
Parasites of the Reintroduced Iberian Lynx (Lynx pardinus) and Sympatric Mesocarnivores in Extremadura, Spain

 ,
,  ,
,  , ,
, ,  ,
,  ,
, 
Abstract

1. Introduction
2. Results
3. Discussion
4. Materials and Methods
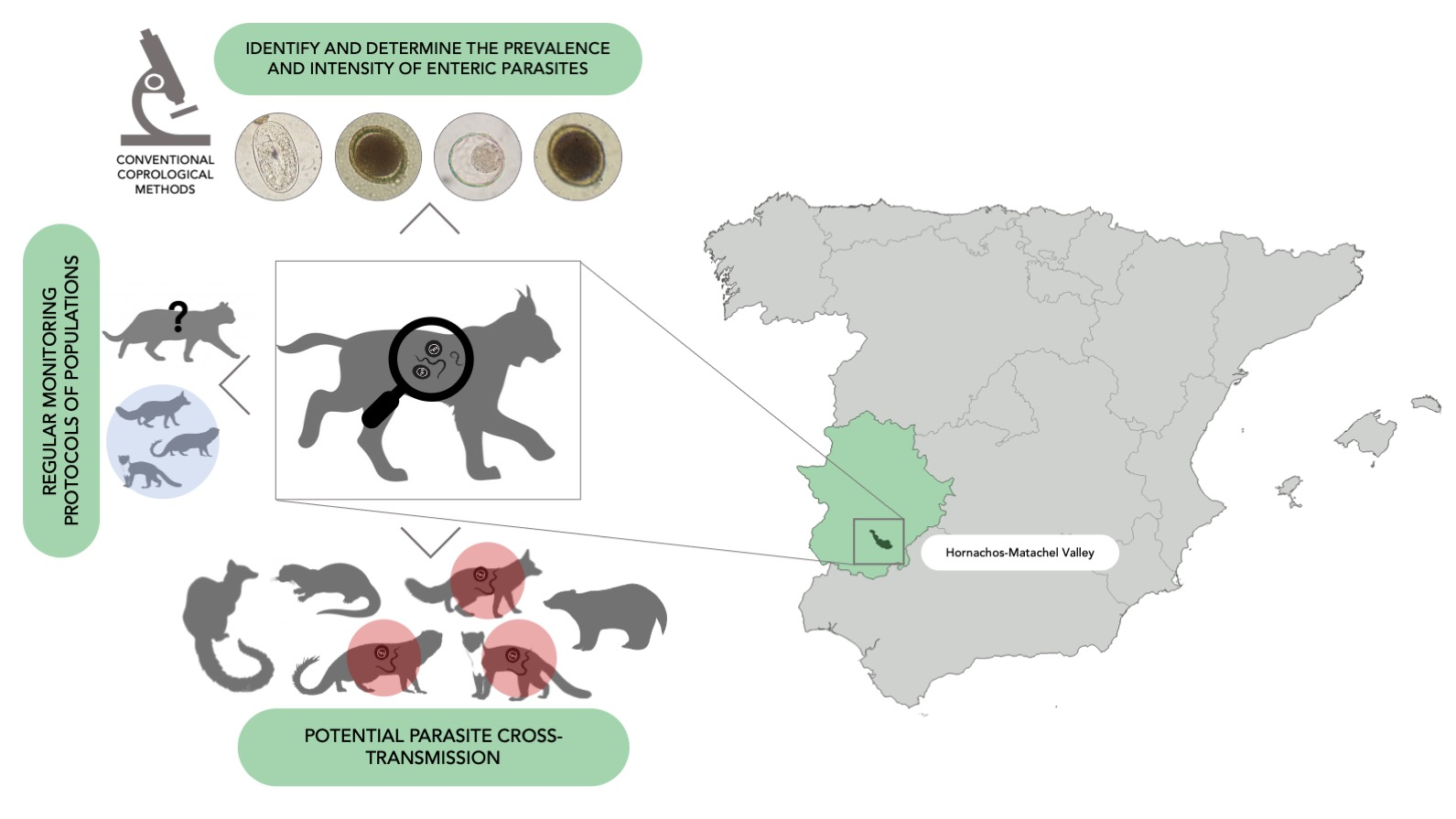
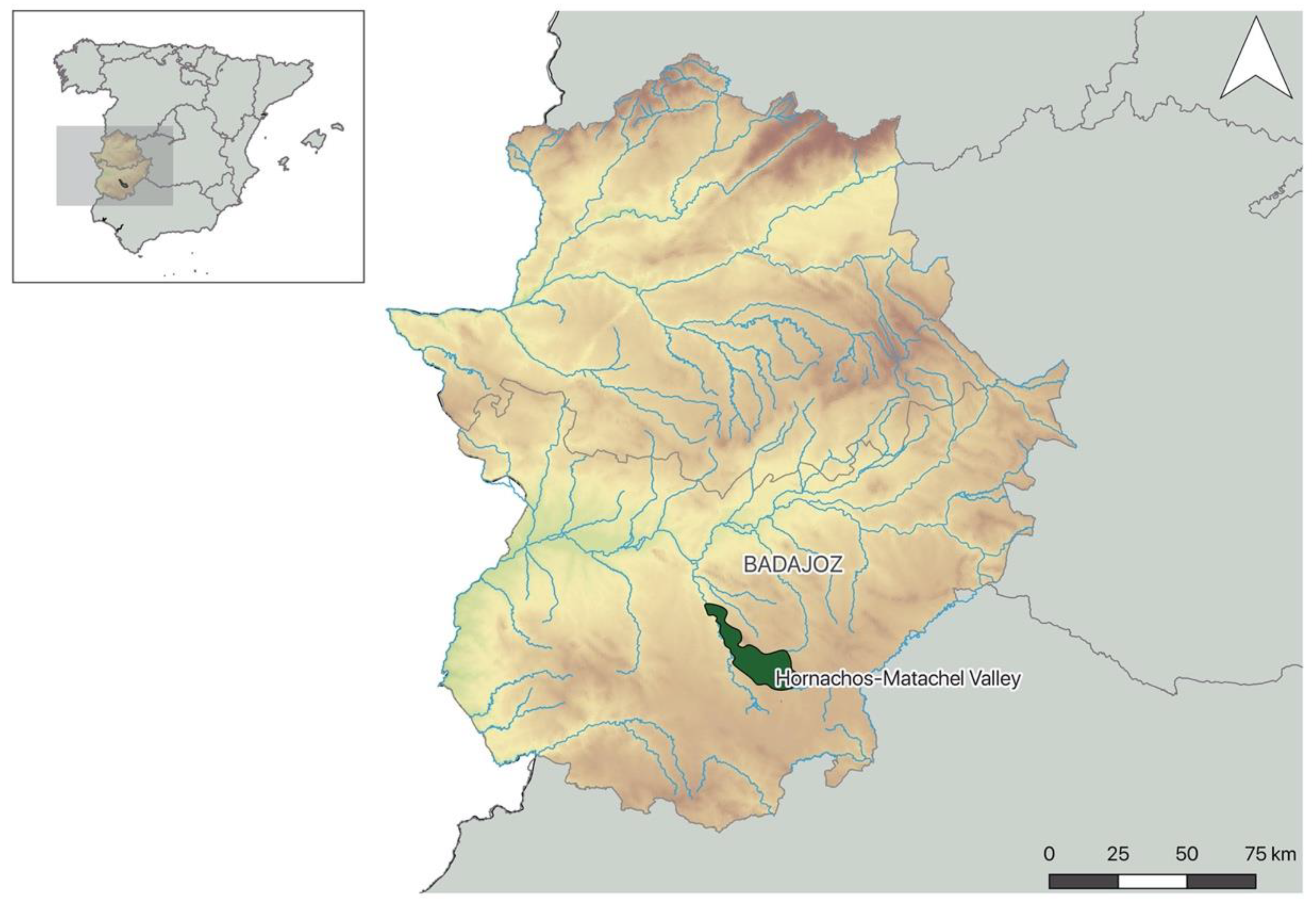
4.1. Study Area
4.2. Sample Collection and Copromicroscopic Analyses
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodríguez, A.; Calzada, J. Lynx pardinus (Errata Version Published in 2020). The IUCN Red List of Threatened Species 2015: e.T12520A174111773. 2015. Available online: https://dx.doi.org/10.2305/IUCN.UK.2015-2.RLTS.T12520A174111773.en (accessed on 7 September 2020).
