
Serological Evidence of Multiple Zoonotic Viral Infections among Wild Rodents in Barbados

 ,
,  and
and
Abstract
1. Introduction
2. Results
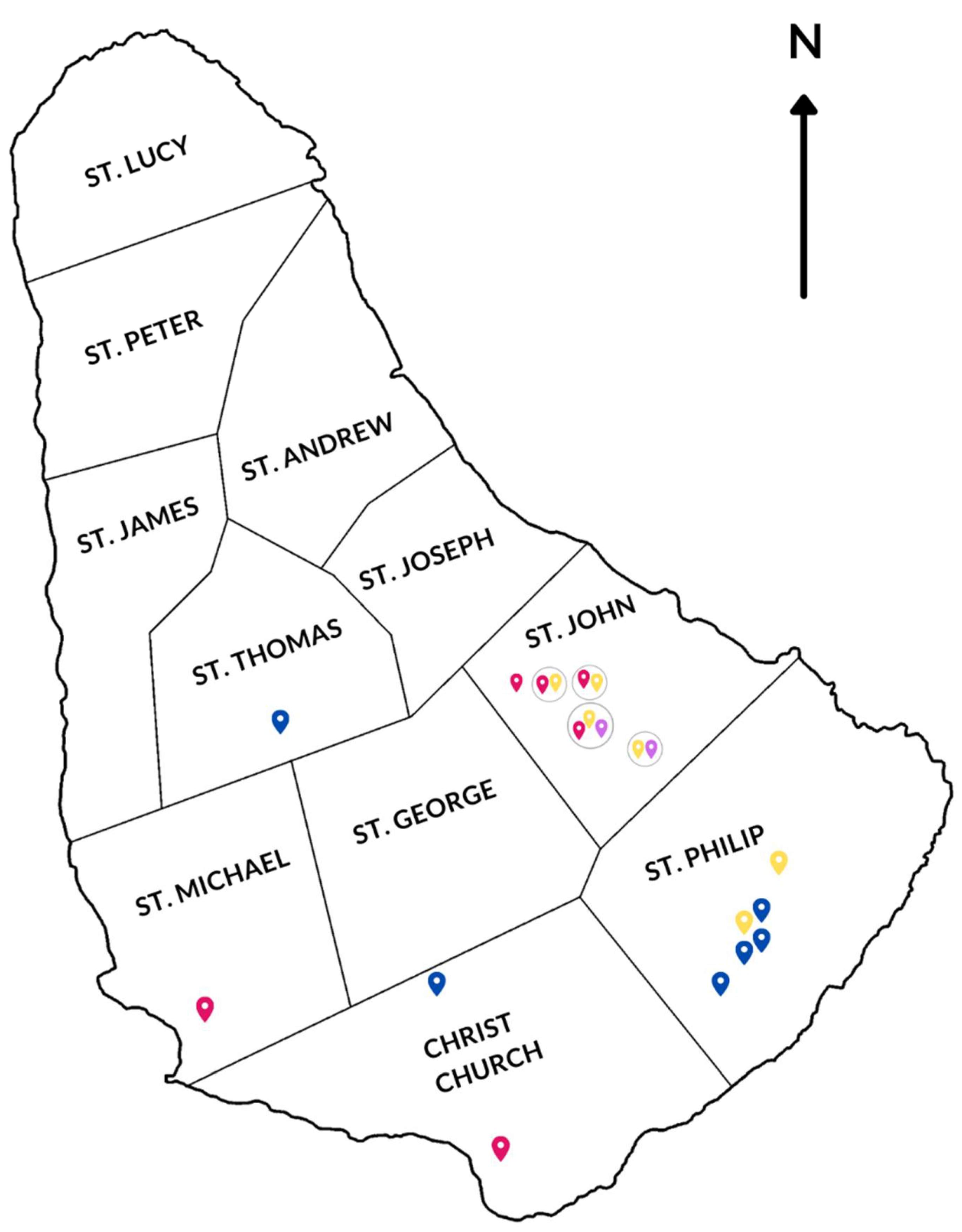
2.1. Wild Rodents Trapping Survey
2.2. Orthohantavirus IFA & RT-PCR Testing of Wild Rodents
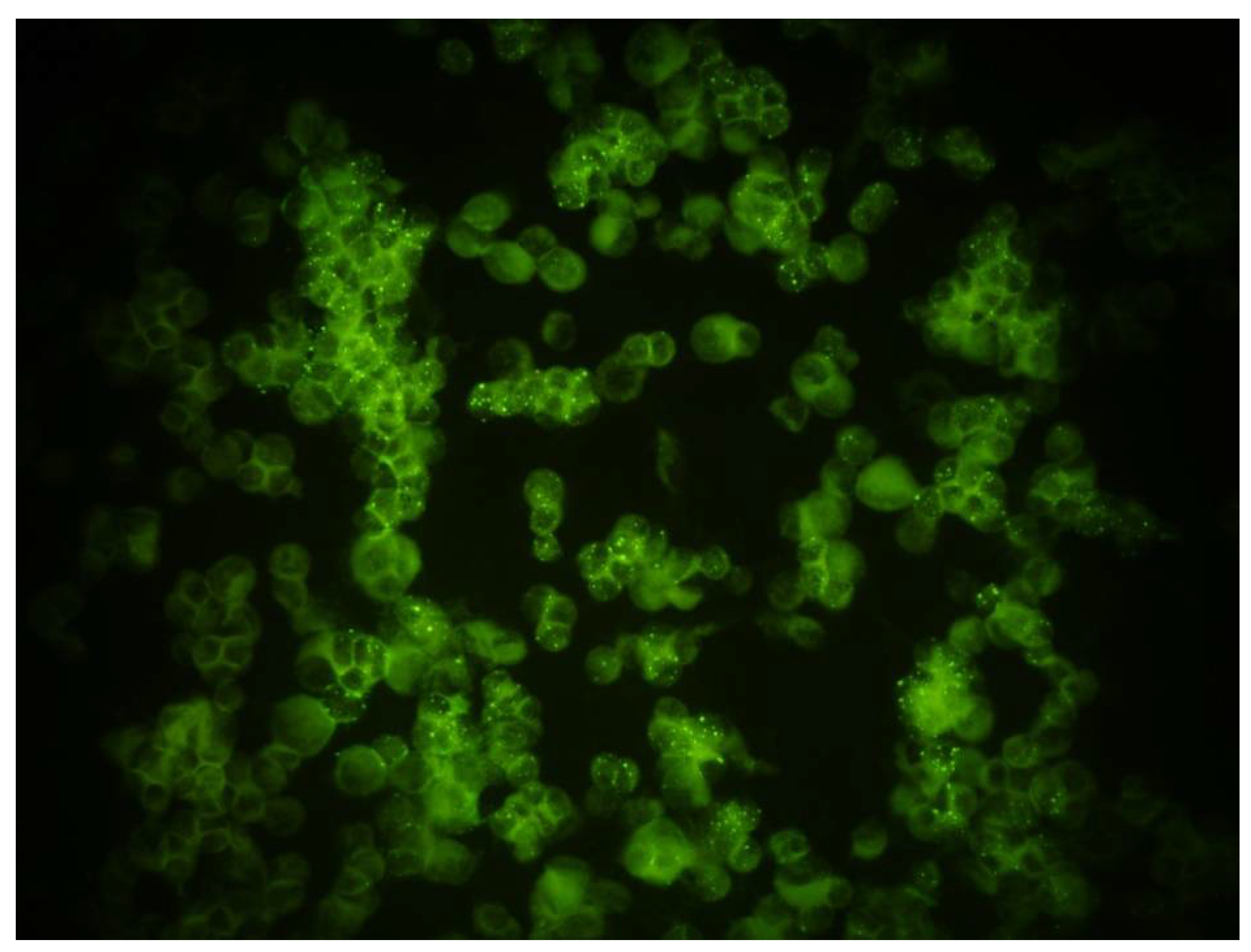
2.3. Orthopoxvirus IFA Testing of Wild Rodents
3. Discussion
3.1. Orthohantavirus Infection among Wild Rodents
3.2. Mammarenavirus IFA Seropositive Rodents
3.3. Orthopoxvirus IFA Seropositive Rodents
3.4. Strengths
3.5. Study Limitations
3.6. Recommendations
4. Materials and Methods
4.1. Rodent Trapping and Sample Collection
4.2. Serological Testing of Rodent Sera
4.3. RNA Extraction and Molecular Testing of Rodent Sera and Tissue
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meerburg, B.G.; Singleton, G.R.; Kijlstra, A. Rodent-borne diseases and their risks for public health. Crit. Rev. Microbiol. 2009, 35, 221–270. [Google Scholar] [CrossRef]
- Meerburg, B.G. Public Health and Rodents: A Game of Cat and Mouse. In Zoonoses—Infections Affecting Humans and Animals; Springer: Berlin/Heidelberg, Germany, 2014; pp. 629–641. [Google Scholar]
- Vapalahti, K.; Paunio, M.; Brummer-Korvenkontio, M.; Vaheri, A.; Vapalahti, O. Puumala virus infections in Finland: Increased occupational risk for farmers. Am. J. Epidemiol. 1999, 149, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Bi, P.; Tong, S.; Donald, K.; Parton, K.; Ni, J. Climatic, reservoir and occupational variables and the transmission of haemorrhagic fever with renal syndrome in China. Int. J. Epidemiol. 2002, 31, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Forbes, K.M.; Sironen, T.; Plyusnin, A. Hantavirus maintenance and transmission in reservoir host populations. Curr. Opin. Virol. 2018, 28, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Joshi, Y.P.; Kim, E.-H.; Cheong, H.-K. The influence of climatic factors on the development of hemorrhagic fever with renal syndrome and leptospirosis during the peak season in Korea: An ecologic study. BMC Infect. Dis. 2017, 17, 406. [Google Scholar] [CrossRef] [PubMed]
