
Gut Microbiota in Military International Travelers with Doxycycline Malaria Prophylaxis: Towards the Risk of a Simpson Paradox in the Human Microbiome Field

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Clinical Characteristics of the Study Population
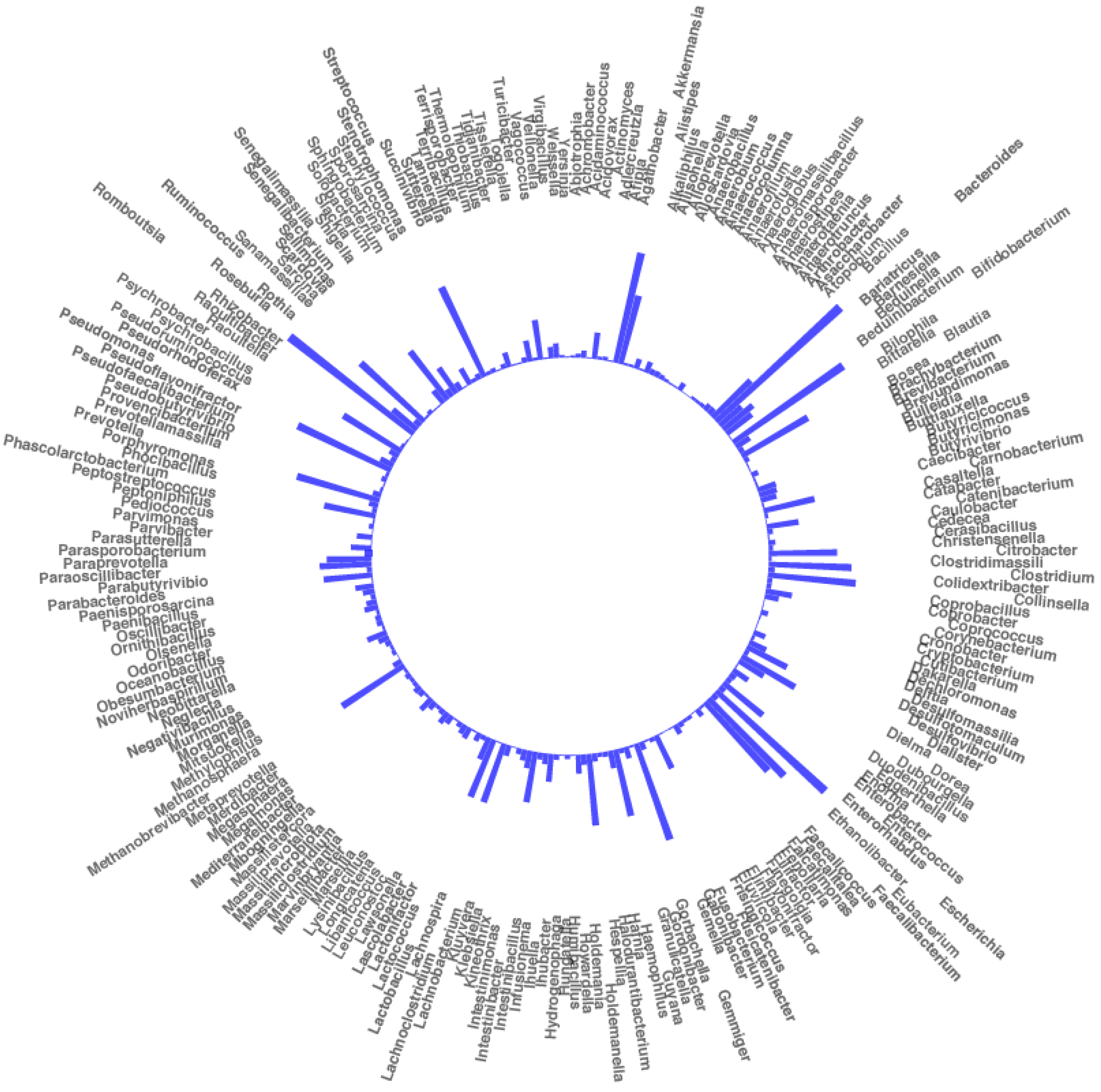
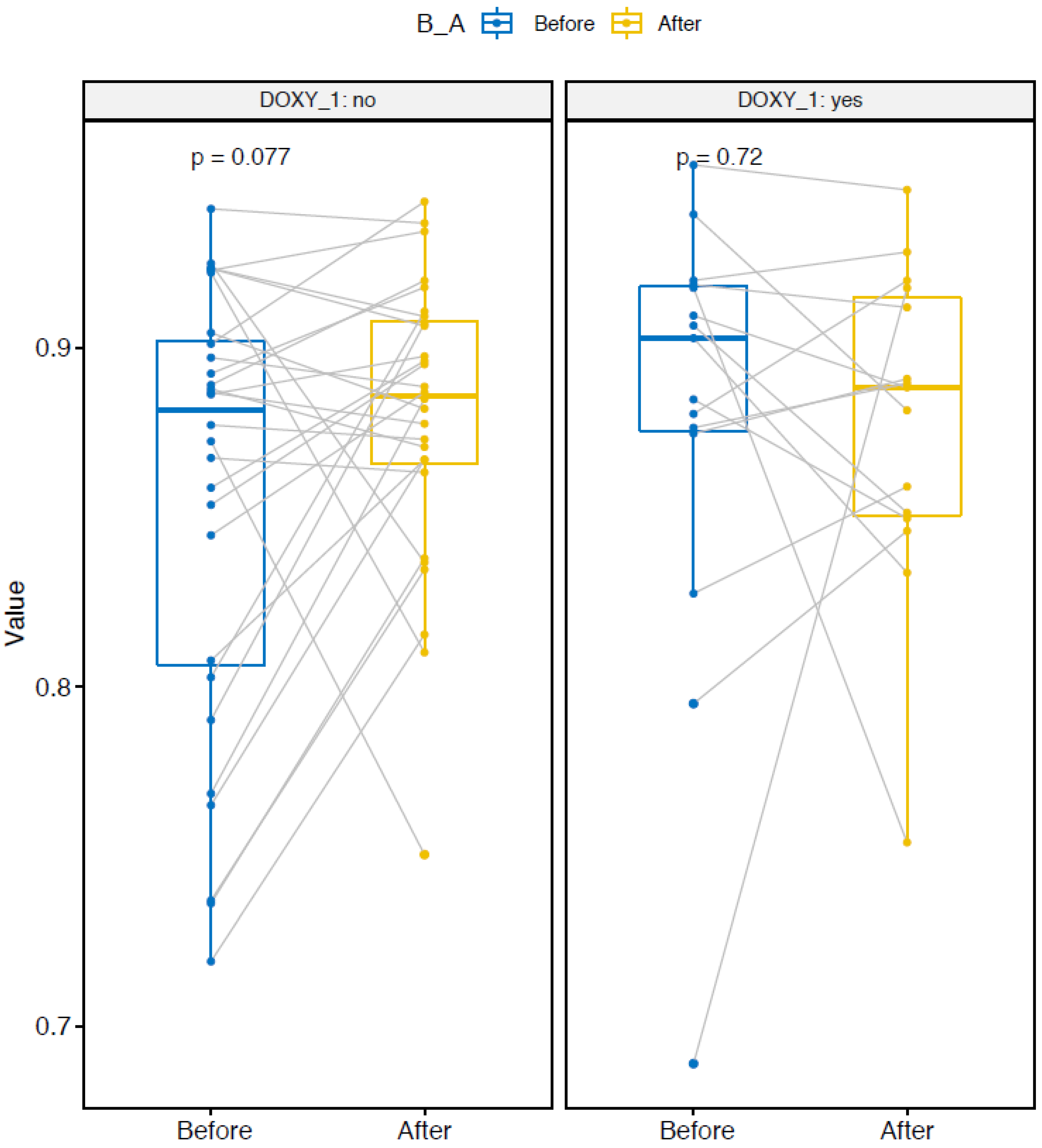
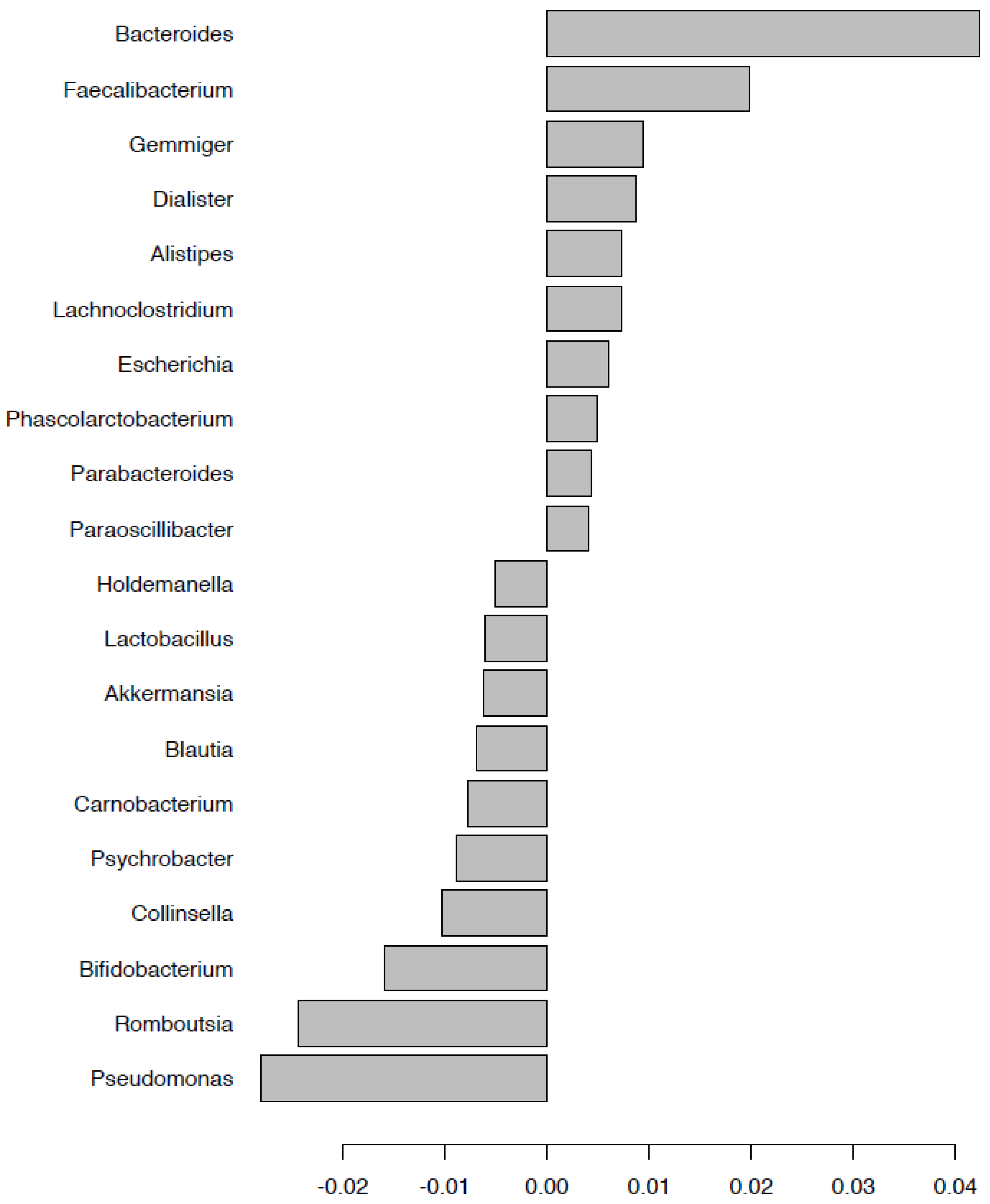
2.2. Changes in Bacterial Community Structure
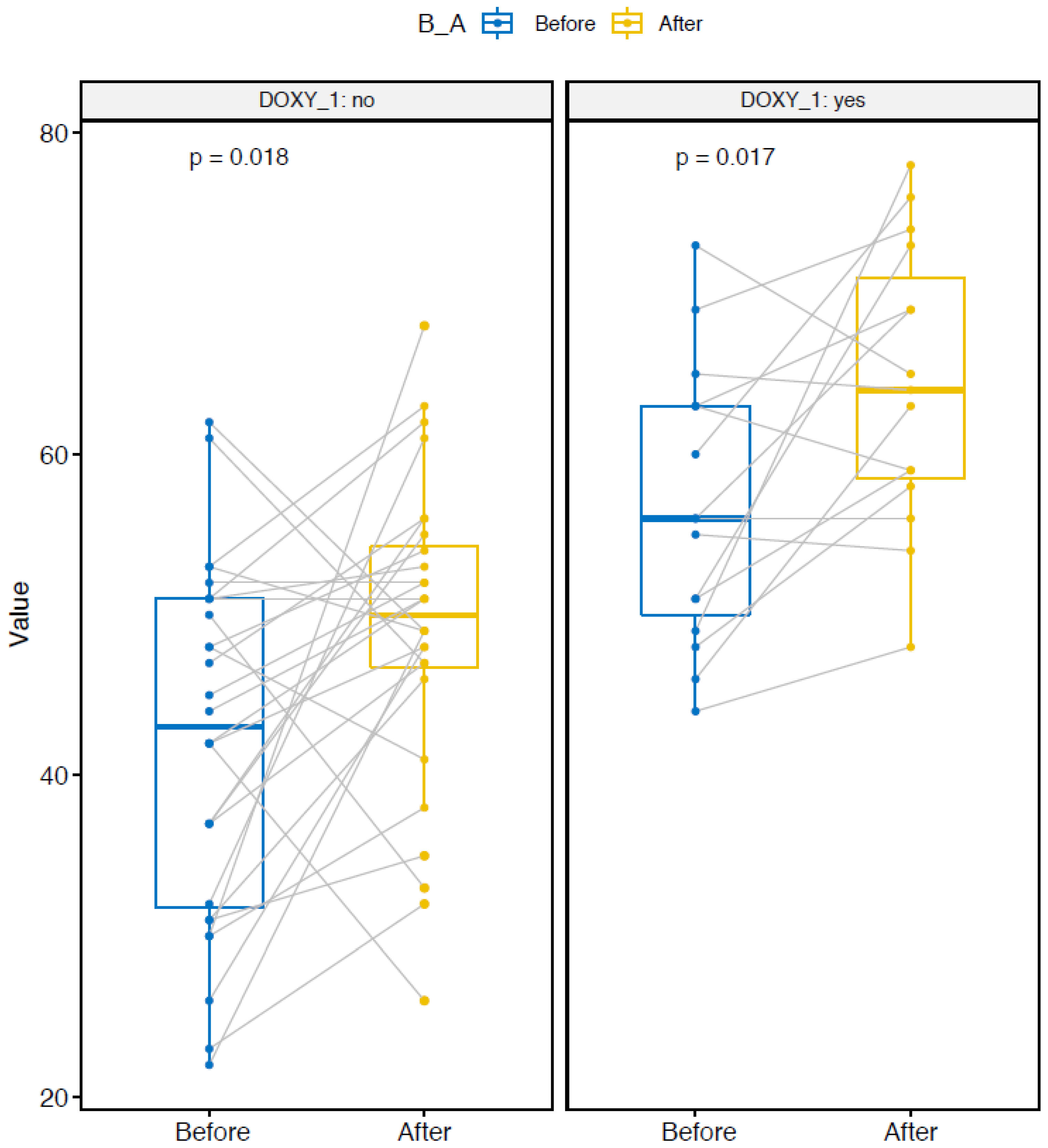
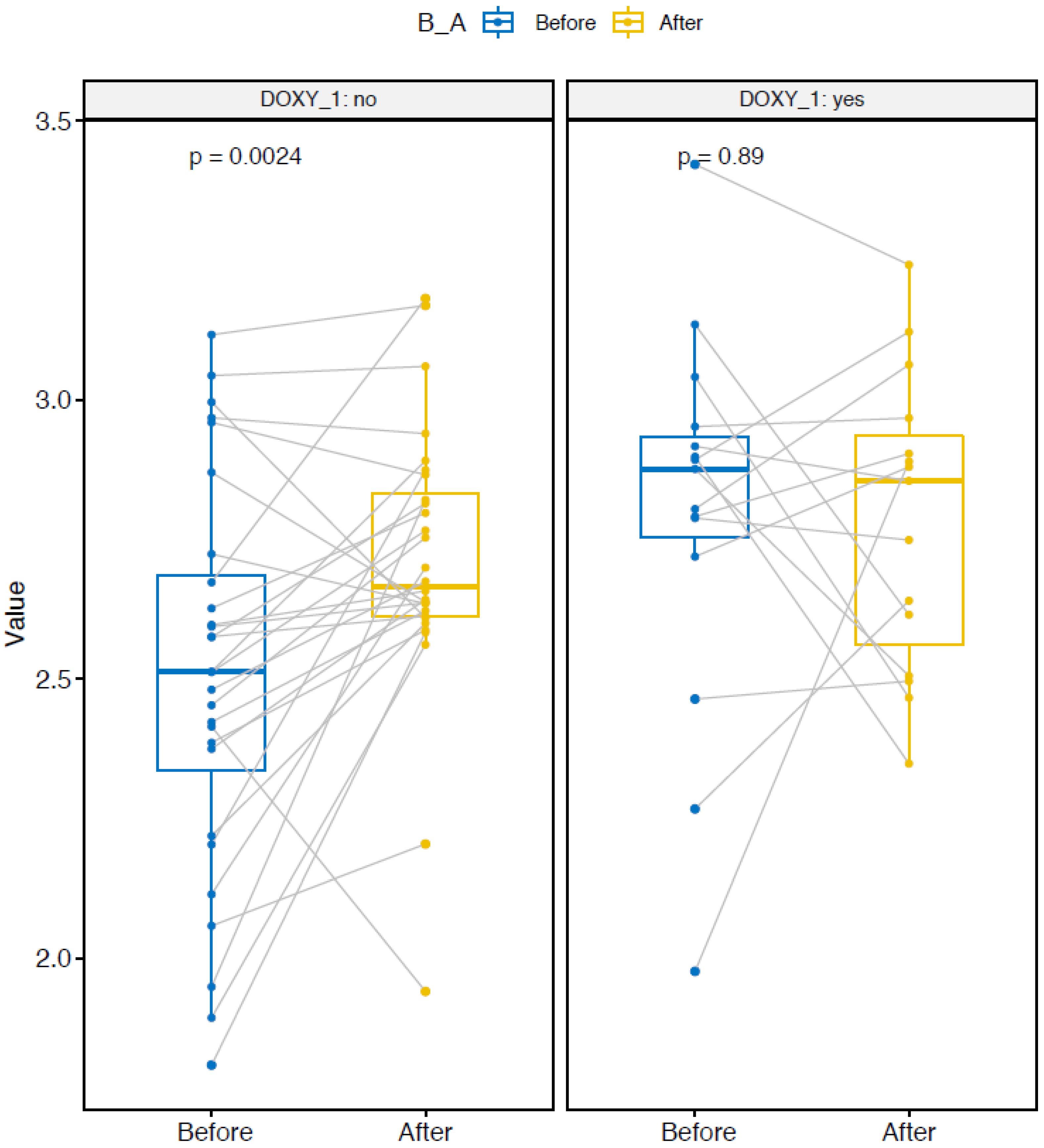
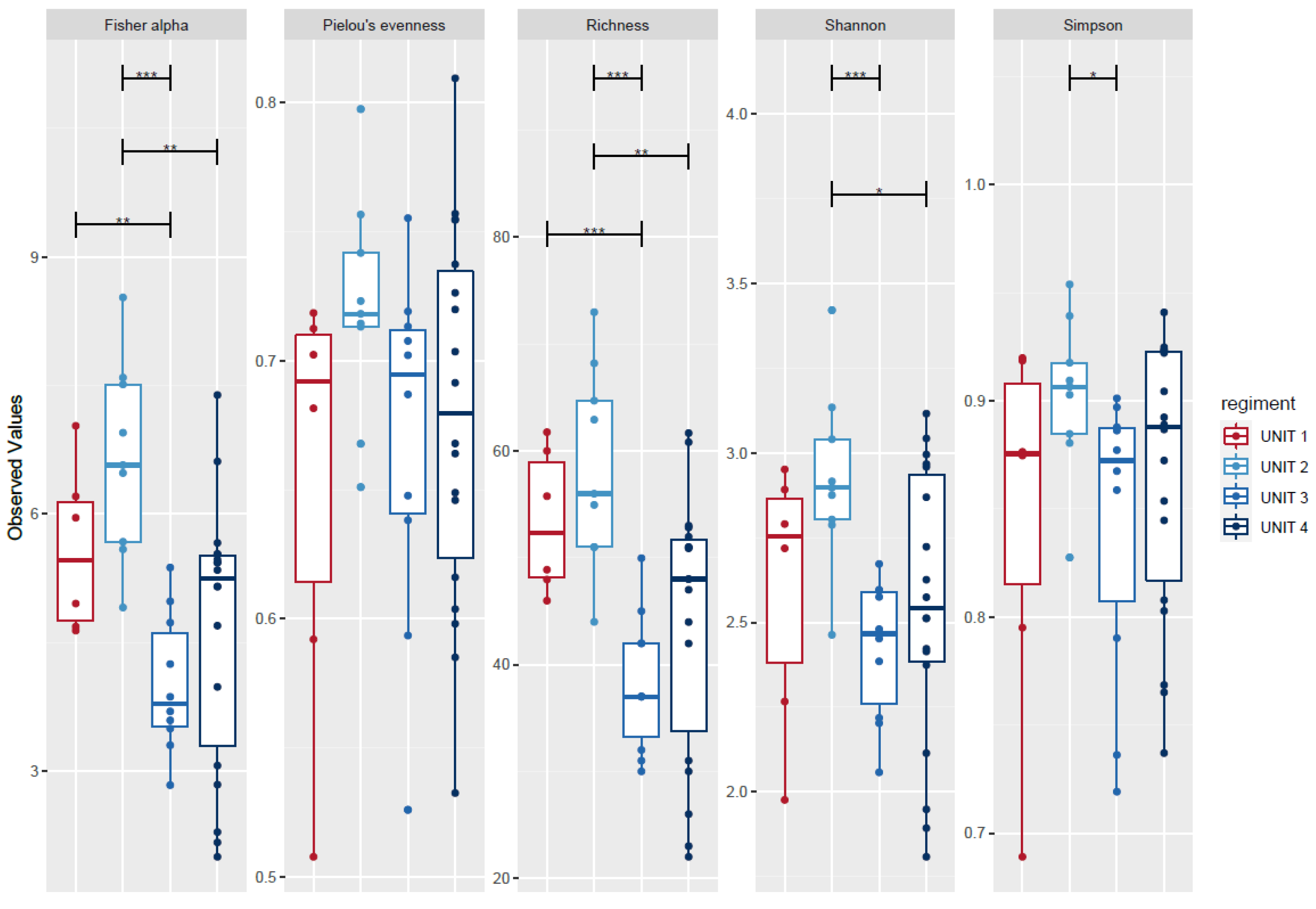
2.3. Microbial Diversity
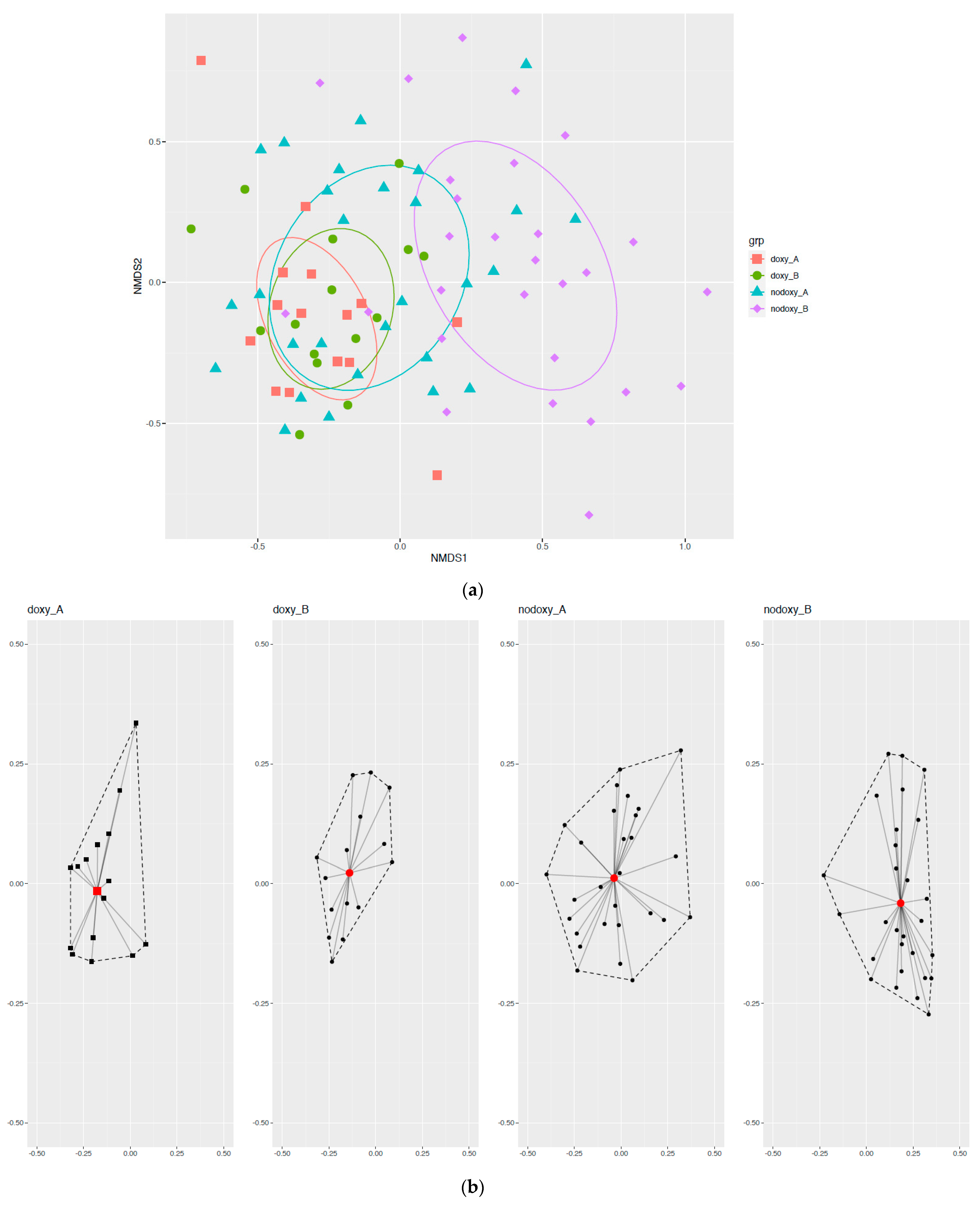
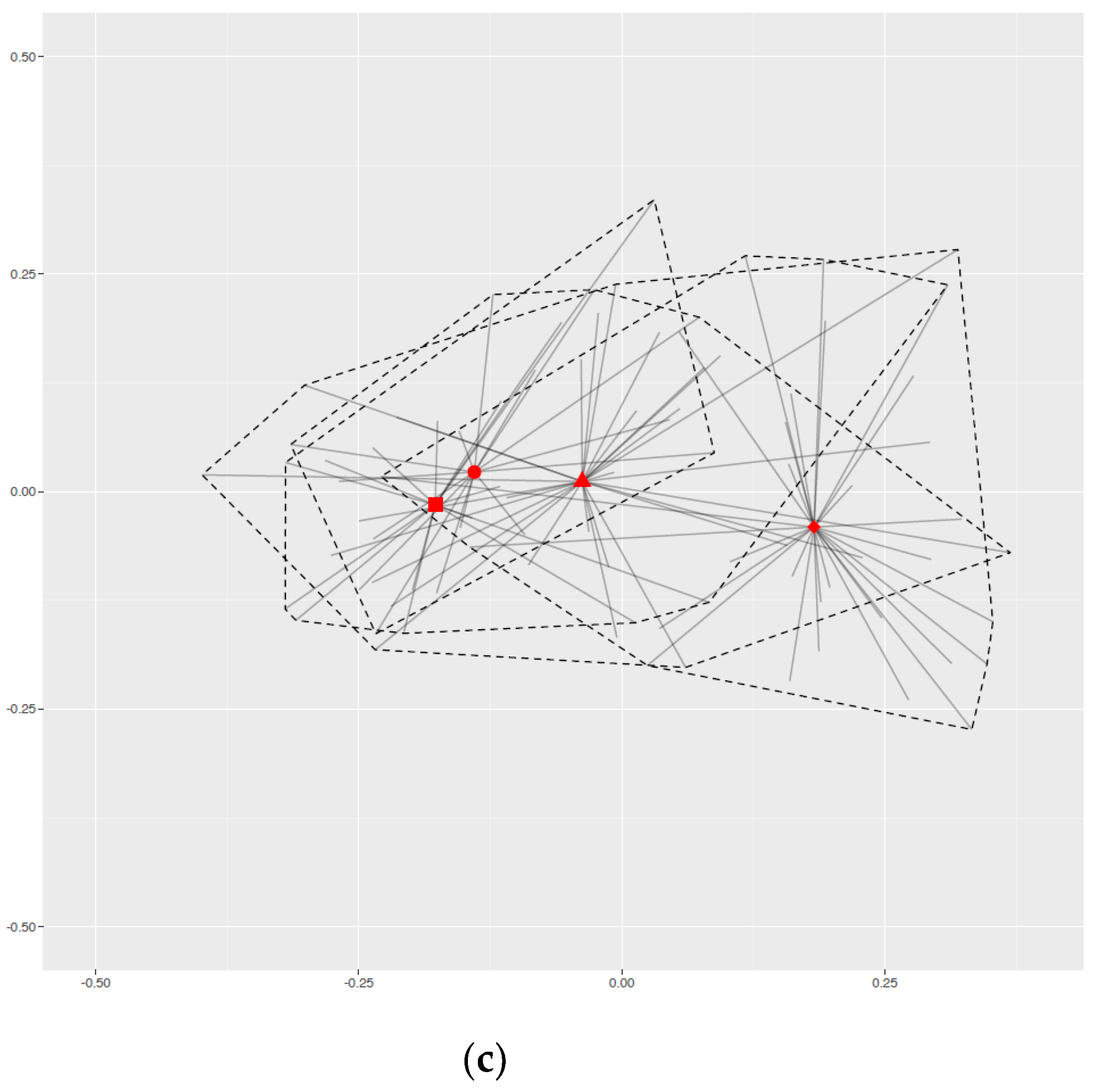
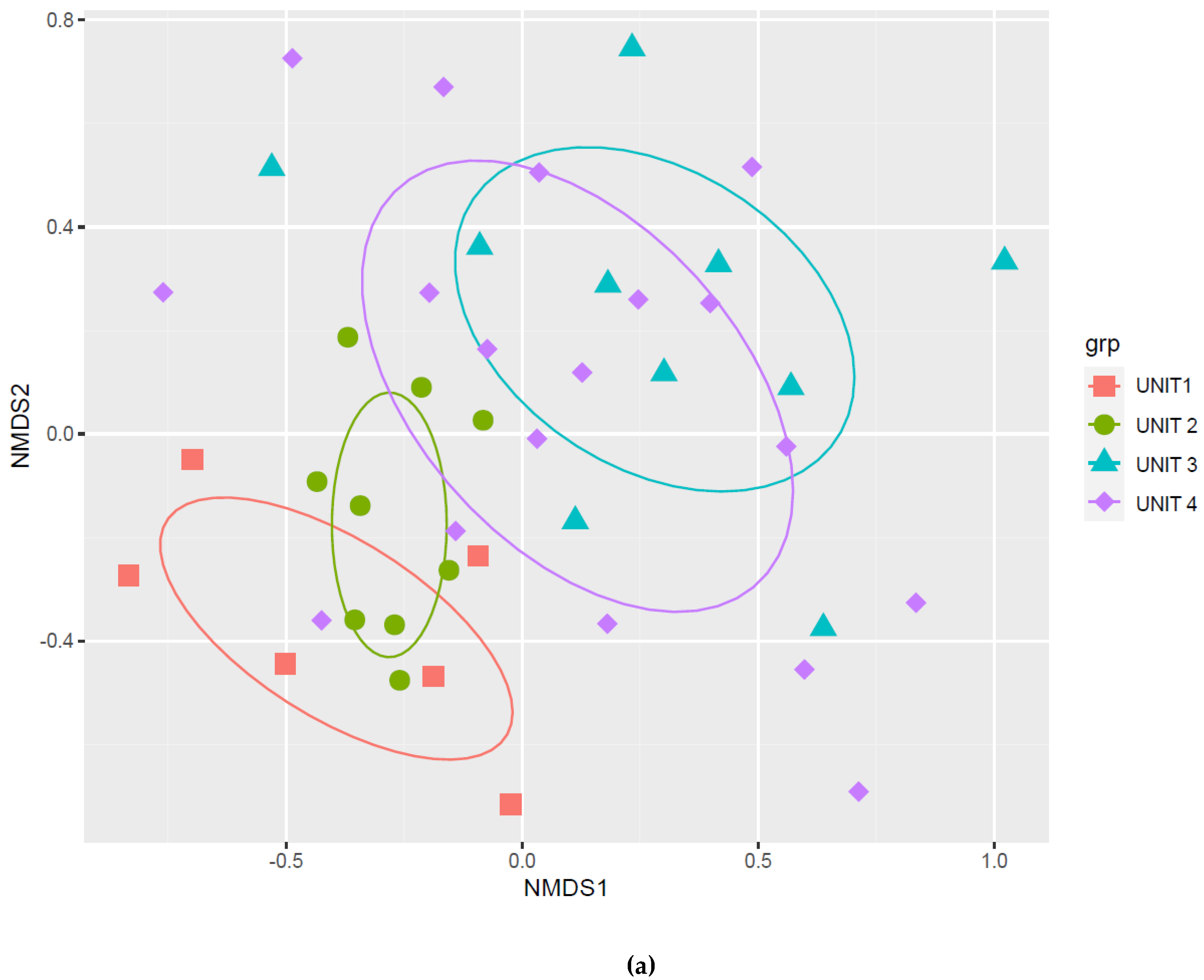
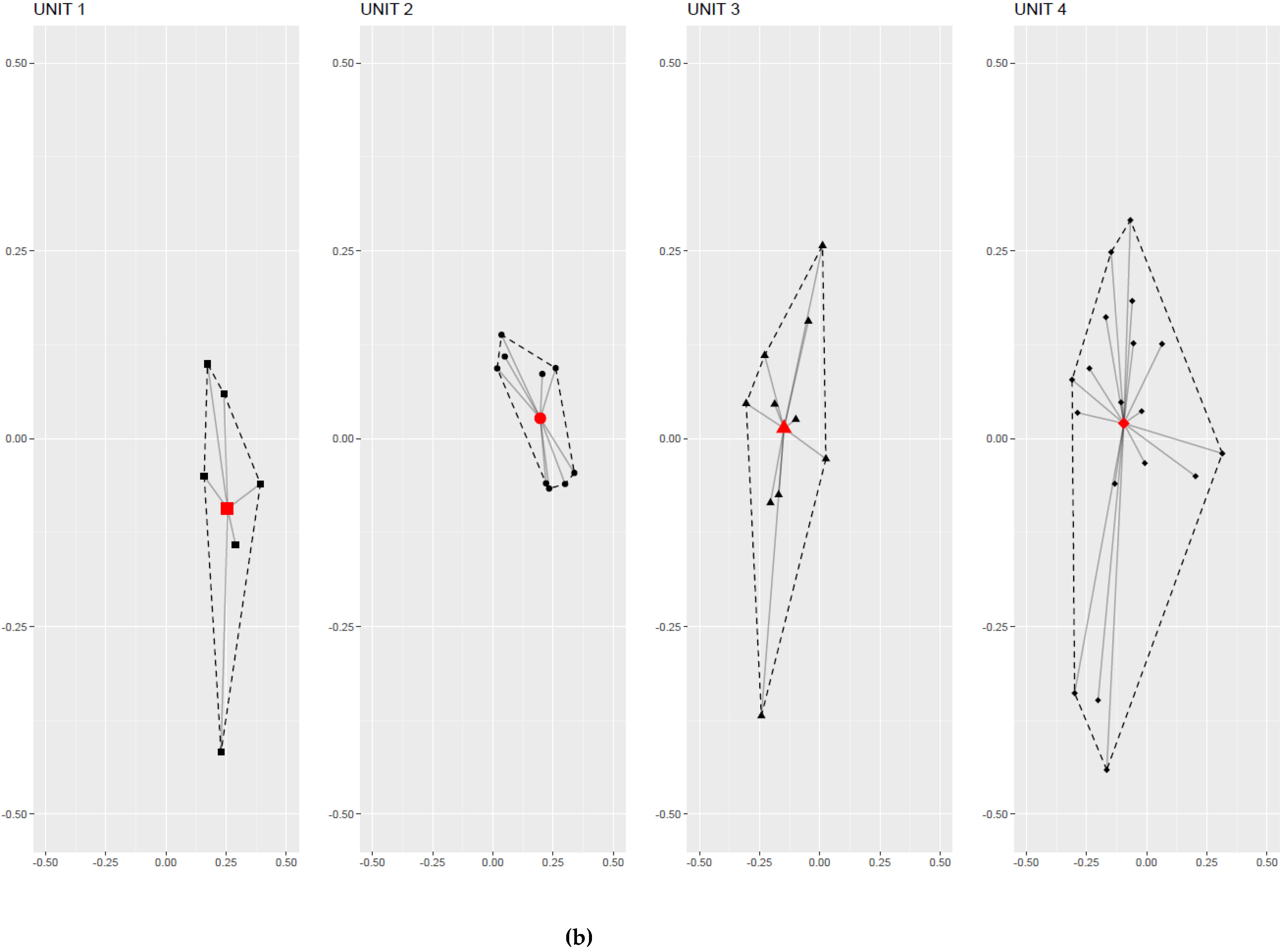
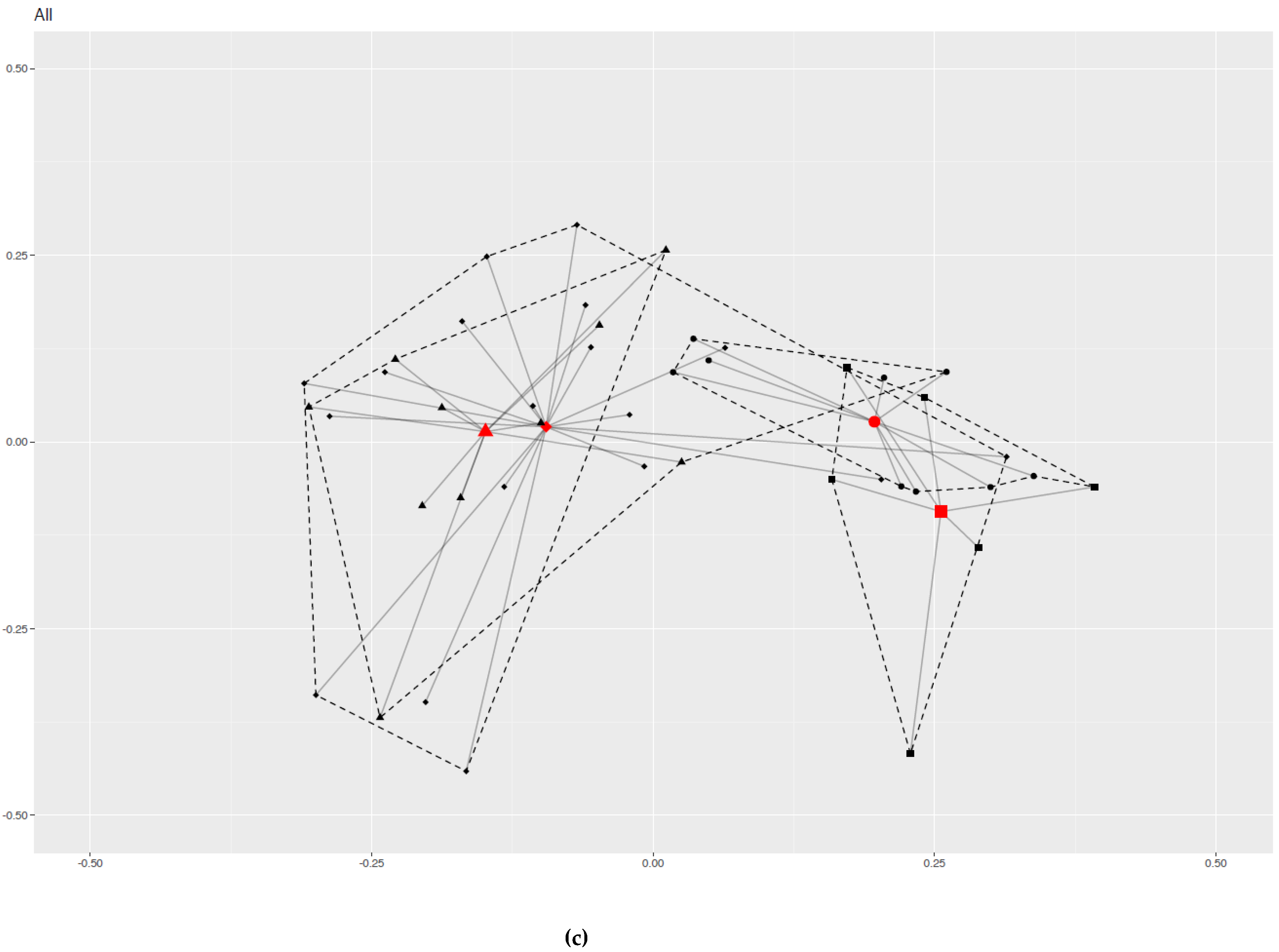
2.4. Group-Effect
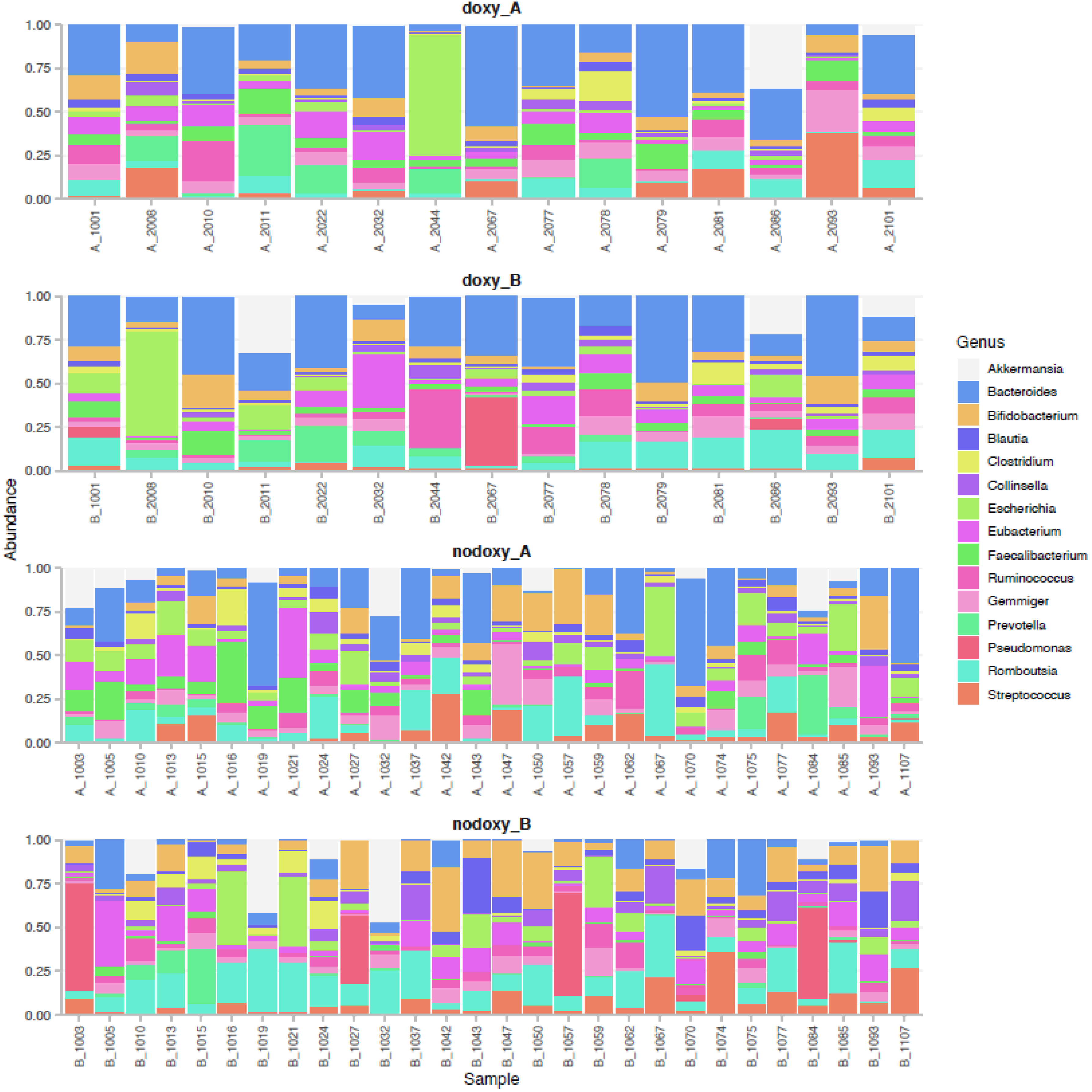
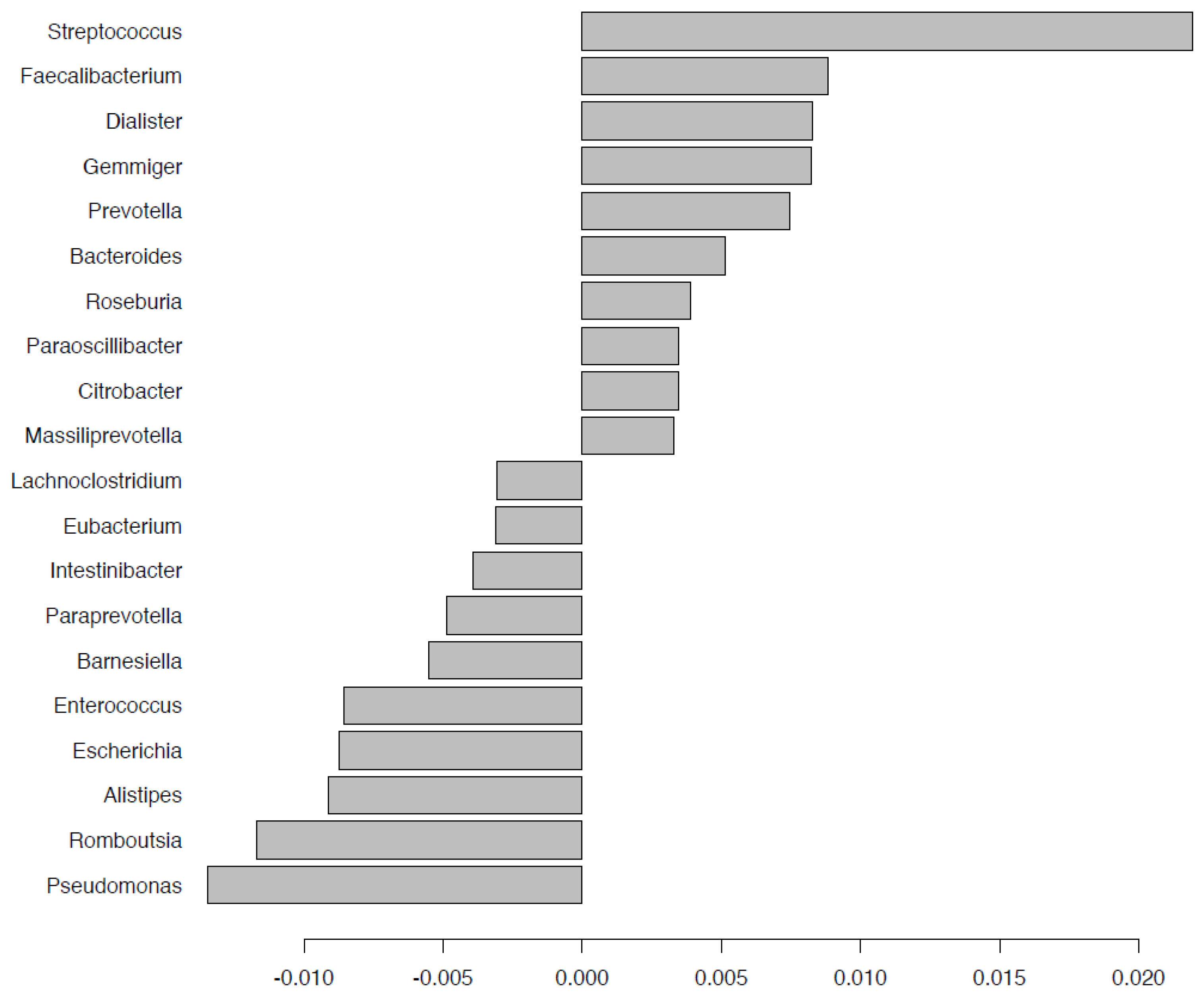