- Simon, M.A.; Gil-Sanchez, J.M.; Ruiz, G.; Garrote, G.; Mccain, E.B.; Fernandez, L.; Lopez-Parra, M.; Rojas, E.; Arenas-Rojas, R.; Del Rey, T.; et al. Reverse of the decline of the endangered Iberian lynx. Conserv. Biol. 2012, 26, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Delibes, M. Patterns and causes of non-natural mortality in the Iberian lynx during a 40 year period of range contraction. Biol. Conserv. 2004, 118, 151–161. [Google Scholar] [CrossRef]
- García-Bocanegra, I.; Dubey, J.P.; Martínez, F.; Vargas, A.; Cabezón, O.; Zorrilla, I.; Arenas, A.; Almería, S. Factors affecting seroprevalence of Toxoplasma gondii in the endangered Iberian lynx (Lynx pardinus). Vet. Parasitol. 2010, 167, 36–42. [Google Scholar] [CrossRef] [PubMed]
- López, G.; López-Parra, M.; Garrote, G.; Fernández, L.; del Rey-Wamba, T.; Arenas-Rojas, R.; García-Tardío, M.; Ruiz, G.; Zorilla, I.; Moral, M.; et al. Evaluating mortality rates and causalities in a critically endangered felid across its whole distribution range. Eur. J. Wildl. Res. 2014, 60, 359–366. [Google Scholar] [CrossRef]
- Simón, M. Censo de Las Poblaciones de Lince Ibérico Anõ 2018; Technical report; Life+Iberlince, (LIFE10NAT/ES/570); Junta de Andalucía: Sevilla, Spain, 2019; p. 15. [Google Scholar]
- Roelke, M.E.; Johnson, W.E.; Millán, J.; Palomares, F.; Revilla, E.; Rodríguez, A.; Calzada, J.; Ferreras, P.; León-Vizcaíno, L.; Delibes, M.; et al. Exposure to disease agents in the endangered Iberian lynx (Lynx pardinus). Eur. J. Wildl. Res. 2008, 54, 171–178. [Google Scholar] [CrossRef]
- Meli, M.L.; Cattori, V.; Martínez, F.; López, G.; Vargas, A.; Palomares, F.; López-Bao, J.V.; Hofmann-Lehmann, R.; Lutz, H. Feline leukemia virus infection: A threat for the survival of the critically endangered Iberian lynx (Lynx pardinus). Vet. Immunol. Immunopathol. 2010, 134, 61–67. [Google Scholar] [CrossRef]
- Masot, A.J.; Gil, M.; Risco, D.; Jiménez, O.M.; Núñez, J.I.; Redondo, E. Pseudorabies virus infection (Aujeszky’s disease) in an Iberian lynx (Lynx pardinus) in Spain: A case report. BMC Vet. Res. 2017, 13, 6. [Google Scholar] [CrossRef] [PubMed]
- Oleaga, A.; García, A.; Balseiro, A.; Casais, R.; Mata, E.; Crespo, E. First description of sarcoptic mange in the endangered Iberian lynx (Lynx pardinus): Clinical and epidemiological features. Eur. J. Wildl. Res. 2019, 65, 40. [Google Scholar] [CrossRef]
- Hudson, P.J.; Dobson, A.P.; Lafferty, K.D. Is a healthy ecosystem one that is rich in parasites? Trends Ecol. Evol. 2006, 21, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C.A.; Lymbery, A.J.; Smith, A. Parasites, emerging disease and wildlife conservation. Int. J. Parasitol. 2010, 40, 1163–1170. [Google Scholar] [CrossRef]
- Poulin, R. Network analysis shining light on parasite ecology and diversity. Trends Parasitol. 2010, 26, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Carbonell, E. Gastrointestinal parasites of the Iberian lynx and other wild carnivores from central Spain. Acta Parasitol. 1998, 43, 128–136. [Google Scholar]