- Antezana, J.P.E.; Rodriguez-Villena, O.J.; Arancibia-Alba, A.W.; Alvarado-Arnez, L.E.; Bonilla-Aldana, D.K.; Rodríguez-Morales, A.J. Clinical features of fatal cases of Chapare virus hemorrhagic fever originating from rural La Paz, Bolivia, 2019: A cluster analysis. Travel Med. Infect. Dis. 2020, 36, 101589. [Google Scholar] [CrossRef]
- Knust, B.; Ströher, U.; Edison, L.; Albariño, C.G.; Lovejoy, J.; Armeanu, E.; House, J.; Cory, D.; Horton, C.; Fowler, K.L.; et al. Lymphocytic Choriomeningitis Virus in Employees and Mice at Multipremises Feeder-Rodent Operation, United States, 2012. Emerg. Infect. Dis. 2014, 20, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Peters, C.J. Human Infection with Arenaviruses in the Americas. In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2002; Volume 262, pp. 65–74. [Google Scholar]
- Padula, P.; Figueroa, R.; Navarrete, M.; Pizarro, E.; Cadiz, R.; Bellomo, C.; Jofre, C.; Zaror, L.; Rodriguez, E.; Murúa, R. Transmission Study of Andes Hantavirus Infection in Wild Sigmodontine Rodents. J. Virol. 2004, 78, 11972–11979. [Google Scholar] [CrossRef]
- Nuzum, E.O.; Stephenson, E.H.; LeDuc, J.W.; Rossi, C.A. Aerosol Transmission of Hantaan and Related Viruses to Laboratory Rats. Am. J. Trop. Med. Hyg. 1988, 38, 636–640. [Google Scholar] [CrossRef]
- Skinner, H.H.; Knight, E.H.; Grove, R. Murine lymphocytic choriomeningitis: The history of a natural cross-infection from wild to laboratory mice. Lab. Anim. 1977, 11, 219–222. [Google Scholar] [CrossRef]
- Klein, S.L.; Glass, G.E.; Bird, B.H. Sex differences in immune responses and viral shedding following Seoul virus infection in Norway rats. Am. J. Trop. Med. Hyg. 2001, 65, 57–63. [Google Scholar] [CrossRef]
