; Finn, A. Population density profiles of nasopharyngeal carriage of 5 bacterial species in pre-school children measured using quantitative pcr offer potential insights into the dynamics of transmission. Hum. Vaccines Immunother. 2016, 12, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Physical stress and bacterial colonization. FEMS Microbiol. Rev. 2014, 38, 1250–1270. [Google Scholar] [CrossRef] [PubMed]
- Leiva-Juárez, M.M.; Kolls, J.K.; Evans, S.E. Lung epithelial cells: Therapeutically inducible effectors of antimicrobial defense. Mucosal Immunol. 2018, 11, 21–34. [Google Scholar] [CrossRef]
- Desch, A.N.; Gibbings, S.L.; Goyal, R.; Kolde, R.; Bednarek, J.; Bruno, T.; Slansky, J.E.; Jacobelli, J.; Mason, R.; Ito, Y.; et al. Flow cytometric analysis of mononuclear phagocytes in nondiseased human lung and lung-draining lymph nodes. Am. J. Respir. Crit. Care Med. 2016, 193, 614–626. [Google Scholar] [CrossRef]
- Baharom, F.; Rankin, G.; Blomberg, A.; Smed-Sörensen, A. Human lung mononuclear phagocytes in health and disease. Front. Immunol. 2017, 8, 499. [Google Scholar] [CrossRef]
- Pai, S.; Muruganandah, V.; Kupz, A. What lies beneath the airway mucosal barrier? Throwing the spotlight on antigen-presenting cell function in the lower respiratory tract. Clin. Transl. Immunol. 2020, 9, e1158. [Google Scholar] [CrossRef]
- Westphalen, K.; Gusarova, G.A.; Islam, M.N.; Subramanian, M.; Cohen, T.S.; Prince, A.S.; Bhattacharya, J. Sessile alveolar macrophages communicate with alveolar epithelium to modulate immunity. Nature 2014, 506, 503–506. [Google Scholar] [CrossRef]
- Guilliams, M.; Lambrecht, B.N.; Hammad, H. Division of labor between lung dendritic cells and macrophages in the defense against pulmonary infections. Mucosal Immunol. 2013, 6, 464–473. [Google Scholar] [CrossRef]
- Mitsi, E.; Kamng’ona, R.; Rylance, J.; Solórzano, C.; Jesus Reiné, J.; Mwandumba, H.C.; Ferreira, D.M.; Jambo, K.C. Human alveolar macrophages predominately express combined classical m1 and m2 surface markers in steady state. Respir. Res. 2018, 19, 66. [Google Scholar] [CrossRef]
- Tschernig, T.; Debertin, A.S.; Paulsen, F.; Kleemann, W.J.; Pabst, R. Dendritic cells in the mucosa of the human trachea are not regularly found in the first year of life. Thorax 2001, 56, 427–431. [Google Scholar] [CrossRef]
- Todate, A.; Chida, K.; Suda, T.; Imokawa, S.; Sato, J.; Ide, K.; Tsuchiya, T.; Inui, N.; Nakamura, Y.; Asada, K.; et al. Increased numbers of dendritic cells in the bronchiolar tissues of diffuse panbronchiolitis. Am. J. Respir. Crit. Care Med. 2000, 162, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Baharom, F.; Thomas, S.; Rankin, G.; Lepzien, R.; Pourazar, J.; Behndig, A.F.; Ahlm, C.; Blomberg, A.; Smed-Sörensen, A. Dendritic cells and monocytes with distinct inflammatory responses reside in lung mucosa of healthy humans. J. Immunol. 2016, 196, 4498–4509. [Google Scholar] [CrossRef] [PubMed]
- Förster, R.; Schubel, A.; Breitfeld, D.; Kremmer, E.; Renner-Müller, I.; Wolf, E.; Lipp, M. Ccr7 coordinates the primary immune response by establishing functional microenvironments in secondary lymphoid organs. Cell 1999, 99, 23–33. [Google Scholar] [CrossRef]