- Torres, J.; Garcia-Perea, R.; Gisbert, J.; Feliu, C. Helminth fauna of the Iberian lynx, Lynx pardinus. J. Helminthol. 1998, 72, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.; Palomares, F.; Ruiz de Ibanez, R.; Ortiz, J. Epidemiology of Ancylostoma spp. in the endangered Iberian lynx (Lynx pardinus) in the Doñana National Park, south-west Spain. J. Helminthol. 2004, 78, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Millán, J.; Casanova, J.C. Helminth parasites of the endangered Iberian lynx (Lynx pardinus) and sympatric carnivores. J. Helminthol. 2007, 81, 377. [Google Scholar] [CrossRef] [PubMed]
- Acosta, L.; León-Quinto, T.; Bornay-Llinares, F.J.; Simón, M.A.; Esteban, J.G. Helminth parasites in faecal samples from the endangered Iberian lynx (Lynx pardinus). Vet. Parasitol. 2011, 179, 175–179. [Google Scholar] [CrossRef]
- Jiménez, J.; Nuñez-Arjona, J.C.; Mougeot, F.; Ferreras, P.; González, L.M.; García-Domínguez, F.; Muñoz-Igualada, J.; Palacios, M.J.; Pla, S.; Rueda, C.; et al. Restoring apex predators can reduce mesopredator abundances. Biol. Conserv. 2019, 238, 108234. [Google Scholar] [CrossRef]
- León, C.I.; García-Bocanegra, I.; McCain, E.; Rodríguez, E.; Zorrilla, I.; Gómez, A.M.; Ruiz, C.; Molina, I.; Gómez-Guillamón, F. Prevalence of selected pathogens in small carnivores in reintroduction areas of the Iberian lynx (Lynx pardinus). Vet. Rec. 2017, 180, 252. [Google Scholar] [CrossRef]
- Mateo, M.; de Mingo, M.H.; de Lucio, A.; Morales, L.; Balseiro, A.; Espí, A.; Barral, M.; Barbero, J.F.L.; Habela, M.Á.; Fernández-García, J.L.; et al. Occurrence and molecular genotyping of Giardia duodenalis and Cryptosporidium spp. in wild mesocarnivores in Spain. Vet Parasitol. 2017, 235, 86–93. [Google Scholar] [CrossRef]
- Santín, M.; Calero-Bernal, R.; Carmena, D.; Mateo, M.; Balseiro, A.; Barral, M.; Barbero, J.F.L.; Habela, M.Á. Molecular Characterization of Enterocytozoon bieneusi in wild carnivores in Spain. J. Eukaryot. Microbiol. 2018, 65, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Calero-Bernal, R.; Santín, M.; Maloney, J.G.; Martín-Pérez, M.; Habela, M.Á.; Fernández-García, J.L.; Figueiredo, A.; Nájera, F.; Palacios, M.J.; Mateo, M.; et al. Blastocystis sp. Subtype diversity in wild carnivore species from Spain. J. Eukaryot. Microbiol. 2020, 67, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Thienpont, D.; Rochette, F.; Vanparjis, O.F.J. Diagnosing Helminthiasis by Coprological Examination, 2nd ed.; Janssen Research Foundation: Beerse, Belgium, 1986; pp. 35–36. [Google Scholar]
- Zajac, A.M.; Conboy, G.A. Veterinary Clinical Parasitology, 8th ed.; Wiley: West Sussex, UK, 2012; ISBN-13: 978-0-8138-2053-8/2012. [Google Scholar]
- Horton, J. Helminth-Nematode: Trichuris trichiura. In Encyclopedia of Food Safety, 1st ed.; Motarjemi, Y., Moy, G., Todd, E., Eds.; Academic Press: Washington, DC, USA; Elsevier: San Diego, CA, USA, 2014; pp. 111–115. ISBN 978-0-12-378612-8. [Google Scholar]
- Otranto, D.; Cantacessi, C.; Dantas-Torres, F.; Brianti, E.; Pfeffer, M.; Genchi, C.; Guberti, V.; Capelli, G.; Deplazes, P. The role of wild canids and felids in spreading parasites to dogs and cats in Europe. Part II: Helminths and arthropods. Vet. Parasitol. 2015, 213, 24–37. [Google Scholar] [CrossRef]
- Martínez-Carrasco, C.; De Ybáñez, M.R.; Sagarminaga, J.L.; Garijo, M.M.; Moreno, F.; Acosta, I.; Hernández, S.; Alonso, F.D. Parasites of the red fox (Vulpes vulpes Linnaeus, 1758) in Murcia, southeast Spain. Rev. Med. Vet. 2007, 158, 331–335. [Google Scholar]