- Emuyangwa, M.; Martynova, E.V.; Khaiboullina, S.F.; Morzunov, S.P.; Rizvanov, A.A. Hantaviral Proteins: Structure, Functions, and Role in Hantavirus Infection. Front. Microbiol. 2015, 6, 1326. [Google Scholar] [CrossRef]
- Kim, J.-A.; Kim, W.-K.; No, J.S.; Lee, S.-H.; Lee, S.-Y.; Kim, J.H.; Kho, J.H.; Lee, D.; Song, D.H.; Gu, S.H.; et al. Genetic Diversity and Reassortment of Hantaan Virus Tripartite RNA Genomes in Nature, the Republic of Korea. PLoS Negl. Trop. Dis. 2016, 10, e0004650. [Google Scholar] [CrossRef]
- Calisher, C.H.; Briese, T.; Brister, J.R.; Charrel, R.N.; Dürrwald, R.; Ebihara, H.; Fulhorst, C.F.; Gāo, G.F.; Groschup, M.H.; Haddow, A.D.; et al. Strengthening the Interaction of the Virology Community with the International Committee on Taxonomy of Viruses (ICTV) by Linking Virus Names and Their Abbreviations to Virus Species. Syst. Biol. 2019, 68, 828–839. [Google Scholar] [CrossRef]
- Manigold, T.; Vial, P. Human hantavirus infections: Epidemiology, clinical features, pathogenesis and immunology. Swiss Med. Wkly. 2014, 144, w13937. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.-P.; Lin, X.-D.; Wang, W.; Tian, J.-H.; Cong, M.-L.; Zhang, H.-L.; Wang, M.-R.; Zhou, R.-H.; Wang, J.-B.; Li, M.-H.; et al. Phylogeny and Origins of Hantaviruses Harbored by Bats, Insectivores, and Rodents. PLoS Pathog. 2013, 9, e1003159. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Lee, P.W.; Johnson, K.M. Isolation of the Etiologic Agent of Korean Hemorrhagic Fever. J. Infect. Dis. 1978, 137, 298–308. [Google Scholar] [CrossRef]
- Yanagihara, R.; Gu, S.H.; Arai, S.; Kang, H.J.; Song, J.-W. Hantaviruses: Rediscovery and new beginnings. Virus Res. 2014, 187, 6–14. [Google Scholar] [CrossRef]
- Ministry of Health Malaysia. Health Facts 2008; Ministry of Health Malaysia: Putraya, Malaysia, 2009.
