
Investigation of the Genomic and Pathogenic Features of the Potentially Zoonotic Streptococcus parasuis

 ,
, 
Abstract
:1. Introduction
2. Results
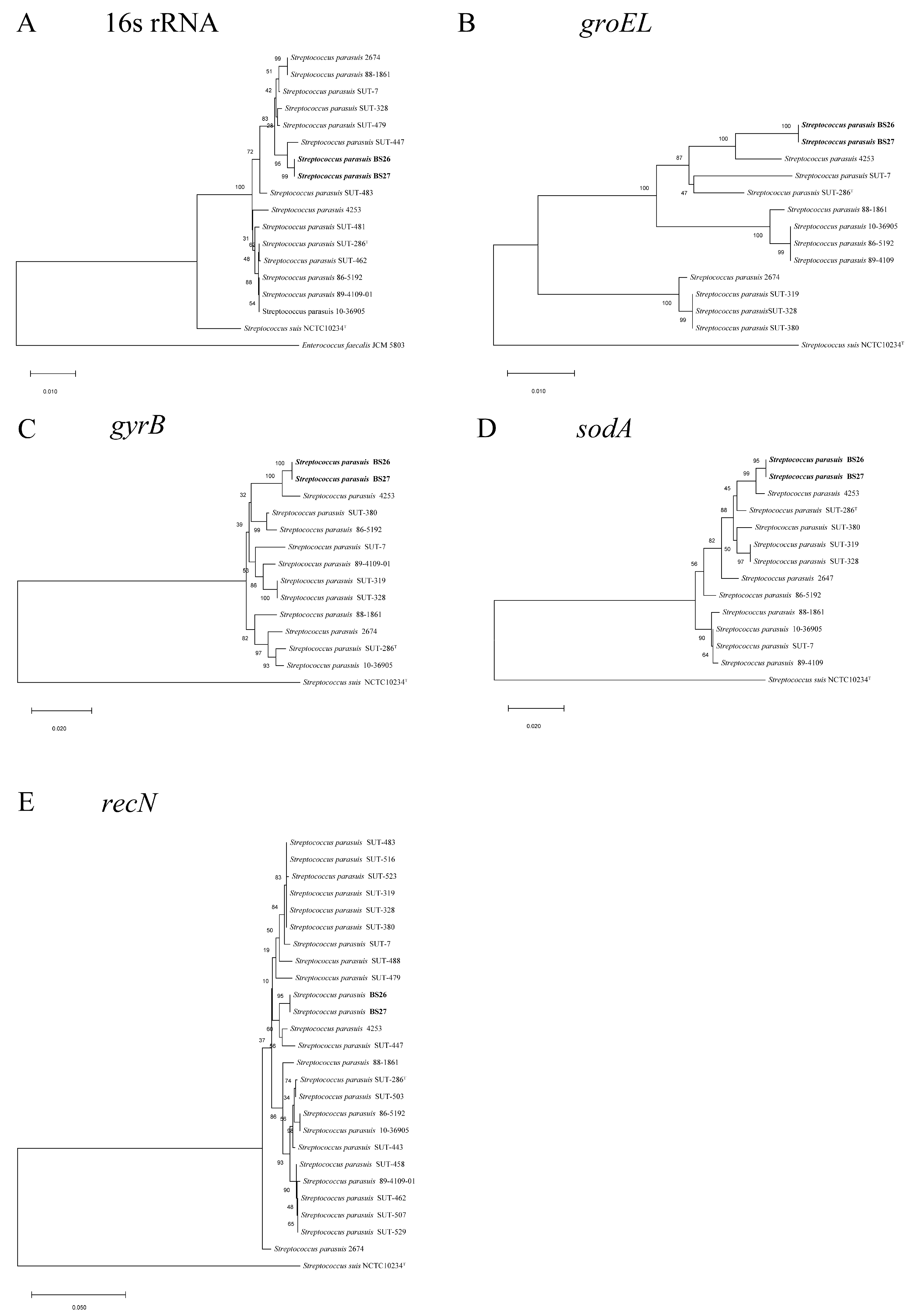
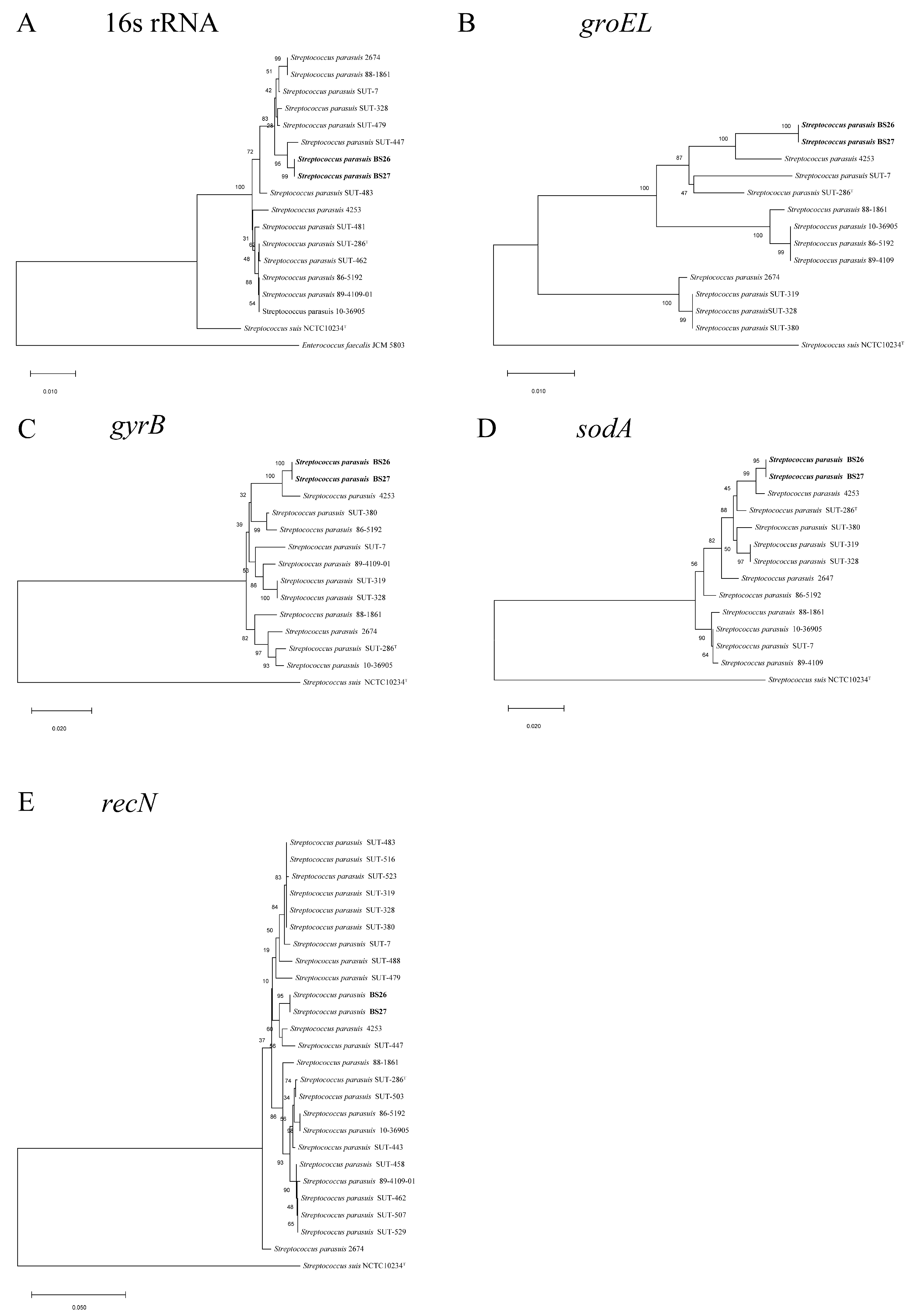
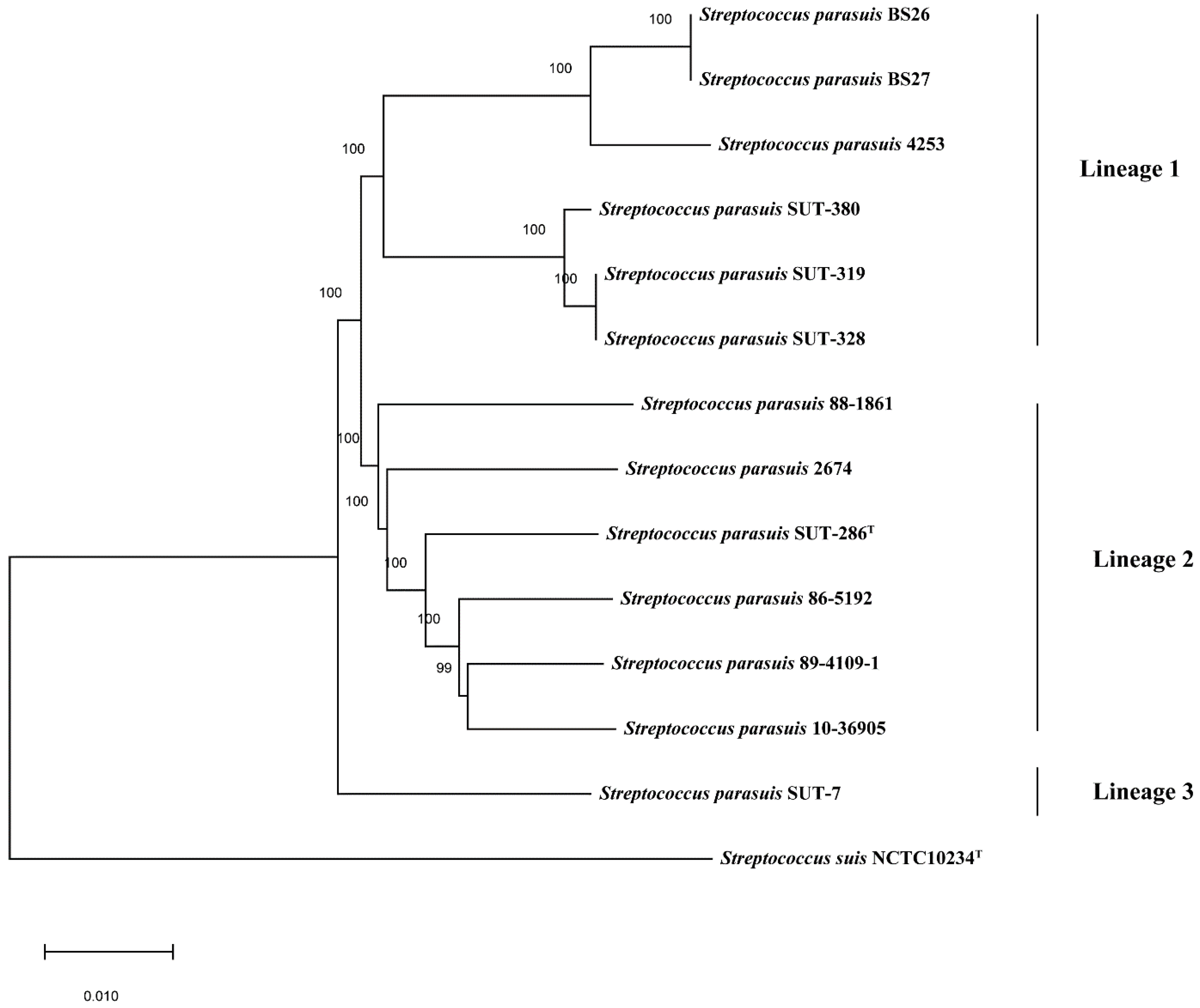
2.1. Phylogenetic Analysis of Full-Length 16S rRNA and Housekeeping Genes groEL, gyrB, sodA, and recN of S. parasuis
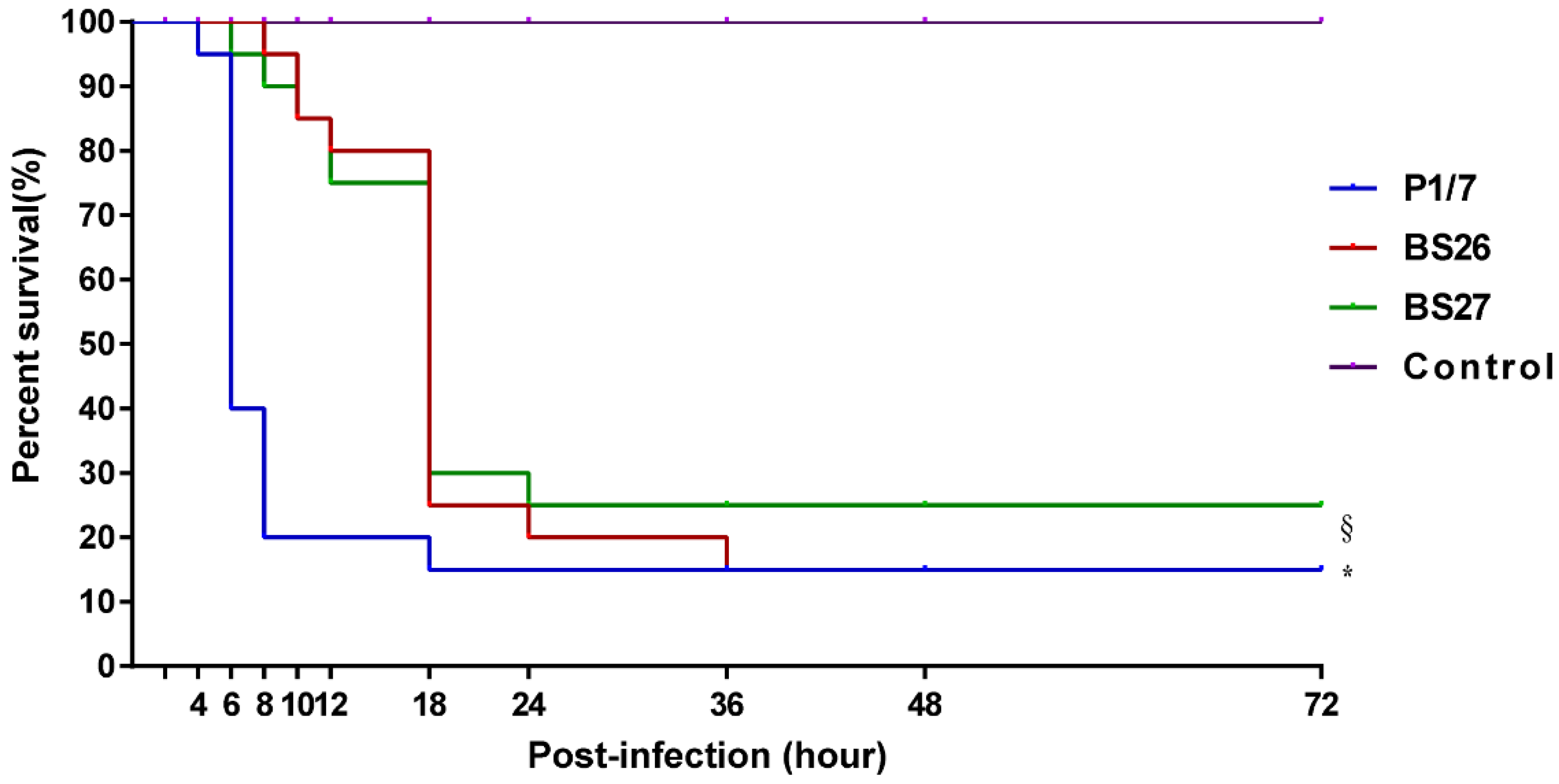
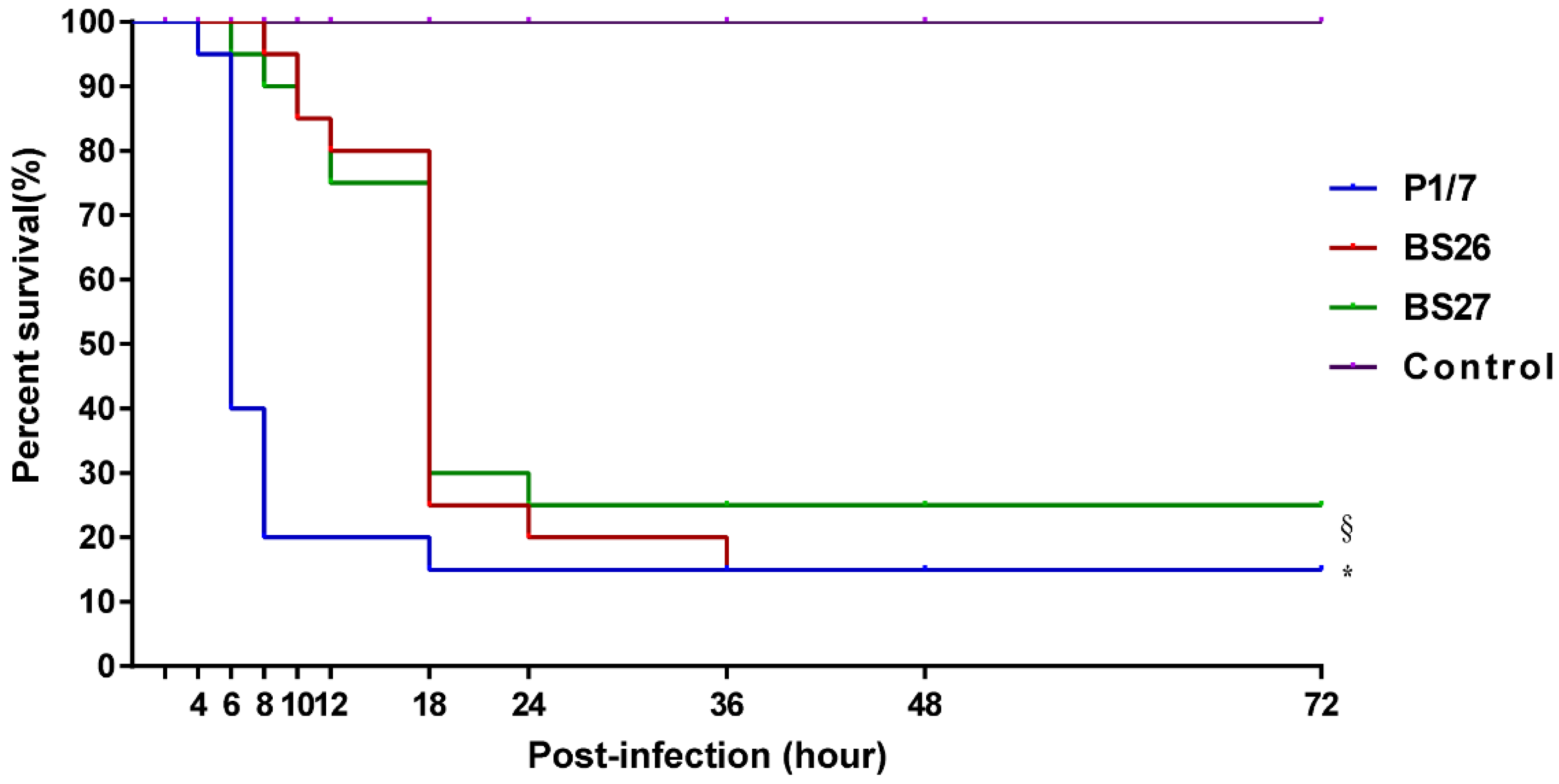
2.2. Difference in Virulence between S. suis Strain P1/7 and Two S. parasuis Strains
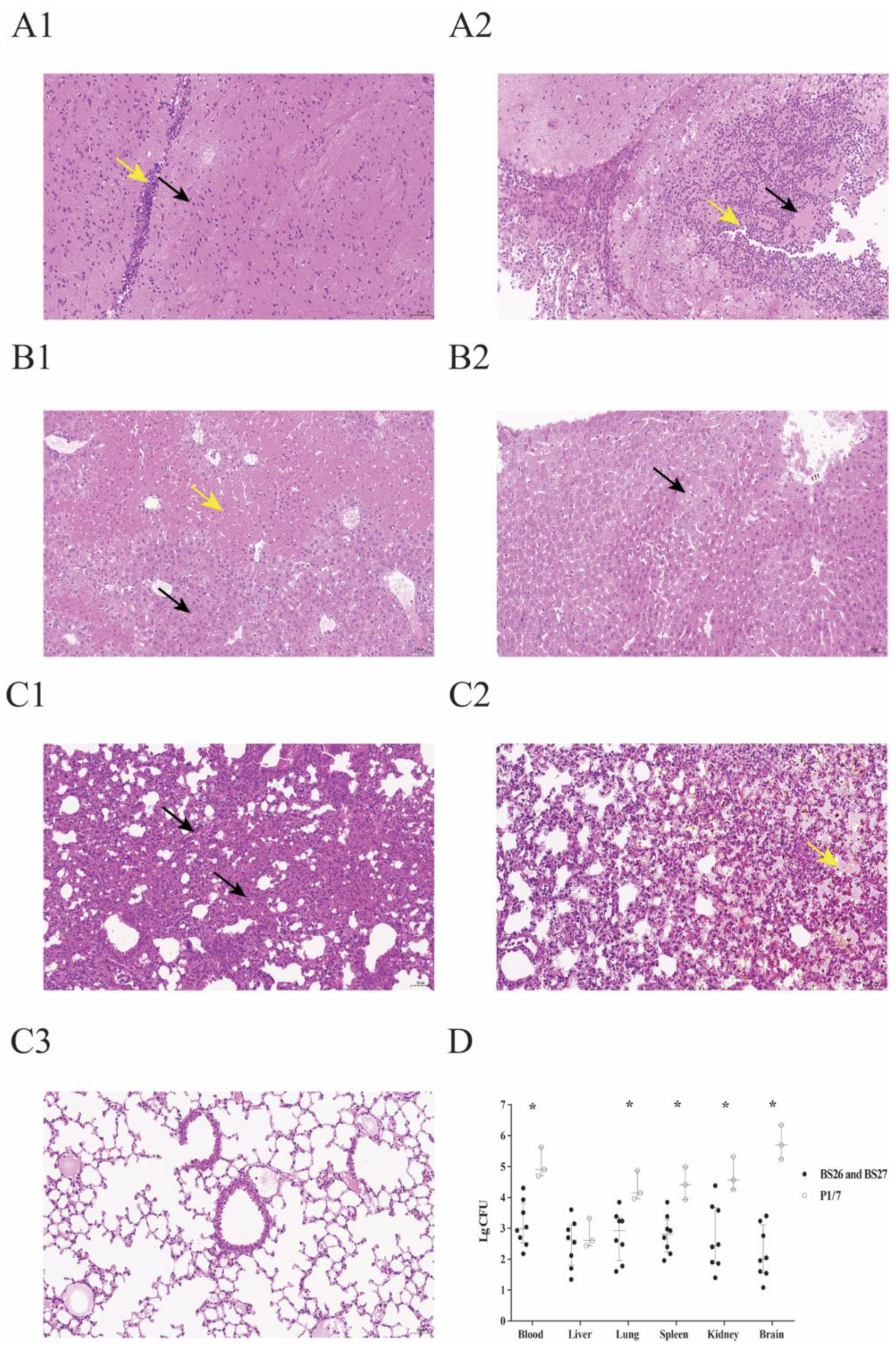
2.3. Histopathological Lesions and Bacterial Load in Survival Mice
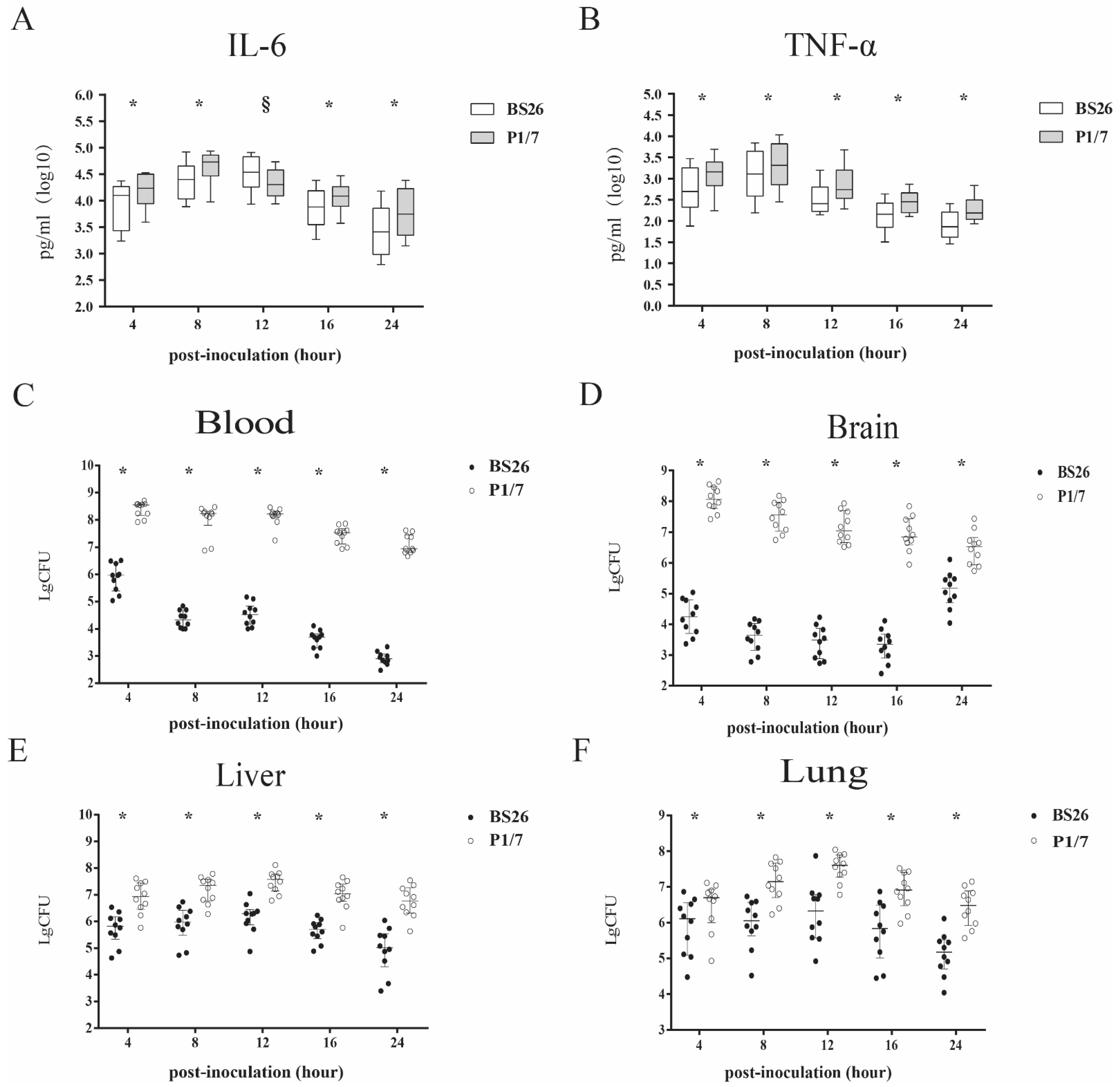
2.4. Pro-Inflammatory Cytokine Production and Bacterial Loads in Mice Infected with S. parasuis Strain BS26 and S. parasuis Strain P1/7
2.5. AR Genes and Antimicrobial Susceptibility Profiles of BS26 and BS27
2.6. Comparative Genomic Analysis of S. parasuis
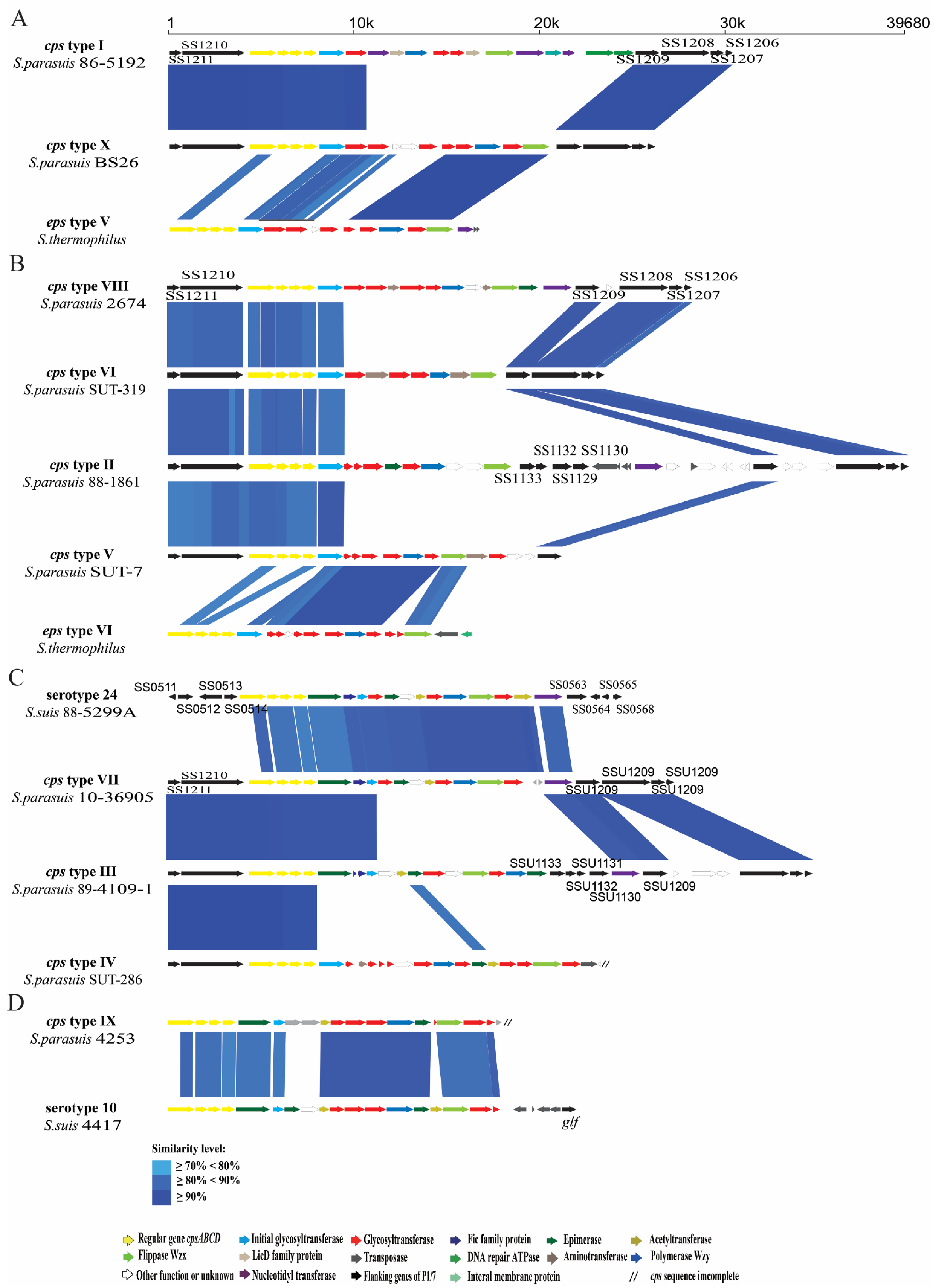
2.7. The Analysis of Cps Gene Clusters of S. parasuis
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Case Description
4.2. DNA Extraction, Sequencing, and Bioinformatic Analysis
4.3. Phylogenetic Analysis of 16S RNA and Housekeeping Genes
4.4. Experimental Infection
4.4.1. Survival Assay
4.4.2. Histopathological Analysis and Bacterial Loads in Survival Mice
4.4.3. In Vivo Cytokine Production and Measurement of Bacterial Loads
4.5. Detection of Antibiotic Resistance (AR) Determinants and Antimicrobial Susceptibility Profiles
4.6. Comparative Genomic Analysis of S. parasuis
4.7. Analysis of cps Loci
4.8. Statistics