- Criado-Fornelio, A.; Gutierrez-Garcia, L.; Rodriguez-Caabeiro, F.; Reus-Garcia, E.; Roldan-Soriano, M.A.; Diaz-Sanchez, M.A. A parasitological survey of wild red foxes (Vulpes vulpes) from the province of Guadalajara, Spain. Vet. Parasitol. 2000, 92, 245–251. [Google Scholar] [CrossRef]
- Traversa, D.; Morelli, S.; Di Cesare, A.; Diakou, A. Felid Cardiopulmonary Nematodes: Dilemmas Solved and New Questions Posed. Pathogens 2021, 10, 30. [Google Scholar] [CrossRef]
- Szczesna, J.; Popiołek, M.; Schmidt, K.; Kowalczyk, R. Coprological study on helminth fauna in Eurasian lynx (Lynx lynx) from the Białowieza Primeval Forest in eastern Poland. J Parasitol. 2008, 94, 981–984. [Google Scholar] [CrossRef]
- Rodan, I.; Sparkes, A.H. Preventive health care for cats. In The Cat: Clinical Medicine and Management; Little, S.E., Ed.; Saunders: Philadelphia, PA, USA; Elsevier: Amsterdam, The Netherlands, 2012; pp. 151–180. ISBN 978-1-4377-0660-4. [Google Scholar]
- Böhm, M.; White, P.C.; Chambers, J.; Smith, L.; Hutchings, M.R. Wild deer as a source of infection for livestock and humans in the UK. Vet. J. 2007, 174, 260–276. [Google Scholar] [CrossRef]
- Barrera, J.P.; Carmena, D.; Rodríguez, E.; Checa, R.; López, A.M.; Fidalgo, L.E.; Gálvez, R.; Marino, V.; Fuentes, I.; Miró, G.; et al. The red fox (Vulpes vulpes) as a potential natural reservoir of human cryptosporidiosis by Cryptosporidium hominis in Northwest Spain. Transbound. Emerg. Dis. 2020, 174, 260–276. [Google Scholar] [CrossRef]
- Nájera, F.; Sánchez-Cuerda, S.; López, G.; Rey-Wamba, T.; Rueda, C.; Panadero, J.; Palacios, M.J.; Jiménez, J. Lynx eats cat: Disease risk assessment during an Iberian lynx intraguild predation. Eur. J. Wildl. Res. 2019, 65, 1–5. [Google Scholar] [CrossRef]
- Ribas, A.; Milazzo, C.; Foronza, P.; Casanova, J.C. New data on helminths of stone marten, Martes foina (Carnivora, Mustelidae), in Italy. Helminthologia 2004, 4, 59–61. [Google Scholar]
- Olmedo, C.; Refoyo, P.; García, D.; Muñoz, B. Intestinal parasites in a population of stone marten (Martes foina) in central Spain. Arxius de Miscel lània Zoològica 2018, 16, 163–172. [Google Scholar] [CrossRef]
- Acero, F.J.; Fernández-Fernández, M.I.; Carrasco, V.M.S.; Parey, S.; Hoang, T.T.H.; Dacunha-Castelle, D.; García, J.A. Changes in heat wave characteristics over Extremadura (SW Spain). Theor. Appl. Climatol. 2018, 133, 605–617. [Google Scholar] [CrossRef]
- Jones, A.; Pybus, M.J. Taeniasis and echinococcosis. In Parasitic Disease of Wild Mammals, 2nd ed.; Samuel, W.M., Pybus, M.J., Kocan, A.A., Eds.; Iowa State University Press: Ames, IA, USA, 2001; pp. 163–165. ISBN 081382978X. [Google Scholar]
- Torchin, M.E.; Lafferty, K.D.; Dobson, A.P.; McKenzie, V.J.; Kuris, A.M. Introduced species and their missing parasites. Nature 2003, 421, 628–630. [Google Scholar] [CrossRef]
- Al-Sabi, M.N.S.; Rääf, L.; Osterman-Lind, E.; Uhlhorn, H.; Kapel, C.M.O. Gastrointestinal helminths of gray wolves (Canis lupus lupus) from Sweden. Parasitol. Res. 2018, 177, 1891–1898. [Google Scholar] [CrossRef] [PubMed]
- Lindenfors, P.; Nunn, C.L.; Jones, K.E.; Cunningham, A.A.; Sechrest, W.; Gittleman, J.L. Parasite species richness in carnivores: Effects of host body mass, latitude, geographical range and population density. Glob. Ecol. Biogeogr. 2007, 16, 496–509. [Google Scholar] [CrossRef]
- Plumb, D.C. Plumb’s Veterinary Drug Handbook, 8th ed.; Willey-Blackwell: Ames, IA, USA, 2015; pp. 1292–1294. ISBN 9781118911938. [Google Scholar]