- De Thoisy, B.; Matheus, S.; Guitet, S.; Donato, D.; Clément, L.; Lavergne, A.; Brunaux, O.; Guidez, A.; Catzeflis, F.; Lacoste, V.; et al. Maripa Hantavirus in French Guiana: Phylogenetic Position and Predicted Spatial Distribution of Rodent Hosts. Am. J. Trop. Med. Hyg. 2014, 90, 988–992. [Google Scholar] [CrossRef]
- Henderson, W.W.; Monroe, M.C.; Jeor, S.C.S.; Thayerbc, W.P.; Rowe, J.E.; Peters, C.; Nichol, S.T. Naturally Occurring Sin Nombre Virus Genetic Reassortants. Virology 1995, 214, 602–610. [Google Scholar] [CrossRef]
- Bowen, M.D.; Peters, C.J.; Nichol, S.T. Phylogenetic analysis of theArenaviridae: Patterns of virus evolution and evidence for cospeciation between arenaviruses and their rodent hosts. Mol. Phylogenetics Evol. 1997, 8, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus taxonomy: The database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2018, 46, D708–D717. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Sampedro, L.; Perdiguero, B.; Mejías-Pérez, E.; García-Arriaza, J.; Di Pilato, M.; Esteban, M. The Evolution of Poxvirus Vaccines. Viruses 2015, 7, 1726–1803. [Google Scholar] [CrossRef] [PubMed]
- Chantrey, J.; Meyer, H.; Baxby, D.; Begon, M.; Bown, K.J.; Hazel, S.M.; Jones, T.; Montgomery, W.I.; Bennett, M. Cowpox: Reservoir hosts and geographic range. Epidemiol. Infect. 1999, 122, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Vorou, R.M.; Papavassiliou, V.G.; Pierroutsakos, I.N. Cowpox virus infection: An emerging health threat. Curr. Opin. Infect. Dis. 2008, 21, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Suepaul, R.B.; Seetahal, J.F.; Oura, C.; Gyan, L.; Ramoutar, V.V.; Ramkissoon, V.; Sahadeo, N.; Carrington, C.V. Novel poxviral infection in three finch species illegally imported into trinidad, west indies, with implications for native birds. J. Zoo Wildl. Med. 2019, 50, 231–237. [Google Scholar] [CrossRef]
- Douglas, K.; Samuels, T.; Iheozor-Ejiofor, R.; Vapalahti, O.; Sironen, T.; Hilaire, M.G.-S. Serological Evidence of Human Orthohantavirus Infections in Barbados, 2008 to 2016. Pathogens 2021, 10, 571. [Google Scholar] [CrossRef]
- Groen, J.; Koraka, P.; Edwards, C.; Branch, S.; Douglas, K.; Osterhaus, A.; Levett, P. Serological Evidence of Hantavirus in Humans and Rodents in Barbados. J. Infect. 2002, 45, 109–110. [Google Scholar] [CrossRef]
- Clement, J.; LeDuc, J.W.; Lloyd, G.; Reynes, J.-M.; McElhinney, L.; Van Ranst, M.; Lee, H.-W. Wild Rats, Laboratory Rats, Pet Rats: Global Seoul Hantavirus Disease Revisited. Viruses 2019, 11, 652. [Google Scholar] [CrossRef]
- Keenan, L.M.; Chikweto, A.; Sharma, R.N.; Stone, D.M. Seroprevalence of hantavirus in Rattus norvegicus in Grenada, West Indies. West Indian Vet. J. 2008, 8, 67–71. [Google Scholar]
- Raboni, S.M.; Delfraro, A.; De Borba, L.; Teixeira, B.R.; Stella, V.; De Araujo, M.R.; Carstensen, S.; Rubio, G.; Maron, A.; Lemos, E.R.S.; et al. Hantavirus Infection Prevalence in Wild Rodents and Human Anti-Hantavirus Serological Profiles from Different Geographic Areas of South Brazil. Am. J. Trop. Med. Hyg. 2012, 87, 371–378. [Google Scholar] [CrossRef]
- Armién, A.G.; Lee, S.-J.; Avila, M.; Armién, B.; Gonzalez, P.; Gracia, F.; Pascale, J.M.; Mendoza, Y.; Yates, T.L.; Koster, F.; et al. Hantavirus infection and habitat associations among rodent populations in agroecosystems of Panama: Implications for human disease risk. Am. J. Trop. Med. Hyg. 2009, 81, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Bravo, J.; Armién, B.; Suzán, G.; Armién, A.; Ruedas, L.A.; Avila, M.; Zaldívar, Y.; Pascale, J.M.; Gracia, F.; Yates, T.L. Serosurvey of Wild Rodents for Hantaviruses in Panama, 2000–2002. J. Wildl. Dis. 2004, 40, 103–109. [Google Scholar] [CrossRef]
- Eastwood, G.; Camp, J.V.; Chu, Y.K.; Sawyer, A.M.; Owen, R.D.; Cao, X.; Taylor, M.K.; Valdivieso-Torres, L.; Sage, R.D.; Yu, A.; et al. Habitat, species richness and hantaviruses of sigmodontine rodents within the Interior Atlantic Forest, Paraguay. PLoS ONE 2018, 13, e0201307. [Google Scholar] [CrossRef] [PubMed]
- Pacsa, A.; Elbishbishi, E.; Chaturvedi, U.; Chu, K.; Mustafa, A. Hantavirus-specific antibodies in rodents and humans living in Kuwait. FEMS Immunol. Med Microbiol. 2002, 33, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.-F.; Zhao, W.-J.; Cao, W.-C.; Wu, X.-M.; Tang, F.; Wang, R.-M.; Zhang, P.-H.; Zhang, W.-Y.; Dun, Z.; Zuo, S.-Q.; et al. Prevalence and genetic diversities of hantaviruses in rodents in Beijing, China. Am. J. Trop. Med. Hyg. 2008, 78, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Heyman, P.; Baert, K.; Plyusnina, A.; Cochez, C.; Lundkvist, A.; Van Esbroeck, M.; Goossens, E.; Vandenvelde, C.; Plyusnin, A.; Stuyck, J. Serological and genetic evidence for the presence of Seoul hantavirus in Rattus norvegicus in Flanders, Belgium. Scand. J. Infect. Dis. 2009, 41, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Glass, G.E.; Childs, J.E.; Korch, G.W.; LeDuc, J.W. Association of intraspecific wounding with hantaviral infection in wild rats (Rattus norvegicus). Epidemiol. Infect. 1988, 101, 459–472. [Google Scholar] [CrossRef]