2.5. Fecal Microbiota Composition
2.6. Gut Microbiota and Clinical Data Other Than Doxycycline Exposure
3. Discussion
4. Materials and Methods
4.1. Study Design and Population
4.2. Clinical Data Collection
4.3. Sample Collection and Storage
4.4. DNA Extraction
4.5. Metagenomic Sequencing
4.6. Metagenomic Bioinformatics
4.6.1. Reads Analysis
4.6.2. Clusterization and Taxonomic Assignment
4.6.3. Filters and Corrections
4.7. Metagenomic Bioanalysis
4.8. Statistical Analysis
4.9. Ethics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NIH HMP Working Group; Peterson, J.; Garges, S.; Giovanni, M.; McInnes, P.; Wang, L.; Schloss, J.A.; Bonazzi, V.; McEwen, J.E.; Wetterstrand, K.A.; et al. The NIH Human Microbiome Project. Genome Res. 2009, 19, 2317–2323. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and Evolutionary Forces Shaping Microbial Diversity in the Human Intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef] [Green Version]
- Thuny, F.; Richet, H.; Casalta, J.-P.; Angelakis, E.; Habib, G.; Raoult, D. Vancomycin Treatment of Infective Endocarditis Is Linked with Recently Acquired Obesity. PLoS ONE 2010, 5, e9074. [Google Scholar] [CrossRef] [Green Version]
- Modi, S.R.; Collins, J.J.; Relman, D.A. Antibiotics and the gut microbiota. J. Clin. Investig. 2014, 124, 4212–4218. [Google Scholar] [CrossRef] [Green Version]
- Stirling, A. A general framework for analysing diversity in science, technology and society. J. R. Soc. Interface 2007, 4, 707–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.R.; Magill, A.J.; Parise, M.E.; Arguin, P.M. Doxycycline for Malaria Chemoprophylaxis and Treatment: Report from the CDC Expert Meeting on Malaria Chemoprophylaxis. Am. J. Trop. Med. Hyg. 2011, 84, 517–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heimdahl, A.; Nord, C.E. Influence