4.9. Nucleotide Sequence Accession Numbers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microbes Infect. 2014, 3, e45. [Google Scholar] [CrossRef]
- Gajdács, M.; Németh, A.; Knausz, M.; Barrak, I.; Stájer, A.; Mestyán, G.; Melegh, S.; Nyul, A.; Tóth, Á.; Ágoston, Z.; et al. Streptococcus suis: An Underestimated Emerging Pathogen in Hungary? Microorganisms 2020, 8, 1292. [Google Scholar] [CrossRef] [PubMed]
- Ágoston, Z.; Terhes, G.; Hannauer, P.; Gajdács, M.; Urbán, E. Fatal case of bacteremia caused by Streptococcus suis in a splenectomized man and a review of the European literature. Acta Microbiol. Immunol. Hung. 2020, 67, 148–155. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.E.; Gottschalk, M.; Brousseau, R.; Harel, J.; Hemmingsen, S.M.; Goh, S.H. Biochemical analysis, cpn60 and 16S rDNA sequence data indicate that Streptococcus suis serotypes 32 and 34, isolated from pigs, are Streptococcus orisratti. Vet. Microbiol. 2005, 107, 63–69. [Google Scholar] [CrossRef]
- Nomoto, R.; Maruyama, F.; Ishida, S.; Tohya, M.; Sekizaki, T.; Osawa, R. Reappraisal of the taxonomy of Streptococcus suis serotypes 20, 22 and 26: Streptococcus parasuis sp. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Yamada, R.; Tien, L.H.T.; Arai, S.; Tohya, M.; Ishida-Kuroki, K.; Nomoto, R.; Kim, H.; Suzuki, E.; Osawa, R.; Watanabe, T.; et al. Development of PCR for identifying Streptococcus parasuis, a close relative of Streptococcus suis. J. Vet. Med. Sci. 2018, 80, 1101–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottschalk, M.; Lacouture, S. Canada: Distribution of Streptococcus suis (from 2012 to 2014) and Actinobacillus pleuropneumoniae (from 2011 to 2014) serotypes isolated from diseased pigs. Can. Vet. J. 2015, 56, 1093–1094. [Google Scholar]