- Rivas, Y.J.; Moros, Z.; Morón, D.; Uzcátegui, M.G.; Duran, Z.; Pujol, F.H.; Liprandi, F.; Ludert, J.E. The seroprevalences of anti-hantavirus IgG antibodies among selected Venezuelan populations. Ann. Trop. Med. Parasitol. 2003, 97, 61–67. [Google Scholar] [CrossRef]
- Fulhorst, C.F.; Cajimat, M.N.; Utrera, A.; Milazzo, M.L.; Duno, G.M. Maporal virus, a hantavirus associated with the fulvous pygmy rice rat (Oligoryzomys fulvescens) in western Venezuela. Virus Res. 2004, 104, 139–144. [Google Scholar] [CrossRef]
- Vargas, J.A.; Dickson, S.; Rifakis, P.M.; Meo, F.; Gonçalves, N.; Cardoso, E.; Hernández, F.; Alurralde, C.; Rodriguez-Morales, A.J.; Franco-Paredes, C. Rodent reservoirs of zoonotic viral hemorrhagic fevers in Venezuela. Acta Cient. Estud 2006, 4, 80–87. [Google Scholar]
- Milazzo, M.L.; Duno, G.; Utrera, A.; Richter, M.H.; Duno, F.; De Manzione, N.; Fulhorst, C.F. Natural Host Relationships of Hantaviruses Native to Western Venezuela. Vector-Borne Zoonotic Dis. 2010, 10, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Matheus, S.; Al, S.M.E.; Mayence, C.; Bremand, L.; Houcke, S.; Rousset, D.; Lacoste, V.; De Thoisy, B.; Hommel, D.; Lavergne, A. Hantavirus Pulmonary Syndrome Caused by Maripa Virus in French Guiana, 2008–2016. Emerg. Infect. Dis. 2017, 23, 1722–1725. [Google Scholar] [CrossRef] [PubMed]
- Tersago, K.; Verhagen, R.; Servais, A.; Heyman, P.; Ducoffre, G.; Leirs, H. Hantavirus disease (nephropathia epidemica) in Belgium: Effects of tree seed production and climate. Epidemiol. Infect. 2008, 137, 250–256. [Google Scholar] [CrossRef]
- Xiao, H.; Tian, H.-Y.; Cazelles, B.; Li, X.-J.; Tong, S.-L.; Gao, L.-D.; Qin, J.-X.; Lin, X.-L.; Liu, H.-N.; Zhang, X.-X. Atmospheric Moisture Variability and Transmission of Hemorrhagic Fever with Renal Syndrome in Changsha City, Mainland China, 1991–2010. PLoS Negl. Trop. Dis. 2013, 7, e2260. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.R.; Lee, Y.T.; Park, C.H. A new natural reservoir of hantavirus: isolation of hantaviruses from lung tissues of bats. Arch. Virol. 1994, 134, 85–95. [Google Scholar] [CrossRef]
- Sabino-Santos Jr, G.; Maia, F.G.M.; Vieira, T.M.; de Lara Muylaert, R.; Lima, S.M.; Gonçalves, C.B.; Barroso, P.D.; Melo, M.N.; Jonsson, C.B.; Goodin, D. Evidence of hantavirus infection among bats in Brazil. Am. J. Trop. Med. Hyg. 2015, 93, 404–406. [Google Scholar] [CrossRef]
- Weiss, S.; Witkowski, P.T.; Auste, B.; Nowak, K.; Weber, N.; Fahr, J.; Mombouli, J.-V.; Wolfe, N.D.; Drexler, J.F.; Drosten, C. Hantavirus in bat, sierra leone. Emerg. Infect. Dis. 2012, 18, 159. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wu, J.; He, B.; Qin, S.; Xia, L.; Qin, M.; Li, N.; Tu, C. Novel hantavirus identified in black-bearded tomb bats, China. Infect. Genet. Evol. 2015, 31, 158–160. [Google Scholar] [CrossRef]
- de Araujo, J.; Thomazelli, L.M.; Henriques, D.A.; Lautenschalager, D.; Ometto, T.; Dutra, L.M.; Aires, C.C.; Favorito, S.; Durigon, E.L. Detection of hantavirus in bats from remaining rain forest in São Paulo, Brazil. BMC Res. Notes 2012, 5, 690. [Google Scholar] [CrossRef]
- Lavergne, A.; de Thoisy, B.; Tirera, S.; Donato, D.; Bouchier, C.; Catzeflis, F.; Lacoste, V. Identification of lymphocytic choriomeningitis mammarenavirus in house mouse (Mus musculus, Rodentia) in French Guiana. Infect. Genet. Evol. 2016, 37, 225–230. [Google Scholar] [CrossRef]
- Fernandes, J.; Guterres, A.; de Oliveira, R.C.; Chamberlain, J.; Lewandowski, K.; Teixeira, B.R.; Coelho, T.A.; Crisóstomo, C.F.; Bonvicino, C.R.; D’Andrea, P.S. Xapuri virus, a novel mammarenavirus: Natural reassortment and increased diversity between New World viruses. Emerg. Microbes Infect. 2018, 7, 1–10. [Google Scholar] [CrossRef]
- Castellar, A.; Guevara, M.; Rodas, J.D.; Londoño, A.F.; Arroyave, E.; Díaz, F.J.; Levis, S.; Blanco, P.J. First evidence of Lymphocytic Choriomeningitis virus (Arenaviruses) infection in Mus musculus rodents captured in the urban area of the municipality of Sincelejo-Sucre, Colombia. Biomédica 2016, 37, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Malmlov, A.; Seetahal, J.; Carrington, C.; Ramkisson, V.; Foster, J.; Miazgowicz, K.L.; Quackenbush, S.; Rovnak, J.; Negrete, O.; Munster, V.; et al. Serological evidence of arenavirus circulation among fruit bats in Trinidad. PLoS ONE 2017, 12, e0185308. [Google Scholar] [CrossRef] [PubMed]
- Talley, P. Lymphocytic Choriomeningitis Virus Meningoencephalitis from a Household Rodent Infestation—Minnesota, 2015; MMWR Morbidity and Mortality Weekly Report; CDC: Atlanta, GA, USA, 2016; Volume 65.