of doxycycline on the normal human flora and colonization of the oral cavity and colon. Scand. J. Infect. Dis. 1983, 15, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.-U.; Panagiotidis, G.; Bäckström, T.; Weintraub, A.; Nord, C.E. Ecological impact of doxycycline at low dose on normal oropharyngeal and intestinal microflora. Int. J. Antimicrob. Agents 2013, 41, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Lago, K.; Telu, K.; Tribble, D.; Ganesan, A.; Kunz, A.; Geist, C.; Fraser, J.; Mitra, I.; Lalani, T.; Yun, H.C.; et al. Doxycycline Malaria Prophylaxis Impact on Risk of Travelers’ Diarrhea among International Travelers. Am. J. Trop. Med. Hyg. 2020, 103, 1864–1870. [Google Scholar] [CrossRef]
- Elvers, K.T.; Wilson, V.J.; Hammond, A.; Duncan, L.; Huntley, A.L.; Hay, A.D.; van der Werf, E.T. Antibiotic-induced changes in the human gut microbiota for the most commonly prescribed antibiotics in primary care in the UK: A systematic review. BMJ Open 2020, 10, e035677. [Google Scholar] [CrossRef] [PubMed]
- Mättö, J.; Maukonen, J.; Alakomi, H.-L.; Suihko, M.-L.; Saarela, M. Influence of oral doxycycline therapy on the diversity and antibiotic susceptibility of human intestinal bifidobacterial population. J. Appl. Microbiol. 2008, 105, 279–289. [Google Scholar] [CrossRef]
- Angelakis, E.; Million, M.; Kankoe, S.; Lagier, J.-C.; Armougom, F.; Giorgi, R.; Raoult, D. Abnormal weight gain and gut microbiota modifications are side effects of long-term doxycycline and hydroxychloroquine treatment. Antimicrob. Agents Chemother. 2014, 58, 3342–3347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boynton, F.D.D.; Ericsson, A.C.; Uchihashi, M.; Dunbar, M.L.; Wilkinson, J.E. Doxycycline induces dysbiosis in female C57BL/6NCrl mice. BMC Res. Notes 2017, 10, 644. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.Y.; Ahn, Y.; Khare, S.; Gokulan, K.; Piñeiro, S.A.; Cerniglia, C.E. An in vitro study to assess the impact of tetracycline on the human intestinal microbiome. Anaerobe 2018, 49, 85–94. [Google Scholar] [CrossRef]
- Julious, S.A.; Mullee, M.A. Confounding and Simpson’s paradox. BMJ 1994, 309, 1480–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, K.H.; Brown, N.J.; Pelecanos, A.; Brown, A.F. Simpson’s paradox: A statistician’s case study. Emerg. Med. Australas. 2018, 30, 431–433. [Google Scholar] [CrossRef]
- Julian, L.J. Measures of Anxiety. Arthritis Care Res. 2011, 63, S467–S472. [Google Scholar] [CrossRef] [Green Version]
- Chao, A. Nonparametric Estimation of the Number of Classes in a Population. Scand. J. Stat. 1984, 11, 265–270. Available online: https://www.jstor.org/stable/4615964 (accessed on 21 April 2021).