- Gottschalk, M.; Lacouture, S.; Bonifait, L.; Roy, D.; Fittipaldi, N.; Grenier, D. Characterization of Streptococcus suis isolates recovered between 2008 and 2011 from diseased pigs in Québec, Canada. Vet. Microbiol. 2013, 162, 819–825. [Google Scholar] [CrossRef]
- Wei, Z.; Li, R.; Zhang, A.; He, H.; Hua, Y.; Xia, J.; Cai, X.; Chen, H.; Jin, M. Characterization of Streptococcus suis isolates from the diseased pigs in China between 2003 and 2007. Vet. Microbiol. 2009, 137, 196–201. [Google Scholar] [CrossRef]
- Gottschalk, M.; Higgins, R.; Jacques, M.; Beaudoin, M.; Henrichsen, J. Characterization of six new capsular types (23 through 28) of Streptococcus suis. J. Clin. Microbiol. 1991, 29, 2590–2594. [Google Scholar] [CrossRef] [Green Version]
- Holden, M.T.G.; Hauser, H.; Sanders, M.; Ngo, T.H.; Cherevach, I.; Cronin, A.; Goodhead, I.; Mungall, K.; Quail, M.A.; Price, C.; et al. Rapid Evolution of Virulence and Drug Resistance in the Emerging Zoonotic Pathogen Streptococcus suis. PLoS ONE 2009, 4, e6072. [Google Scholar] [CrossRef] [Green Version]
- Okura, M.; Osaki, M.; Nomoto, R.; Arai, S.; Osawa, R.; Sekizaki, T.; Takamatsu, D. Current Taxonomical Situation of Streptococcus suis. Pathogens 2016, 5, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, L.; Lu, S.; Hu, S.; Jin, D.; Lai, X.; Yang, J.; Chen, C.; Wang, Y.; Wang, Y.; Bai, X.; et al. Streptococcus marmotae sp. nov., isolated from the respiratory tract of Marmota himalayana. Int. J. Syst. Evol. Microbiol. 2016, 66, 4315–4322. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Huang, W.; Xie, H.; Ye, C.; Jing, H.; Ren, Z.; Xu, J. The genetically modified suilysin, rSLYP353L, provides a candidate vaccine that suppresses proinflammatory response and reduces fatality following infection with Streptococcus suis. Vaccine 2013, 31, 4209–4215. [Google Scholar] [CrossRef]