- Centers for Disease Control and Prevention. Notes from the Field: Lymphocytic Choriomeningitis Virus Infections in Employees of a Rodent Breeding Facility--Indiana, May–June 2012; MMWR. Morbidity and Mortality Weekly Report; CDC: Atlanta, GA, USA, 2012; Volume 61, pp. 622–623.
- Bisordi, I.; Levis, S.; Maeda, A.Y.; Suzuki, A.; Nagasse-Sugahara, T.K.; de Souza, R.P.; Pereira, L.E.; Garcia, J.B.; Cerroni, M.D.P.; Silva, F.D.A.E.; et al. Pinhal Virus, a New Arenavirus Isolated from Calomys tener in Brazil. Vector-Borne Zoonotic Dis. 2015, 15, 694–700. [Google Scholar] [CrossRef]
- Milazzo, M.L.; Cajimat, M.N.B.; Mauldin, M.R.; Bennett, S.G.; Hess, B.D.; Rood, M.P.; Conlan, C.A.; Nguyen, K.; Wekesa, J.W.; Ramos, R.D.; et al. Epizootiology of Tacaribe Serocomplex Viruses (Arenaviridae) Associated with Neotomine Rodents (Cricetidae, Neotominae) in Southern California. Vector-Borne Zoonotic Dis. 2015, 15, 156–166. [Google Scholar] [CrossRef]
- Lavergne, A.; De Thoisy, B.; Donato, D.; Guidez, A.; Matheus, S.; Catzeflis, F.; Lacoste, V. Patawa Virus, a New Arenavirus Hosted by Forest Rodents in French Guiana. EcoHealth 2014, 12, 339–346. [Google Scholar] [CrossRef]
- Milazzo, M.L.; Iralu, J.; Fulhorst, C.F.; Koster, F. Antibody against Arenaviruses in Humans, Southwestern United States. Emerg. Infect. Dis. 2014, 20, 1592–1593. [Google Scholar] [CrossRef]
- Milazzo, M.L.; Campbell, G.L.; Fulhorst, C.F. Novel Arenavirus Infection in Humans, United States. Emerg. Infect. Dis. 2011, 17, 1417–1420. [Google Scholar] [CrossRef] [PubMed]
- Milazzo, M.L.; Amman, B.R.; Cajimat, M.N.; Méndez-Harclerode, F.M.; Suchecki, J.R.; Hanson, J.D.; Haynie, M.L.; Baxter, B.D.; Milazzo, C., Jr.; Carroll, S.A. Ecology of Catarina virus (family Arenaviridae) in southern Texas, 2001–2004. Vector-Borne Zoonotic Dis. 2013, 13, 50–59. [Google Scholar] [CrossRef]
- Laakkonen, J.; Kallio-Kokko, H.; Öktem, M.A.; Blasdell, K.; Plyusnina, A.; Niemimaa, J.; Karatas, A.; Vaheri, A.; Henttonen, H. Serological Survey for Viral Pathogens in Turkish Rodents. J. Wildl. Dis. 2006, 42, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Kallio-Kokko, H.; Laakkonen, J.; Rizzoli, A.; Tagliapietra, V.; Cattadori, I.; Perkins, S.E.; Hudson, P.J.; Cristofolini, A.; Versini, W.; Vapalahti, O.; et al. Hantavirus and arenavirus antibody prevalence in rodents and humans in Trentino, Northern Italy. Epidemiol. Infect. 2005, 134, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Blasdell, K.R.; Becker, S.D.; Hurst, J.; Begon, M.; Bennett, M. Host Range and Genetic Diversity of Arenaviruses in Rodents, United Kingdom. Emerg. Infect. Dis. 2008, 14, 1455–1458. [Google Scholar] [CrossRef] [PubMed]
- Kosoy, M.Y.; Maupin, G.O.; Rollin, P.E.; Fulhorst, C.F.; Mills, J.N.; Childs, J.E.; Ksiazek, T.G.; Peters, C.J.; Elliott, L.H. Prevalence of Antibodies to Arenaviruses in Rodents from the Southern and Western United States: Evidence for an Arenavirus Associated with the Genus Neotoma. Am. J. Trop. Med. Hyg. 1996, 54, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Amanat, F.; Duehr, J.; Oestereich, L.; Hastie, K.M.; Saphire, E.O.; Krammer, F. Antibodies to the Glycoprotein GP2 Subunit Cross-React between Old and New World Arenaviruses. mSphere 2018, 3, e00189-18. [Google Scholar] [CrossRef] [PubMed]
- Buchmeier, M.J.; Lewicki, H.A.; Tomori, O.; Oldstone, M.B. Monoclonal antibodies to lymphocytic choriomeningitis and pichinde viruses: Generation, characterization, and cross-reactivity with other arenaviruses. Virology 1981, 113, 73–85. [Google Scholar] [CrossRef]
- Downs, W.G.; Anderson, C.R.; Spence, L.; Aitken, T.H.G.; Greenhall, A.H. Tacaribe Virus, a New Agent Isolated from Artibeus Bats and Mosquitoes in Trinidad, West Indies. Am. J. Trop. Med. Hyg. 1963, 12, 640–646. [Google Scholar] [CrossRef]