- Del Rio, M.L.; Rodriguez-Barbosa, J.I.; Kremmer, E.; Förster, R. Cd103- and cd103+ bronchial lymph node dendritic cells are specialized in presenting and cross-presenting innocuous antigen to cd4+ and cd8+ t cells. J. Immunol. 2007, 178, 6861–6866. [Google Scholar] [CrossRef] [PubMed]
- Jakubzick, C.; Tacke, F.; Llodra, J.; van Rooijen, N.; Randolph, G.J. Modulation of dendritic cell trafficking to and from the airways. J. Immunol. 2006, 176, 3578–3584. [Google Scholar] [CrossRef]
- Patel, V.I.; Metcalf, J.P. Airway macrophage and dendritic cell subsets in the resting human lung. Crit. Rev. Immunol. 2018, 38, 303–331. [Google Scholar] [CrossRef] [PubMed]
- Haniffa, M.; Shin, A.; Bigley, V.; McGovern, N.; Teo, P.; See, P.; Wasan, P.S.; Wang, X.N.; Malinarich, F.; Malleret, B.; et al. Human tissues contain cd141hi cross-presenting dendritic cells with functional homology to mouse cd103+ nonlymphoid dendritic cells. Immunity 2012, 37, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Schlitzer, A.; McGovern, N.; Teo, P.; Zelante, T.; Atarashi, K.; Low, D.; Ho, A.W.; See, P.; Shin, A.; Wasan, P.S.; et al. Irf4 transcription factor-dependent cd11b+ dendritic cells in human and mouse control mucosal il-17 cytokine responses. Immunity 2013, 38, 970–983. [Google Scholar] [CrossRef] [PubMed]
- Colonna, M.; Trinchieri, G.; Liu, Y.J. Plasmacytoid dendritic cells in immunity. Nat. Immunol. 2004, 5, 1219–1226. [Google Scholar] [CrossRef]
- Rodrigues, P.F.; Alberti-Servera, L.; Eremin, A.; Grajales-Reyes, G.E.; Ivanek, R.; Tussiwand, R. Distinct progenitor lineages contribute to the heterogeneity of plasmacytoid dendritic cells. Nat. Immunol. 2018, 19, 711–722. [Google Scholar] [CrossRef]
- Jakubzick, C.; Gautier, E.L.; Gibbings, S.L.; Sojka, D.K.; Schlitzer, A.; Johnson, T.E.; Ivanov, S.; Duan, Q.; Bala, S.; Condon, T.; et al. Minimal differentiation of classical monocytes as they survey steady-state tissues and transport antigen to lymph nodes. Immunity 2013, 39, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Pamer, E.G. Monocyte recruitment during infection and inflammation. Nat. Rev. Immunol. 2011, 11, 762–774. [Google Scholar] [CrossRef]
- Jardine, L.; Wiscombe, S.; Reynolds, G.; McDonald, D.; Fuller, A.; Green, K.; Filby, A.; Forrest, I.; Ruchaud-Sparagano, M.-H.; Scott, J.; et al. Lipopolysaccharide inhalation recruits monocytes and dendritic cell subsets to the alveolar airspace. Nat. Commun. 2019, 10, 1999. [Google Scholar] [CrossRef]
- Rodero, M.P.; Poupel, L.; Loyher, P.L.; Hamon, P.; Licata, F.; Pessel, C.; Hume, D.A.; Combadière, C.; Boissonnas, A. Immune surveillance of the lung by migrating tissue monocytes. Elife 2015, 4, e07847. [Google Scholar] [CrossRef]
- Wright, A.K.; Bangert, M.; Gritzfeld, J.F.; Ferreira, D.M.; Jambo, K.C.; Wright, A.D.; Collins, A.M.; Gordon, S.B. Experimental human pneumococcal carriage augments il-17a-dependent t-cell defence of the lung. PLoS Pathog. 2013, 9, e1003274. [Google Scholar] [CrossRef]