- Domínguez-Punaro, M.C.; Segura, M.; Plante, M.-M.; Lacouture, S.; Rivest, S.; Gottschalk, M. Streptococcus suis Serotype 2, an Important Swine and Human Pathogen, Induces Strong Systemic and Cerebral Inflammatory Responses in a Mouse Model of Infection. J. Immunol. 2007, 179, 1842–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachance, C.; Gottschalk, M.; Gerber, P.P.; Lemire, P.; Xu, J.; Segura, M. Exacerbated Type II Interferon Response Drives Hypervirulence and Toxic Shock by an Emergent Epidemic Strain of Streptococcus suis. Infect. Immun. 2013, 81, 1928–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okura, M.; Takamatsu, D.; Maruyama, F.; Nozawa, T.; Nakagawa, I.; Osaki, M.; Sekizaki, T.; Gottschalk, M.; Kumagai, Y.; Hamada, S. Genetic Analysis of Capsular Polysaccharide Synthesis Gene Clusters from All Serotypes of Streptococcus suis: Potential Mechanisms for Generation of Capsular Variation. Appl. Environ. Microbiol. 2013, 79, 2796–2806. [Google Scholar] [CrossRef] [Green Version]
- Yarza, P.; Richter, M.; Peplies, J.; Euzeby, J.; Amann, R.; Schleifer, K.-H.; Ludwig, W.; Glöckner, F.O.; Rosselló-Móra, R. The All-Species Living Tree project: A 16S rRNA-based phylogenetic tree of all sequenced type strains. Syst. Appl. Microbiol. 2008, 31, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Du, P.; Zheng, H.; Yu, W.; Wan, L.; Chen, C. Whole-genome sequence comparison as a method for improving bacterial species definition. J. Gen. Appl. Microbiol. 