- Cogswell-Hawkinson, A.; Bowen, R.; James, S.; Gardiner, D.; Calisher, C.H.; Adams, R.; Schountz, T. Tacaribe Virus Causes Fatal Infection of An Ostensible Reservoir Host, the Jamaican Fruit Bat. J. Virol. 2012, 86, 5791–5799. [Google Scholar] [CrossRef]
- Sayler, K.A.; Barbet, A.F.; Chamberlain, C.; Clapp, W.L.; Alleman, R.; Loeb, J.C.; Lednicky, J.A. Isolation of Tacaribe Virus, a Caribbean Arenavirus, from Host-Seeking Amblyomma americanum Ticks in Florida. PLoS ONE 2014, 9, e115769. [Google Scholar] [CrossRef]
- Kinnunen, P.M.; Henttonen, H.; Hoffmann, B.; Kallio, E.R.; Korthase, C.; Laakkonen, J.; Niemimaa, J.; Palva, A.; Schlegel, M.; Ali, H.S.; et al. Orthopox Virus Infections in Eurasian Wild Rodents. Vector-Borne Zoonotic Dis. 2011, 11, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Oldal, M.; Sironen, T.; Henttonen, H.; Vapalahti, O.; Madai, M.; Horváth, G.; Dallos, B.; Kutas, A.; Földes, F.; Kemenesi, G.; et al. Serologic Survey of Orthopoxvirus Infection Among Rodents in Hungary. Vector-Borne Zoonotic Dis. 2015, 15, 317–322. [Google Scholar] [CrossRef]
- Salzer, J.S.; Carroll, D.S.; Rwego, I.B.; Li, Y.; Falendysz, E.A.; Shisler, J.L.; Karem, K.L.; Damon, I.K.; Gillespie, T.R. Serologic evidence for circulating orthopoxviruses in peridomestic rodents from rural uganda. J. Wildl. Dis. 2013, 49, 125–131. [Google Scholar] [CrossRef][Green Version]
- Vaughan, A.; Aarons, E.; Astbury, J.; Brooks, T.; Chand, M.; Flegg, P.; Hardman, A.; Harper, N.; Jarvis, R.; Mawdsley, S.; et al. Human-to-Human Transmission of Monkeypox Virus, United Kingdom, October 2018. Emerg. Infect. Dis. 2020, 26, 782–785. [Google Scholar] [CrossRef] [PubMed]
- Parker, S.; Nuara, A.; Buller, R.M.L.; A Schultz, D. Human monkeypox: An emerging zoonotic disease. Future Microbiol. 2007, 2, 17–34. [Google Scholar] [CrossRef]
- Marennikova, S.S.; Shelukhina, E.M. White rats as source of pox infection in carnivora of the family Felidae. Acta Virol. 1976, 20, 442. [Google Scholar] [PubMed]
- Wolfs, T.F.; Wagenaar, J.A.; Niesters, H.G.; Osterhaus, A. Rat-to-Human Transmission of Cowpox Infection. Emerg. Infect. Dis. 2002, 8, 1495–1496. [Google Scholar] [CrossRef]
- Baxby, D.; Bennett, M. Cowpox: A re-evaluation of the risks of human cowpox based on new epidemiological information. Viral Zoonoses Food Anim. Orig. 1997, 13, 1–12. [Google Scholar] [CrossRef]
- Baxby, D.; Bennett, M.; Getty, B. Human cowpox 1969–93: A review based on 54 cases. Br. J. Dermatol. 1994, 131, 598–607. [Google Scholar] [CrossRef] [PubMed]
- Campe, H.; Zimmermann, P.; Glos, K.; Bayer, M.; Bergemann, H.; Dreweck, C.; Graf, P.; Weber, B.K.; Meyer, H.; Büttner, M.; et al. Cowpox Virus Transmission from Pet Rats to Humans, Germany. Emerg. Infect. Dis. 2009, 15, 777–780. [Google Scholar] [CrossRef]
- Favier, A.-L.; Flusin, O.; Lepreux, S.; Fleury, H.; Labrèze, C.; Georges, A.; Crance, J.-M.; Boralevi, F. Necrotic Ulcerated Lesion in a Young Boy Caused by Cowpox Virus Infection. Case Rep. Dermatol. 2011, 3, 186–194. [Google Scholar] [CrossRef]
- Becker, C.; Kurth, A.; Hessler, F.; Kramp, H.; Gokel, M.; Hoffmann, R.; Kuczka, A.; Nitsche, A. Cowpox virus infection in pet rat owners: Not always immediately recognized. Dtsch. Ärzteblatt Int. 2009, 106, 329. [Google Scholar] [CrossRef]
- Kurth, A.; Straube, M.; Kuczka, A.; Dunsche, A.J.; Meyer, H.; Nitsche, A. Cowpox Virus Outbreak in Banded Mongooses (Mungos mungo) and Jaguarundis (Herpailurus yagouaroundi) with a Time-Delayed Infection to Humans. PLoS ONE 2009, 4, e6883. [Google Scholar] [CrossRef]