- Mitsi, E.; Carniel, B.; Reiné, J.; Rylance, J.; Zaidi, S.; Soares-Schanoski, A.; Connor, V.; Collins, A.M.; Schlitzer, A.; Nikolaou, E.; et al. Nasal pneumococcal density is associated with microaspiration and heightened human alveolar macrophage responsiveness to bacterial pathogens. Am. J. Respir. Crit. Care Med. 2020, 201, 335–347. [Google Scholar] [CrossRef]
- Cole, A.L.; Muthukrishnan, G.; Chong, C.; Beavis, A.; Eade, C.R.; Wood, M.P.; Deichen, M.G.; Cole, A.M. Host innate inflammatory factors and staphylococcal protein a influence the duration of human staphylococcus aureus nasal carriage. Mucosal Immunol. 2016, 9, 1537–1548. [Google Scholar] [CrossRef]
- Cole, A.L.; Cosgrove Sweeney, Y.; Lasseter, A.G.; Gray, J.M.; Beavis, A.C.; Chong, C.F.; Hajheidari, S.V.; Beyene, A.; Patton, D.L.; Cole, A.M. Evaluation of the pig-tailed macaque (macaca nemestrina) as a model of human staphylococcus aureus nasal carriage. Infect. Immun. 2018, 86, e00043-18. [Google Scholar] [CrossRef] [PubMed]
- Dockrell, D.H.; Marriott, H.M.; Prince, L.R.; Ridger, V.C.; Ince, P.G.; Hellewell, P.G.; Whyte, M.K. Alveolar macrophage apoptosis contributes to pneumococcal clearance in a resolving model of pulmonary infection. J. Immunol. 2003, 171, 5380–5388. [Google Scholar] [CrossRef]
- Preston, J.A.; Bewley, M.A.; Marriott, H.M.; McGarry Houghton, A.; Mohasin, M.; Jubrail, J.; Morris, L.; Stephenson, Y.L.; Cross, S.; Greaves, D.R.; et al. Alveolar macrophage apoptosis-associated bacterial killing helps prevent murine pneumonia. Am. J. Respir. Crit. Care Med. 2019, 200, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Nikolaou, E.; Jochems, S.P.; Mitsi, E.; Pojar, S.; Blizard, A.; Reiné, J.; Solórzano, C.; Negera, E.; Carniel, B.; Soares-Schanoski, A.; et al. Experimental human challenge defines distinct pneumococcal kinetic profiles and mucosal responses between colonized and non-colonized adults. mBio 2021, 12, e02020-20. [Google Scholar] [CrossRef]
- Jochems, S.P.; de Ruiter, K.; Solorzano, C.; Voskamp, A.; Mitsi, E.; Nikolaou, E.; Carniel, B.F.; Pojar, S.; German, E.L.; Reine, J.; et al. Innate and adaptive nasal mucosal immune responses following experimental human pneumococcal colonization. J. Clin. Investig. 2019, 129, 4523–4538. [Google Scholar] [CrossRef]
- Linehan, E.; Fitzgerald, D.C. Ageing and the immune system: Focus on macrophages. Eur. J. Microbiol. Immunol. 2015, 5, 14–24. [Google Scholar] [CrossRef]
- Chapman, S.J.; Hill, A.V. Human genetic susceptibility to infectious disease. Nat. Rev. Genet. 2012, 13, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Martin, R.J.; Lafasto, S.; Efaw, B.J.; Rino, J.G.; Harbeck, R.J.; Chu, H.W. Toll-like receptor 2 down-regulation in established mouse allergic lungs contributes to decreased mycoplasma clearance. Am. J. Respir. Crit. Care Med. 2008, 177, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Lugg, S.T.; Scott, A.; Parekh, D.; Naidu, B.; Thickett, D.R. Cigarette smoke exposure and alveolar macrophages: Mechanisms for lung disease. Thorax 2021. [Google Scholar] [CrossRef]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.-C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome definition re-visited: Old concepts and new challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef]