2014, 60, 75–78. [Google Scholar] [CrossRef] [Green Version]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Zheng, H.; Zhang, J.; Jing, H.; Wang, L.; Xiong, Y.; Wang, W.; Zhou, Z.; Sun, Q.; Luo, X.; et al. Clinical, Experimental, and Genomic Differences between Intermediately Pathogenic, Highly Pathogenic, and EpidemicStreptococcus suis. J. Infect. Dis. 2009, 199, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Dofferhoff, A.S.; Vellenga, E.; Limburg, P.C.; Van Zanten, A.; Mulder, P.O.; Weits, J. Tumour necrosis factor (cachectin) and other cytokines in septic shock: A review of the literature. Neth. J. Med. 1991, 39, 45–62. [Google Scholar]
- Norrby-Teglund, A.; Pauksens, K.; Norgren, M.; Holm, S.E. Correlation between Serum TNFα and IL6 levels and Severity of Group: A Streptococcal Infections. Scand. J. Infect. Dis. 1995, 27, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, L.; Cooper, A.; Fantino, C.; Altmann, D.; Sriskandan, S. The Mechanism of Superantigen-Mediated Toxic Shock: Not a Simple Th1 Cytokine Storm. J. Immunol. 2005, 175, 6870–6877. [Google Scholar] [CrossRef]
- Zheng, H.; Ye, C.; Segura, M.; Gottschalk, M.; Xu, J. Mitogenic effect contributes to increased virulence of Streptococcus suis sequence type 7 to cause streptococcal toxic shock-like syndrome. Clin. Exp. Immunol. 2008, 153, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Du, P.; Wang, J.; Lan, R.; Huang, J.; Luo, M.; Jiang, Y.; Zeng, J.; Quan, Y.; Shi, Z.; et al. Genomic Epidemiology of Streptococcus suis Sequence Type 7 Sporadic Infections in the Guangxi Zhuang Autonomous Region of China. Pathogens 2019, 8, 187. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, C.P.; Varaldo, P.E.M.; Facinelli, B.P. Streptococcus suis, an Emerging Drug-Resistant Animal and Human Pathogen. Front. Microbiol. 