
Scrub Typhus and Molecular Characterization of Orientia tsutsugamushi from Central Nepal

 ,
, {kind=link}
{kind=link}
Abstract
1. Introduction
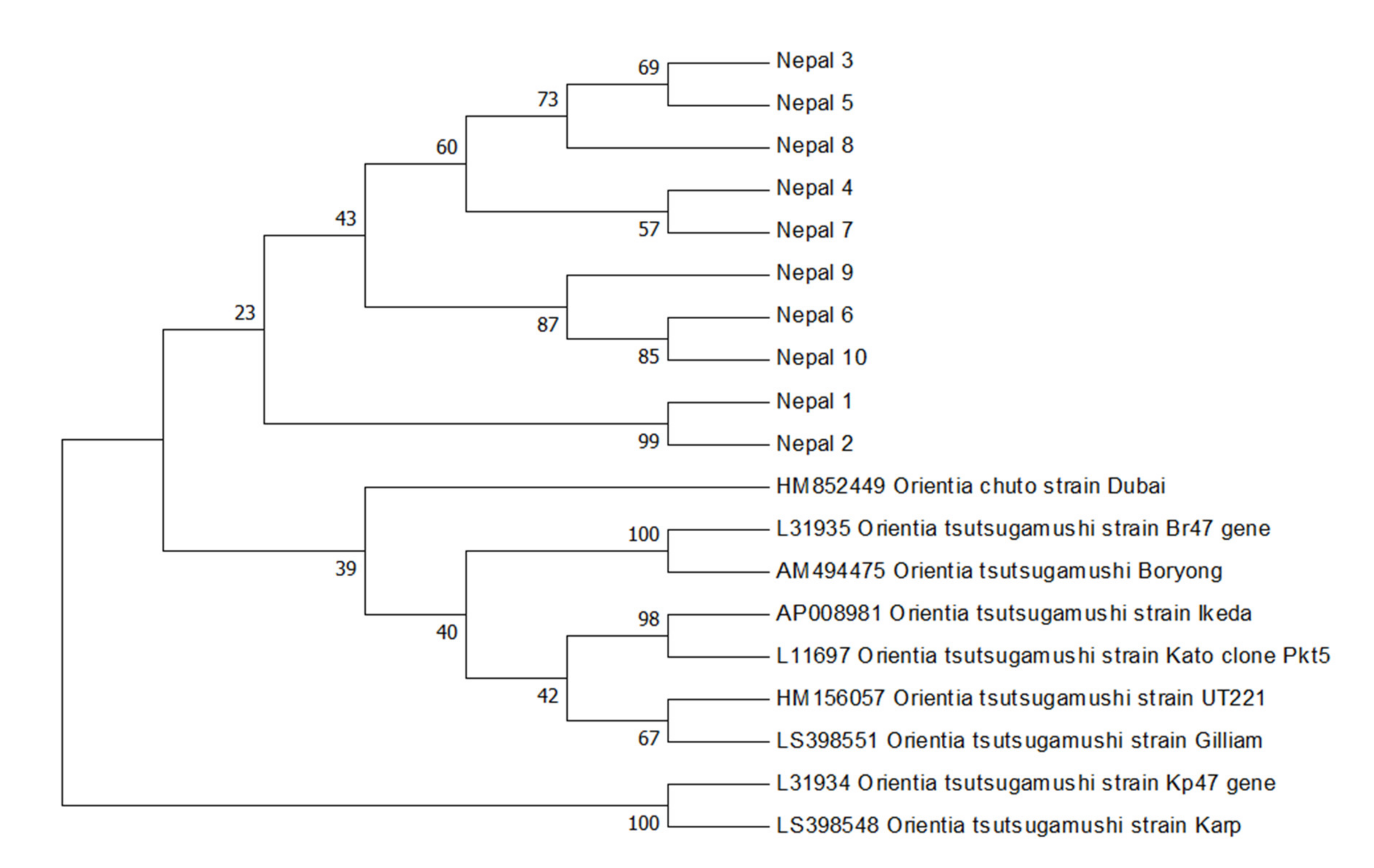
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Rickettsia PCR Screening and Sequencing
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luce-Fedrow, A.; Lehman, M.L.; Kelly, D.J.; Mullins, K.; Maina, A.N.; Stewart, R.L.; Ge, H.; John, H.S.; Jiang, J.; Richards, A.L. A review of scrub typhus (Orientia tsutsugamushi and related organisms): Then, now, and tomorrow. Trop. Med. Infect. Dis. 2018, 3, 8. [Google Scholar] [CrossRef]
- La Scola, B.; Raoult, D. Laboratory diagnosis of rickettsioses: Current approaches to diagnosis of old and new rickettsial diseases. J. Clin. Microbiol. 1997, 35, 2715–2727. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Chan, T.C.; Temenak, J.J.; Dasch, G.A.; Ching, W.M.; Richards, A.L. Development of a quantitative real-time polymerase chain reaction assay specific for Orientia tsutsugamushi. Am. J. Trop. Med. Hyg. 2004, 70, 351–356. [Google Scholar] [CrossRef]
- Ohashi, N.; Nashimoto, H.; Ikeda, H.; Tamura, A. Diversity of immunodominant 56-kDa type-specific antigen (TSA) of Rickettsia tsutsugamushi. Sequence and comparative analyses of the genes encoding TSA homologues from four antigenic variants. J. Biol. Chem. 1992, 267, 12728–12735. [Google Scholar] [CrossRef]
- Ni, Y.S.; Chan, T.C.; Chao, C.C.; Richards, A.L.; Dasch, G.A.; Ching, W.M. Protection against scrub typhus by a plasmid vaccine encoding the 56-KD outer membrane protein antigen gene. Am. J. Trop. Med. Hyg. 2005, 73, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Paris, D.H.; Blacksell, S.D.; Aukkanit, N.; Newton, P.N.; Phetsouvanh, R.; Izzard, L.; Stenos, J.; Graves, S.R.; Day, N.P.; et