- Reyman, M.; van Houten, M.A.; van Baarle, D.; Bosch, A.; Man, W.H.; Chu, M.; Arp, K.; Watson, R.L.; Sanders, E.A.M.; Fuentes, S.; et al. Impact of delivery mode-associated gut microbiota dynamics on health in the first year of life. Nat. Commun. 2019, 10, 4997. [Google Scholar] [CrossRef]
- Charlson, E.S.; Bittinger, K.; Haas, A.R.; Fitzgerald, A.S.; Frank, I.; Yadav, A.; Bushman, F.D.; Collman, R.G. Topographical continuity of bacterial populations in the healthy human respiratory tract. Am. J. Respir. Crit. Care Med. 2011, 184, 957–963. [Google Scholar] [CrossRef]
- Dickson, R.P.; Erb-Downward, J.R.; Freeman, C.M.; McCloskey, L.; Falkowski, N.R.; Huffnagle, G.B.; Curtis, J.L. Bacterial topography of the healthy human lower respiratory tract. mBio 2017, 8, e02287-16. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.M.; Young, V.B.; Huffnagle, G.B. The microbiome of the lung. Transl. Res. 2012, 160, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Yun, Y.; Srinivas, G.; Kuenzel, S.; Linnenbrink, M.; Alnahas, S.; Bruce, K.D.; Steinhoff, U.; Baines, J.F.; Schaible, U.E. Environmentally determined differences in the murine lung microbiota and their relation to alveolar architecture. PLoS ONE 2014, 9, e113466. [Google Scholar] [CrossRef] [PubMed]
- Huxley, E.J.; Viroslav, J.; Gray, W.R.; Pierce, A.K. Pharyngeal aspiration in normal adults and patients with depressed consciousness. Am. J. Med. 1978, 64, 564–568. [Google Scholar] [CrossRef]
- Gleeson, K.; Eggli, D.F.; Maxwell, S.L. Quantitative aspiration during sleep in normal subjects. Chest 1997, 111, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, R.; Watabe, N.; Konno, T.; Mishina, N.; Sekizawa, K.; Sasaki, H. High incidence of silent aspiration in elderly patients with community-acquired pneumonia. Am. J. Respir. Crit. Care Med. 1994, 150, 251–253. [Google Scholar] [CrossRef]
- Cvejic, L.; Harding, R.; Churchward, T.; Turton, A.; Finlay, P.; Massey, D.; Bardin, P.G.; Guy, P. Laryngeal penetration and aspiration in individuals with stable copd. Respirology 2011, 16, 269–275. [Google Scholar] [CrossRef]
- Harding, S.M. Gastroesophageal reflux, asthma, and mechanisms of interaction. Am. J. Med. 2001, 111 (Suppl. 8A), 8S–12S. [Google Scholar] [CrossRef]
- Brown, R.L.; Sequeira, R.P.; Clarke, T.B. The microbiota protects against respiratory infection via gm-csf signaling. Nat. Commun. 2017, 8, 1512. [Google Scholar] [CrossRef]
- Bernasconi, E.; Pattaroni, C.; Koutsokera, A.; Pison, C.; Kessler, R.; Benden, C.; Soccal, P.M.; Magnan, A.; Aubert, J.D.; Marsland, B.J.; et al. Airway microbiota determines innate cell inflammatory or tissue remodeling profiles in lung transplantation. Am. J. Respir. Crit. Care Med. 2016, 194, 1252–1263. [Google Scholar] [CrossRef]
- Abreu, N.A.; Nagalingam, N.A.; Song, Y.; Roediger, F.C.; Pletcher, S.D.; Goldberg, A.N.; Lynch, S.V. Sinus microbiome diversity depletion and corynebacterium tuberculostearicum enrichment mediates rhinosinusitis. Sci. Transl. Med. 2012, 4, 151ra124. [Google Scholar] [CrossRef]