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef]
- Ioannidis, J.P.A. Why Most Published Research Findings Are False. PLOS Med. 2005, 2, e124. [Google Scholar] [CrossRef] [Green Version]
- Walters, W.A.; Reyes, F.; Soto, G.M.; Reynolds, N.D.; Fraser, J.A.; Aviles, R.; Tribble, D.R.; Irvin, A.P.; Kelley-Loughnane, N.; Gutierrez, R.L.; et al. Epidemiology and associated microbiota changes in deployed military personnel at high risk of traveler’s diarrhea. PLoS ONE 2020, 15, e0236703. [Google Scholar] [CrossRef]
- Walker, C.; Preshaw, P.M.; Novak, J.; Hefti, A.F.; Bradshaw, M.; Powala, C. Long-term treatment with sub-antimicrobial dose doxycycline has no antibacterial effect on intestinal flora. J. Clin. Periodontol. 2005, 32, 1163–1169. [Google Scholar] [CrossRef]
- Xiong, W.; Wang, Y.; Sun, Y.; Ma, L.; Zeng, Q.; Jiang, X.; Li, A.; Zeng, Z.; Zhang, T. Antibiotic-mediated changes in the fecal microbiome of broiler chickens define the incidence of antibiotic resistance genes. Microbiome 2018, 6, 34. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.M.; Georg, L.K.; Waters, L.C. Laboratory identification of Rothia dentocariosa and its occurrence in human clinical materials. Appl. Microbiol. 1969, 17, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Roth, G.D.; Thurn, A.N. Continued study of oral nocardia. J. Dent. Res. 1962, 41, 1279–1292. [Google Scholar] [CrossRef] [PubMed]
- Ramanan, P.; Barreto, J.N.; Osmon, D.R.; Tosh, P.K. Rothia Bacteremia: A 10-Year Experience at Mayo Clinic, Rochester, Minnesota. J. Clin. Microbiol. 2014, 52, 3184–3189. [Google Scholar] [CrossRef] [Green Version]
- Zamakhchari, M.; Wei, G.; Dewhirst, F.; Lee, J.; Schuppan, D.; Oppenheim, F.G.; Helmerhorst, E.J. Identification of Rothia Bacteria as Gluten-Degrading Natural Colonizers of the Upper Gastro-Intestinal Tract. PLoS ONE 2011, 6, e24455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergan, T.; Bovre, K.; Hovig, B. Reisolation of Micrococcus mucilaginosus Migula 1900. Acta Pathol. Microbiol. Scand. B Microbiol. Immunol. 1970, 78, 85–97. [Google Scholar] [CrossRef]
- Li, Y.; Kawamura, Y.; Fujiwara, N.; Naka, T.; Liu, H.; Huang, X.; Kobayashi, K.; Ezaki, T. Rothia aeria sp. nov., Rhodococcus baikonurensis sp. nov. and Arthrobacter russicus sp. nov., isolated from air in the Russian space laboratory Mir. Int. J. Syst. Evol. Microbiol. 2004, 54, 827–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franconieri, F.; Join-Lambert, O.; Creveuil, C.; Auzou, M.; Labombarda, F.; Aouba, A.; Verdon, R.; de La Blanchardière, A. Rothia spp. infective endocarditis: A systematic literature review. Med. Mal. Infect. 2020, 51, 228–235. [Google Scholar] [CrossRef]
- Sung, J.J.Y.; Coker, O.O.; Chu, E.; Szeto, C.H.; Luk, S.T.Y.; Lau, H.C.H.; Yu, J. Gastric microbes associated with gastric inflammation, atrophy and intestinal metaplasia 1 year after Helicobacter pylori eradication. Gut 2020, 69, 1572–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uranga, C.C.; Arroyo, P.; Duggan, B.M.; Gerwick, W.H.; Edlund, A. Commensal Oral Rothia mucilaginosa Produces Enterobactin, a Metal-Chelating Siderophore. mSystems 2020, 5, e00161-20. [Google Scholar] [CrossRef]
- Wei, D.; Matsukuma, K. Doxycycline-Related Gastric Mucosal Injury: A Characteristic Histopathologic Pattern. Am. J. Clin. Pathol. 2018, 150, S26. [Google Scholar] [CrossRef]
- Tamanai-Shacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Roseburia spp.: A marker of health? Future Microbiol. 2017, 12, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.; Schwab, C.; Milinovich, G.; Reichert, J.; Ben Mahfoudh, K.; Decker, T.; Engel, M.; Hai, B.; Hainzl, E.; Heider, S.; et al. Phylotype-level 16S rRNA analysis reveals new bacterial indicators of health state in acute murine colitis. ISME J. 2012, 6, 2091–2106. [Google Scholar] [CrossRef] [PubMed]
- Duck, L.W.; Walter, M.R.; Novak, J.; Kelly, D.; Tomasi, M.; Cong, Y.; Elson, C.O. Isolation of flagellated bacteria implicated in Crohn’s disease. Inflamm. Bowel Dis. 2007, 13, 1191–1201. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.-M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Karl, J.P.; Armstrong, N.J.; McClung, H.L.; Player, R.A.; Rood, J.C.; Racicot, K.; Soares, J.W.; Montain, S.J. A diet of U.S. military food rations alters gut microbiota composition and does not increase intestinal permeability. J. Nutr. Biochem. 2019, 72, 108217. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Sheflin, A.M.; Melby, C.L.; Carbonero, F.; Weir, T.L. Linking dietary patterns with gut microbial composition and function. Gut Microbes 2017, 8, 113–129. [Google Scholar] [CrossRef]
- Karl, J.P.; Hatch, A.M.; Arcidiacono, S.M.; Pearce, S.C.; Pantoja-Feliciano, I.G.; Doherty, L.A.; Soares, J.W. Effects of Psychological, Environmental and Physical Stressors on the Gut Microbiota. Front. Microbiol. 2018, 9, 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karl, J.P.; Margolis, L.M.; Madslien, E.H.; Murphy, N.E.; Castellani, J.W.; Gundersen, Y.; Hoke, A.V.; Levangie, M.W.; Kumar, R.; Chakraborty, N.; et al. Changes in intestinal microbiota composition and metabolism coincide with increased intestinal permeability in young adults under prolonged physiological stress. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G559–G571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Kan, E.M.; Lu, J.; Cao, Y.; Wong, R.K.; Keshavarzian, A.; Wilder-Smith, C.H. Combat-training increases intestinal permeability, immune activation and gastrointestinal symptoms in soldiers. Aliment. Pharmacol. Ther. 2013, 37, 799–809. [Google Scholar] [CrossRef]
- Angelakis, E.; Bachar, D.; Henrissat, B.; Armougom, F.; Audoly, G.; Lagier, J.-C.; Robert, C.; Raoult, D. Glycans affect DNA extraction and induce substantial differences in gut metagenomic studies. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef]
- Zoetendal, E.G.; Booijink, C.C.G.M.; Klaassens, E.S.; Heilig, H.G.H.J.; Kleerebezem, M.; Smidt, H.; de Vos, W.M. Isolation of RNA from bacterial samples of the human gastrointestinal tract. Nat. Protoc. 2006, 1, 954–959. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; Mcveigh, R.; O’Neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A comprehensive update on curation, resources and tools. Database 2020, 2020, baaa062. [Google Scholar] [CrossRef] [PubMed]
- RPubs-Microbiota Analysis in R. Available online: https://rpubs.com/dillmcfarlan/R_microbiotaSOP (accessed on 5 April 2021).