al. Diversity of the 47-kD HtrA nucleic acid and translated amino acid sequences from 17 recent human isolates of Orientia. Vector Borne Zoonotic Dis. 2013, 13, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.J.; Fuerst, P.A.; Ching, W.M.; Richards, A.L. Scrub typhus: The geographic distribution of phenotypic and genotypic variants of Orientia tsutsugamushi. Clin. Infect. Dis. 2009, 48, S203–S230. [Google Scholar] [CrossRef]
- Fournier, P.E.; Siritantikorn, S.; Rolain, J.M.; Suputtamongkol, Y.; Hoontrakul, S.; Charoenwat, S.; Losuwanaluk, K.; Parola, P.; Raoult, D. Detection of new genotypes of Orientia tsutsugamushi infecting humans in Thailand. Clin. Microbiol. Infect. 2008, 14, 168–173. [Google Scholar] [CrossRef]
- Blacksell, S.D.; Luksameetanasan, R.; Kalambaheti, T.; Aukkanit, N.; Paris, D.H.; McGready, R.; Nosten, F.; Peacock, S.J.; Day, N.P. Genetic typing of the 56-kDa type-specific antigen gene of contemporary Orientia tsutsugamushi isolates causing human scrub typhus at two sites in north-eastern and western Thailand. FEMS Immunol. Med. Microbiol. 2008, 52, 335–342. [Google Scholar] [CrossRef]
- Upadhyaya, B.P.; Shakya, G.; Adhikari, S.; Rijal, N.; Acharya, J.; Maharjan, L.; Marasini, B.R. Scrub typhus: An emergent neglected tropical disease in Nepal. J. Nepal. Health Res. Counc. 2016, 14, 122–127. [Google Scholar]
- Gautam, R.; Parajuli, K.; Sherchand, J.B. Epidemiology, risk factors and seasonal variation of scrub typhus fever in Central Nepal. Trop. Med. Infect. Dis. 2019, 4, 27. [Google Scholar] [CrossRef]
- Karki, K. Descriptive Epidemiology of Scrub Typhus in Nepal; Nepal Health Research Council: Kathmandu, Nepal, 2017.