- De Boeck, I.; Wittouck, S.; Martens, K.; Claes, J.; Jorissen, M.; Steelant, B.; van den Broek, M.F.L.; Seys, S.F.; Hellings, P.W.; Vanderveken, O.M.; et al. Anterior nares diversity and pathobionts represent sinus microbiome in chronic rhinosinusitis. mSphere 2019, 4, e00532-19. [Google Scholar] [CrossRef] [PubMed]
- De Steenhuijsen Piters, W.A.A.; Binkowska, J.; Bogaert, D. Early life microbiota and respiratory tract infections. Cell Host Microbe 2020, 28, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Man, W.H.; Clerc, M.; de Steenhuijsen Piters, W.A.A.; van Houten, M.A.; Chu, M.; Kool, J.; Keijser, B.J.F.; Sanders, E.A.M.; Bogaert, D. Loss of microbial topography between oral and nasopharyngeal microbiota and development of respiratory infections early in life. Am. J. Respir. Crit. Care Med. 2019, 200, 760–770. [Google Scholar] [CrossRef]
- Sakwinska, O.; Bastic Schmid, V.; Berger, B.; Bruttin, A.; Keitel, K.; Lepage, M.; Moine, D.; Ngom Bru, C.; Brüssow, H.; Gervaix, A. Nasopharyngeal microbiota in healthy children and pneumonia patients. J. Clin. Microbiol. 2014, 52, 1590–1594. [Google Scholar] [CrossRef]
- Netusha, T.; Fiona, J.W.; Alicja, P.; Eta, A.; Laura, R.; Michael, G.S.; Dawn, M.E.B.; Camilli, A. Streptococcus pneumoniae colonization disrupts the microbial community within the upper respiratory tract of aging mice. Infect. Immun. 2016, 84, 906–916. [Google Scholar]
- Krone, C.L.; Biesbroek, G.; Trzciński, K.; Sanders, E.A.; Bogaert, D. Respiratory microbiota dynamics following streptococcus pneumoniae acquisition in young and elderly mice. Infect. Immun. 2014, 82, 1725–1731. [Google Scholar] [CrossRef]
- Li, N.; Ma, W.T.; Pang, M.; Fan, Q.L.; Hua, J.L. The commensal microbiota and viral infection: A comprehensive review. Front. Immunol. 2019, 10, 1551. [Google Scholar] [CrossRef] [PubMed]
- Segal, L.N.; Clemente, J.C.; Tsay, J.C.; Koralov, S.B.; Keller, B.C.; Wu, B.G.; Li, Y.; Shen, N.; Ghedin, E.; Morris, A.; et al. Enrichment of the lung microbiome with oral taxa is associated with lung inflammation of a th17 phenotype. Nat. Microbiol. 2016, 1, 16031. [Google Scholar] [CrossRef] [PubMed]
- Curtis, M.M.; Way, S.S. Interleukin-17 in host defence against bacterial, mycobacterial and fungal pathogens. Immunology 2009, 126, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.G.; Sulaiman, I.; Tsay, J.J.; Perez, L.; Franca, B.; Li, Y.; Wang, J.; Gonzalez, A.N.; El-Ashmawy, M.; Carpenito, J.; et al. Episodic aspiration with oral commensals induces a myd88-dependent, pulmonary t-helper cell type 17 response that mitigates susceptibility to streptococcus pneumoniae. Am. J. Respir. Crit. Care Med. 2021, 203, 1099–1111. [Google Scholar] [CrossRef]
- Larsen, J.M.; Steen-Jensen, D.B.; Laursen, J.M.; Søndergaard, J.N.; Musavian, H.S.; Butt, T.M.; Brix, S. Divergent pro-inflammatory profile of human dendritic cells in response to commensal and pathogenic bacteria associated with the airway microbiota. PLoS ONE 2012, 7, e31976. [Google Scholar] [CrossRef] [PubMed]