2011, 2, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Ma, J.; Shang, K.; Hu, X.; Liang, Y.; Li, D.; Wu, Z.; Dai, L.; Chen, L.; Wang, L. Evolution and Diversity of the Antimicrobial Resistance Associated Mobilome in Streptococcus suis: A Probable Mobile Genetic Elements Reservoir for Other Streptococci. Front. Cell. Infect. Microbiol. 2016, 6, 118. [Google Scholar] [CrossRef] [Green Version]
- Yun, K.W.; Cho, E.Y.; Choi, E.H.; Lee, H.J. Capsular polysaccharide gene diversity of pneumococcal serotypes 6A, 6B, 6C, and 6D. Int. J. Med Microbiol. 2014, 304, 1109–1117. [Google Scholar] [CrossRef]
- Liu, Z.; Zheng, H.; Gottschalk, M.; Bai, X.; Lan, R.; Ji, S.; Liu, H.; Xu, J. Development of Multiplex PCR Assays for the Identification of the 33 Serotypes of Streptococcus suis. PLoS ONE 2013, 8, e72070. [Google Scholar] [CrossRef] [Green Version]
- Baker, G.; Smith, J.; Cowan, D. Review and re-analysis of domain-specific 16S primers. J. Microbiol. Methods 2003, 55, 541–555. [Google Scholar] [CrossRef] [Green Version]
- Okwumabua, O.; O’Connor, M.; Shull, E. A polymerase chain reaction (PCR) assay specific forStreptococcus suisbased on the gene encoding the glutamate dehydrogenase. FEMS Microbiol. Lett. 2003, 218, 79–84. [Google Scholar] [CrossRef]
- Tien, L.H.T.; Nishibori, T.; Nishitani, Y.; Nomoto, R.; Osawa, R. Reappraisal of the taxonomy of Streptococcus suis serotypes 20, 22, 26, and 33 based on DNA–DNA homology and sodA and recN phylogenies. Vet. Microbiol. 