- Josse, J.; Husson, F. missMDA: A Package for Handling Missing Values in Multivariate Data Analysis. J. Stat. Softw. 2016, 70, 1–31. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Balmford, A.; Jayasuriya, A.H.M.; Green, M.J.B. Using higher-taxon richness as a surrogate for species richness: II. Local applications. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1996, 263, 1571–1575. [Google Scholar] [CrossRef]
- Williams, P.H.; Gaston, K.J. Measuring more of biodiversity: Can higher-taxon richness predict wholesale species richness? Biol. Conserv. 1994, 67, 211–217. [Google Scholar] [CrossRef]
- Fisher, R.A.; Corbet, A.S.; Williams, C.B. The Relation Between the Number of Species and the Number of Individuals in a Random Sample of an Animal Population. J. Anim. Ecol. 1943, 12, 42–58. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef]
- Shade, A.; Handelsman, J. Beyond the Venn diagram: The hunt for a core microbiome. Environ. Microbiol. 2012, 14, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Salonen, A.; Salojärvi, J.; Lahti, L.; Vos, W.M. de The adult intestinal core microbiota is determined by analysis depth and health status. Clin. Microbiol. Infect. 2012, 18, 16–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strahler, A.H. Binary Discriminant Analysis: A New Method for Investigating Species-Environment Relationships. Ecology 1978, 59, 108–116. [Google Scholar] [CrossRef]
- Cáceres, M.D.; Font, X.; Oliva, F. Assessing species diagnostic value in large data sets: A comparison between phi-coefficient and Ochiai index. J. Veg. Sci. 2008, 19, 779–788. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | NODOXY n = 28 | DOXY n = 15 | p-Value |
|---|---|---|---|
| Military units | <0.001 | ||
| Unit 1 | - | 6 (40) | |
| Unit 2 | - | 9 (60) | |
| Unit 3 | 10 (36) | - | |
| Unit 4 | 18 (64) | - | |
| Mission locations | <0.001 | ||
| Mali | - | 28 (100) | |
| Iraq | 18 (64) | - | |
| Lebanon | 10 (36 | - | |
| Age, years, median (IQR) | 29.5 (22.7–34.7) | 30 (27–34) | 0.6 |
| Sex, F/M | 2/26 | 0/15 | 0.56 |
| BMI class | 1 | ||
| Normal < 25 | 16 (57) | 9 (60) | |
| Overweight ≥ 25 | 11 (39) | 5 (33) | |
| Obesity > 30 | 1 (4) | 1 (7) | |
| Military rank, manager | 13 (46) | 8 (53) | 0.75 |
| Marital status, married or attached | 15 (54) | 9 (60) | 0.75 |
| Active smoker | 12 (43) | 7 (47) | 0.75 |
| Probiotics intake | 1 | ||
| Yes | 4 (14) | 2 (13) | |
| No | 16 (57) | 9 (60) | |
| Unknown | 8 (29) | 4 (27 | |
| Baseline sport, hours/week, median (IQR) | 6 (3–8.25) | 6 (5–9) | 0.60 |
| Sport in mission, hours/week, median (IQR) | 5 (2–8.25) | 7 (4–10) | 0.15 |
| Accommodation during mission, urban/rural | 11/17 | 5/10 | 0.75 |
| Diarrhea during mission | 16 (57) | 5 (33) | 0.20 |
| Sedentary activity during mission | 7 (25) | 1 (7) | 0.23 |
| Change in food habits during mission | 23 (82) | 10 (67) | 0.28 |
| Percentage of weight variation after mission, kg, median (IQR) | 0 (−3.25–+1.25) | 0 (−1–+1) | 0.49 |
| BMI variation after mission, kg/m2, median (IQR) | 0 (−1.05–+0.38) | 0 (−0.3–+0.3) | 0.48 |
| Baseline STAI score (0–80), median (IQR) | 44.5 (42.5–46.25) | 46 (43–49) | 0.17 |
| Return STAI score (0–80), median (IQR) | 45 (41–47) | 44 (43–48.5) | 0.19 |
| Variables | NODOXY | DOXY | ||||
|---|---|---|---|---|---|---|
| Before Mission n = 28 | After Mission n = 28 | p-Value | Before Mission n = 15 | After Mission n = 15 | p-Value | |
| Weight, kg, median (IQR) | 72.5 (68–81) | 72 (67.2–83) | 0.79 | 76 (70–81.5) | 74 (70.5–80) | 0.95 |
| BMI, kg/m2, median (IQR) | 24.4 (22.9–26.8) | 24.1 (22.9–26.2) | 0.73 | 24.5 (23.5–25.6) | 24.5 (22.8–26.1) | 1 |
| BMI class | 0.71 | 1 | ||||
| Normal < 25 | 16 (57) | 16 (57) | 9 (60) | 9 (60) | ||
| Overweight ≥ 25 | 11 (39) | 11 (39) | 5 (33) | 5 (33) | ||
| Obesity > 30 | 1 (4) | 1 (4) | 1 (7) | 1 (7) | ||
| Sport, hours/week, median (IQR) | 6 (3–8.25) | 5 (2–8.25) | 0.35 | 6 (5–9) | 7 (4–10) | 0.79 |
| STAI score (0–80), median (IQR) | 44.5 (42.5–46.25) | 45 (41–47) | 0.98 | 46 (43–49) | 44 (43–48.5) | 0.60 |
| Doxy | Nodoxy | |||
|---|---|---|---|---|
| Before | After | Before | After | |
| Alistipes | - | X | X | - |
| Bacteroides | X | X | X | X |
| Bifidobacterium | X | X | X | X |
| Blautia | X | X | X | X |
| Citrobacter | X | - | - | - |
| Clostridium | X | X | X | X |
| Collinsella | X | X | X | X |
| Dorea | X | X | X | X |
| Escherichia | X | X | X | X |
| Eubacterium | X | X | X | X |
| Faecalibacterium | X | X | X | X |
| Fusicatenibacter | X | X | X | X |
| Gemmiger | X | X | X | X |
| Guyana | X | X | X | X |
| Intestinibacter | X | - | - | X |
| Lachnoclostridium | - | X | X | - |
| Parabacteroides | - | X | X | X |
| Romboutsia | X | X | X | X |
| Roseburia | - | X | X | - |
| Ruminococcus | X | X | X | X |
| Senegalimassilia | X | X | X | X |
| Streptococcus | X | X | X | X |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Javelle, E.; Mayet, A.; Million, M.; Levasseur, A.; Allodji, R.S.; Marimoutou, C.; Lavagna, C.; Desplans, J.; Fournier, P.E.; Raoult, D.; et al. Gut Microbiota in Military International Travelers with Doxycycline Malaria Prophylaxis: Towards the Risk of a Simpson Paradox in the Human Microbiome Field. Pathogens 2021, 10, 1063. https://doi.org/10.3390/pathogens10081063
Javelle E, Mayet A, Million M, Levasseur A, Allodji RS, Marimoutou C, Lavagna C, Desplans J, Fournier PE, Raoult D, et al. Gut Microbiota in Military International Travelers with Doxycycline Malaria Prophylaxis: Towards the Risk of a Simpson Paradox in the Human Microbiome Field. Pathogens. 2021; 10(8):1063. https://doi.org/10.3390/pathogens10081063
Chicago/Turabian StyleJavelle, Emilie, Aurélie Mayet, Matthieu Million, Anthony Levasseur, Rodrigue S. Allodji, Catherine Marimoutou, Chrystel Lavagna, Jérôme Desplans, Pierre Edouard Fournier, Didier Raoult, and et al. 2021. "Gut Microbiota in Military International Travelers with Doxycycline Malaria Prophylaxis: Towards the Risk of a Simpson Paradox in the Human Microbiome Field" Pathogens 10, no. 8: 1063. https://doi.org/10.3390/pathogens10081063
APA StyleJavelle, E., Mayet, A., Million, M., Levasseur, A., Allodji, R. S., Marimoutou, C., Lavagna, C., Desplans, J., Fournier, P. E., Raoult, D., & Texier, G. (2021). Gut Microbiota in Military International Travelers with Doxycycline Malaria Prophylaxis: Towards the Risk of a Simpson Paradox in the Human Microbiome Field. Pathogens, 10(8), 1063. https://doi.org/10.3390/pathogens10081063