- Belladonna, M.L.; Renauld, J.C.; Bianchi, R.; Vacca, C.; Fallarino, F.; Orabona, C.; Fioretti, M.C.; Grohmann, U.; Puccetti, P. Il-23 and il-12 have overlapping, but distinct, effects on murine dendritic cells. J. Immunol. 2002, 168, 5448–5454. [Google Scholar] [CrossRef]
- Wang, J.; Li, F.; Sun, R.; Gao, X.; Wei, H.; Li, L.J.; Tian, Z. Bacterial colonization dampens influenza-mediated acute lung injury via induction of m2 alveolar macrophages. Nat. Commun. 2013, 4, 2106. [Google Scholar] [CrossRef]
- Roquilly, A.; Jacqueline, C.; Davieau, M.; Mollé, A.; Sadek, A.; Fourgeux, C.; Rooze, P.; Broquet, A.; Misme-Aucouturier, B.; Chaumette, T.; et al. Alveolar macrophages are epigenetically altered after inflammation, leading to long-term lung immunoparalysis. Nat. Immunol. 2020, 21, 636–648. [Google Scholar] [CrossRef]
- Park, B.S.; Song, D.H.; Kim, H.M.; Choi, B.S.; Lee, H.; Lee, J.O. The structural basis of lipopolysaccharide recognition by the tlr4-md-2 complex. Nature 2009, 458, 1191–1195. [Google Scholar] [CrossRef]
- Brix, S.; Eriksen, C.; Larsen, J.M.; Bisgaard, H. Metagenomic heterogeneity explains dual immune effects of endotoxins. J. Allergy Clin. Immunol. 2015, 135, 277–280. [Google Scholar] [CrossRef]
- Zhao, J.; Schloss, P.D.; Kalikin, L.M.; Carmody, L.A.; Foster, B.K.; Petrosino, J.F.; Cavalcoli, J.D.; VanDevanter, D.R.; Murray, S.; Li, J.Z.; et al. Decade-long bacterial community dynamics in cystic fibrosis airways. Proc. Natl. Acad. Sci. USA 2012, 109, 5809–5814. [Google Scholar] [CrossRef] [PubMed]
- Toraldo, D.M.; Conte, L. Influence of the lung microbiota dysbiosis in chronic obstructive pulmonary disease exacerbations: The controversial use of corticosteroid and antibiotic treatments and the role of eosinophils as a disease marker. J. Clin. Med. Res. 2019, 11, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.; Guarner, F.; Bustos Fernandez, L.; Maruy, A.; Sdepanian, V.L.; Cohen, H. Antibiotics as major disruptors of gut microbiota. Front. Cell. Infect. Microbiol. 2020, 10, 731. [Google Scholar] [CrossRef]
- Langdon, A.; Crook, N.; Dantas, G. The effects of antibiotics on the microbiome throughout development and alternative approaches for therapeutic modulation. Genome Med. 2016, 8, 39. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slimmen, L.J.M.; Janssens, H.M.; van Rossum, A.M.C.; Unger, W.W.J. Antigen-Presenting Cells in the Airways: Moderating Asymptomatic Bacterial Carriage. Pathogens 2021, 10, 945. https://doi.org/10.3390/pathogens10080945
Slimmen LJM, Janssens HM, van Rossum AMC, Unger WWJ. Antigen-Presenting Cells in the Airways: Moderating Asymptomatic Bacterial Carriage. Pathogens. 2021; 10(8):945. https://doi.org/10.3390/pathogens10080945
Chicago/Turabian StyleSlimmen, Lisa J. M., Hettie M. Janssens, Annemarie M. C. van Rossum, and Wendy W. J. Unger. 2021. "Antigen-Presenting Cells in the Airways: Moderating Asymptomatic Bacterial Carriage" Pathogens 10, no. 8: 945. https://doi.org/10.3390/pathogens10080945
APA StyleSlimmen, L. J. M., Janssens, H. M., van Rossum, A. M. C., & Unger, W. W. J. (2021). Antigen-Presenting Cells in the Airways: Moderating Asymptomatic Bacterial Carriage. Pathogens, 10(8), 945. https://doi.org/10.3390/pathogens10080945