2013, 162, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Ishida, S.; Tien, L.H.T.; Osawa, R.; Tohya, M.; Nomoto, R.; Kawamura, Y.; Takahashi, T.; Kikuchi, N.; Kikuchi, K.; Sekizaki, T. Development of an appropriate PCR system for the reclassification of Streptococcus suis. J. Microbiol. Methods 2014, 107, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.J.A.; Cernela, N.; Corti, S.; Stephan, R. Draft Genome Sequence of Streptococcus parasuis 4253, the First Available for the Species. Microbiol. Resour. Announc. 2019, 8, e00203-19. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Yang, X.; Qin, J.; Lu, N.; Cheng, G.; Wu, N.; Pan, Y.; Li, J.; Zhu, L.; Wang, X.; et al. Metagenome-wide analysis of antibiotic resistance genes in a large cohort of human gut microbiota. Nat. Commun. 2013, 4, 2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.; Zhang, Q.; Song, Y.; Zhang, Z.; Zhang, A.; Xiao, J.; Jin, M. Characterization of Spectinomycin Resistance in Streptococcus suis Leads to Two Novel Insights into Drug Resistance Formation and Dissemination Mechanism. Antimicrob. Agents Chemother. 2016, 60, 6390–6392. [Google Scholar] [CrossRef] [Green Version]
- Marie, J.; Morvan, H.; Berthelot-Hérault, F.; Sanders, P.; Kempf, I.; Gautier-Bouchardon, A.V.; Jouy, E.; Kobisch, M. Antimicrobial susceptibility of Streptococcus suis isolated from swine in France and from humans in different countries between 1996 and 2000. J. Antimicrob. Chemother. 2002, 50, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-H.; Ha, S.-M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Source | Location | Year | cps Type | Accession No. |
|---|---|---|---|---|---|
| BS26 | Patient | China | 2018 | X | CP069079 |
| BS27 | Patient | China | 2018 | X | JAETXU000000000 |
| 86-5192 | Diseased calf | United States | 1980’ | I | PRJNA171426 |