- Acharya, K.P.; Adhikari, N.; Tariq, M. Scrub typhys: A serious public health issue in Nepal. Clin. Epidemiol. Glob. Health 2020, 8, 815–817. [Google Scholar] [CrossRef]
- Paris, D.H.; Blacksell, S.D.; Nawtaisong, P.; Jenjaroen, K.; Teeraratkul, A.; Chierakul, W.; Wuthiekanun, V.; Kantipong, P.; Day, N.P. Diagnostic accuracy of a loop-mediated isothermal PCR assay for detection of Orientia tsutsugamushi during acute scrub typhus infection. PLoS Negl. Trop. Dis. 2011, 5, e1307. [Google Scholar] [CrossRef]
- Long, J.; Wei, Y.; Tao, X.; He, P.; Xu, J.; Wu, X.; Zhu, W.; Chen, K.; Yang, Z. Representative genotyping, recombination and evolutionary dynamics analysis of TSA56 gene segment of Orientia tsutsugamushi. Front. Cell. Infect. Microbiol. 2020, 10, 383. [Google Scholar] [CrossRef]
- Kelly, D.J.; Fuerst, P.A.; Richards, A.L. Origins, importance and genetic stability of the prototype strains Gilliam, Karp and Kato of Orientia tsutsugamushi. Trop. Med. Infect. Dis. 2019, 4. [Google Scholar] [CrossRef]
- De, W.; Jing, K.; Huan, Z.; Qiong, Z.H.; Monagin, C.; Min, Z.J.; Ping, H.; Wen, K.C.; Yan, L.J. Scrub typhus, a disease with increasing threat in Guangdong, China. PLoS ONE 2015, 10, e0113968. [Google Scholar] [CrossRef]
- Ruang-Areerate, T.; Jeamwattanalert, P.; Rodkvamtook, W.; Richards, A.L.; Sunyakumthorn, P.; Gaywee, J. Genotype diversity and distribution of Orientia tsutsugamushi causing scrub typhus in Thailand. J. Clin. Microbiol. 2011, 49, 2584–2589. [Google Scholar] [CrossRef]
- Takhampunya, R.; Tippayachai, B.; Promsathaporn, S.; Leepitakrat, S.; Monkanna, T.; Schuster, A.L.; Melendrez, M.C.; Paris, D.H.; Richards, A.L.; Richardson, J.H. Characterization based on the 56-Kda type-specific antigen gene of Orientia tsutsugamushi genotypes isolated from Leptotrombidium mites and the rodent host post-infection. Am. J. Trop. Med. Hyg. 2014, 90, 139–146. [Google Scholar] [CrossRef]
- Kim, G.; Ha, N.Y.; Min, C.K.; Kim, H.I.; Yen, N.T.; Lee, K.H.; Oh, I.; Kang, J.S.; Choi, M.S.; Kim, I.S.; et al. Diversification of Orientia tsutsugamushi genotypes by intragenic recombination and their potential expansion in endemic areas. PLoS Negl. Trop. Dis. 2017, 11, e0005408. [Google Scholar] [CrossRef]
- Sonthayanon, P.; Peacock, S.J.; Chierakul, W.; Wuthiekanun, V.; Blacksell, S.D.; Holden, M.T.; Bentley, S.D.; Feil, E.J.; Day, N.P. High rates of homologous recombination in the mite endosymbiont and opportunistic human pathogen Orientia tsutsugamushi. PLoS Negl. Trop. Dis. 2010, 4, e752. [Google Scholar] [CrossRef]
- Stenos, J.; Graves, S.; Izzard, L. Rickettsia. In PCR for Clinical Microbiology—An Australian and International Perspective; Schuller, M., Sloots, T.P., James, G.S., Halliday, C.L., Carter-Wientjes, C., Eds.; Springer: Dordrecht, The Netherlands, 2010. [Google Scholar]
- Unsworth, N.B.; Stenos, J.; Faa, A.G.; Graves, S.R. Three rickettsioses, Darnley Island, Australia. Emerg. Infect. Dis. 2007, 13, 1105–1107. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gautam, R.; Parajuli, K.; Tadepalli, M.; Graves, S.; Stenos, J.; Sherchand, J.B. Scrub Typhus and Molecular Characterization of Orientia tsutsugamushi from Central Nepal. Pathogens 2021, 10, 422. https://doi.org/10.3390/pathogens10040422
Gautam R, Parajuli K, Tadepalli M, Graves S, Stenos J, Sherchand JB. Scrub Typhus and Molecular Characterization of Orientia tsutsugamushi from Central Nepal. Pathogens. 2021; 10(4):422. https://doi.org/10.3390/pathogens10040422
Chicago/Turabian StyleGautam, Rajendra, Keshab Parajuli, Mythili Tadepalli, Stephen Graves, John Stenos, and Jeevan Bahadur Sherchand. 2021. "Scrub Typhus and Molecular Characterization of Orientia tsutsugamushi from Central Nepal" Pathogens 10, no. 4: 422. https://doi.org/10.3390/pathogens10040422
APA StyleGautam, R., Parajuli, K., Tadepalli, M., Graves, S., Stenos, J., & Sherchand, J. B. (2021). Scrub Typhus and Molecular Characterization of Orientia tsutsugamushi from Central Nepal. Pathogens, 10(4), 422. https://doi.org/10.3390/pathogens10040422