- Eis-Hubinger, A.; Gerritzen, A.; Schneweis, K.; Pfeiff, B.; Pullmann, H.; Mayr, A.; Czerny, C.-P. Fatal cowpox-like virus infection transmitted by cat. Lancet 1990, 336, 880. [Google Scholar] [CrossRef]
- Pelkonen, P.M.; Tarvainen, K.; Hynninen, A.; Al, P.M.P.E.; Henttonen, H.; Palva, A.; Vaheri, A.; Vapalahti, O. Cowpox with Severe Generalized Eruption, Finland. Emerg. Infect. Dis. 2003, 9, 1458–1461. [Google Scholar] [CrossRef]
- Czerny, C.P.; Eis-Hübinger, A.; Mayr, A.; Schneweis, K.; Pfeiff, B. Animal poxviruses transmitted from cat to man: Current event with lethal end. J. Vet. Med. Ser. B 1991, 38, 421–431. [Google Scholar] [CrossRef]
- Sumibcay, L.; Kadjo, B.; Gu, S.H.; Kang, H.J.; Lim, B.K.; A Cook, J.; Song, J.-W.; Yanagihara, R. Divergent lineage of a novel hantavirus in the banana pipistrelle (Neoromicia nanus) in Côte d’Ivoire. Virol. J. 2012, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- Klempa, B.; Fichet-Calvet, E.; Lecompte, E.; Auste, B.; Aniskin, V.; Meisel, H.; Barrière, P.; Koivogui, L.; Ter Meulen, J.; Krüger, D.H. Novel Hantavirus Sequences in Shrew, Guinea. Emerg. Infect. Dis. 2007, 13, 520–522. [Google Scholar] [CrossRef] [PubMed]
- Vieth, S.; Drosten, C.; Lenz, O.; Vincent, M.; Omilabu, S.; Hass, M.; Becker-Ziaja, B.; Ter Meulen, J.; Nichol, S.T.; Schmitz, H.; et al. RT-PCR assay for detection of Lassa virus and related Old World arenaviruses targeting the L gene. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 1253–1264. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gender and Reproductive Status | |||||||
|---|---|---|---|---|---|---|---|
| Male | Female | ||||||
| No. of Trapped Rodents | Rodent Species | N | SC | Total | PR | NP | Total |
| 150 *,+ | Mus musculus | 35 | 46 | 81 | 12 | 52 | 64 |
| 8 * | Rattus norvegicus | 1 | 0 | 1 | - | 6 | 6 |
| 2 | Rattus rattus | - | 2 | 2 | - | - | 0 |
| 160 | 36 | 48 | 84 | 12 | 58 | 70 | |
| Rodent Species | No. of Trapped Rodents | Trapping Location | IFA Testing | RT-PCR | |||
|---|---|---|---|---|---|---|---|
| Orthohantavirus (PUUV) | Mammarenavirus (LCMV) | Orthopoxvirus (CPXV) | Orthohantavirus | Mammarenavirus | |||
| Rattus rattus | 2 | St. John | 0/2 | 0/2 | 1/2 | - | - |
| Rattus norvegicus | 8 | St. Philip | 0/8 | 0/8 | 1/8 | - | - |
| Mus musculus | 5 | St. Michael | 1/5 | 0/5 | 0/5 | 0/1 | - |
| 3 | St. Thomas | 0/3 | 0/3 | 0/3 | - | - | |
| 90 | St. John | 4/90 | 4/90 | 5/90 | 0/4 | 0/4 | |
| 35 | St. Philip | 0/35 | 0/35 | 5/35 | - | - | |
| 7 | Christ Church | 1/7 | 0/7 | 0/7 | 0/1 | - | |
| 160 | 6/160 (4.0%) | 4/160 (2.5%) | 12/160 (7.5%) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Douglas, K.O.; Cayol, C.; Forbes, K.M.; Samuels, T.A.; Vapalahti, O.; Sironen, T.; Gittens-St. Hilaire, M. Serological Evidence of Multiple Zoonotic Viral Infections among Wild Rodents in Barbados. Pathogens 2021, 10, 663. https://doi.org/10.3390/pathogens10060663
Douglas KO, Cayol C, Forbes KM, Samuels TA, Vapalahti O, Sironen T, Gittens-St. Hilaire M. Serological Evidence of Multiple Zoonotic Viral Infections among Wild Rodents in Barbados. Pathogens. 2021; 10(6):663. https://doi.org/10.3390/pathogens10060663
Chicago/Turabian StyleDouglas, Kirk Osmond, Claire Cayol, Kristian Michael Forbes, Thelma Alafia Samuels, Olli Vapalahti, Tarja Sironen, and Marquita Gittens-St. Hilaire. 2021. "Serological Evidence of Multiple Zoonotic Viral Infections among Wild Rodents in Barbados" Pathogens 10, no. 6: 663. https://doi.org/10.3390/pathogens10060663
APA StyleDouglas, K. O., Cayol, C., Forbes, K. M., Samuels, T. A., Vapalahti, O., Sironen, T., & Gittens-St. Hilaire, M. (2021). Serological Evidence of Multiple Zoonotic Viral Infections among Wild Rodents in Barbados. Pathogens, 10(6), 663. https://doi.org/10.3390/pathogens10060663