| 88-1861 | Diseased pig | Canada | 1980’ | II | PRJNA171444 |
| 89-4109-1 | Diseased pig | / | 1980’ | III | PRJNA171433 |
| SUT-7 | Healthy pig | Japan | / | V | DRX016751 |
| SUT-286 | Healthy pig | Japan | / | IV | DRX016752 |
| SUT-319 | Healthy pig | Japan | / | VI | DRX016753 |
| SUT-328 | Healthy pig | Japan | / | VI | DRX016754 |
| SUT-380 | Healthy pig | Japan | / | VI | DRX016755 |
| 10-36905 | Healthy Bos taurus | United States | 2010 | VII | PRJNA590796 |
| 2674 | Healthy pig | China | 2014 | VIII | POIG00000000 |
| 4253 | Healthy cow | Switzerland | 2018 | IX | SHGT00000000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Yi, X.; Liang, P.; Tao, Y.; Wang, Y.; Jin, D.; Luo, B.; Yang, J.; Zheng, H. Investigation of the Genomic and Pathogenic Features of the Potentially Zoonotic Streptococcus parasuis. Pathogens 2021, 10, 834. https://doi.org/10.3390/pathogens10070834
Wang J, Yi X, Liang P, Tao Y, Wang Y, Jin D, Luo B, Yang J, Zheng H. Investigation of the Genomic and Pathogenic Features of the Potentially Zoonotic Streptococcus parasuis. Pathogens. 2021; 10(7):834. https://doi.org/10.3390/pathogens10070834
Chicago/Turabian StyleWang, Jianping, Xueli Yi, Pujun Liang, Yuanmeihui Tao, Yan Wang, Dong Jin, Bin Luo, Jing Yang, and Han Zheng. 2021. "Investigation of the Genomic and Pathogenic Features of the Potentially Zoonotic Streptococcus parasuis" Pathogens 10, no. 7: 834. https://doi.org/10.3390/pathogens10070834
APA StyleWang, J., Yi, X., Liang, P., Tao, Y., Wang, Y., Jin, D., Luo, B., Yang, J., & Zheng, H. (2021). Investigation of the Genomic and Pathogenic Features of the Potentially Zoonotic Streptococcus parasuis. Pathogens, 10(7), 834. https://doi.org/10.3390/pathogens10070834