- Millán, J.; Blasco-Costa, I. Molecular evidence of shared hookworm Ancylostoma tubaeforme haplotypes between the critically endangered Iberian lynx and sympatric domestic cats. Vet. Parasitol. 2012, 186, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, A.M.; Barros, T.; Valente, A.M.; Fonseca, C.; de Carvalho, L.M.; Torres, R.T. Prevalence of zoonotic parasites in an endangered Iberian wolf (Canis lupus signatus) population in Portugal. Mamm. Biol. 2019, 98, 154–162. [Google Scholar] [CrossRef]
- Gil-Sánchez, J.M.; Ballesteros-Duperón, E.; Bueno-Segura, J.F. Feeding ecology of the Iberian lynx Lynx pardinus in eastern Sierra Morena (southern Spain). Acta Theriol. 2006, 51, 85–90. [Google Scholar] [CrossRef]
- Ferreras, P.; Rodríguez, A.; Palomares, F.; Delibes, M. Iberian lynx: The uncertain future of a critically endangered cat. In Biology and Conservation of Wild Felids, 1st ed.; Macdonald, D.W., Loveridge, A.J., Eds.; Oxford University Press: Oxford, UK, 2010; pp. 511–524. ISBN 978-0-19-923444-8. [Google Scholar]
- Figueiredo, A.; Torres, R.T.; Pratas-Santiago, L.P.; Pérez, S.; Fonseca, C.; González, M.J.P.; Nájera, F. Reintroduction of the Iberian lynx (Lynx pardinus): A preliminary case study in Extremadura, Spain. J. Ethol. 2019, 37, 343–351. [Google Scholar] [CrossRef]
- LYNXEXSITU. The Iberian Lynx Ex Situ Conservation Programme. Available online: https://www.lynxexsitu.es/programa-en.php?sec=programa/ (accessed on 1 April 2020).
- Domínguez, G.; de la Torre, J.A. Aportaciones al conocimiento de los endoparásitos del lobo ibérico (Canis lupus signatus, Cabrera 1907) en el Norte de Burgos. Galemys 2002, 14, 49–58. [Google Scholar]
- Paradies, P.; Schnyder, M.; Capogna, A.; Lia, R.P.; Sasanelli, M. Canine angiostrongylosis in naturally infected dogs: Clinical approach and monitoring of infection after treatment. Sci. World J. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Rózsa, L.; Reiczigel, J.; Majoros, G. Quantifying parasites in samples of hosts. J. Parasitol. 2000, 86, 228–233. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Species | Iberian Lynx (Lynx pardinus) | Red Fox (Vulpes vulpes) | Egyptian Mongoose (Herpestes ichneumon) | Common Genet (Genetta genetta) | Eurasian Otter (Lutra lutra) | European Badger (Meles meles) | Stone Marten (Martes foina) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total samples | 32 | 21 | 18 | 3 | 3 | 1 | 1 | |||||||
| Parasites | n | % (CI 95%) | n | % (CI 95%) | n | % (CI 95%) | n | % (CI 95%) | n | % (CI 95%) | n | % (CI 95%) | n | % (CI 95%) |
| Nematodes | ||||||||||||||
| Ancylostomatidae | 5 | 15.6 (5.3–32.8) | 2 | 9.5 (1.2–30.4) | 1 | 5.6 (0.1–27.3) | - | - | - | - | - | - | 1 | NA a |
| Toxocara cati | 5 | 15.6 (5.3–32.8) | - | - | - | - | - | - | - | - | - | - | - | - |
| Toxascaris leonina | 4 | 12.5 (3.5–29.0) | - | - | - | - | - | - | - | - | - | - | - | - |
| Trichuris sp. | 3 | 9.4 (2.0–25.0) | - | - | - | - | - | - | - | - | - | - | - | - |
| Protozoa | ||||||||||||||
| Cystoisospora spp. | 3 | 9.4 (2.0–25.0) | 1 | 4.8 (0.1–23.8) | - | - | - | - | - | - | - | - | - | - |
| Species | Iberian Lynx (Lynx pardinus) | Red Fox (Vulpes vulpes) | Egyptian Mongoose (Herpestes ichneumon) | Common Genet (Genetta genetta) | Eurasian Otter (Lutra lutra) | European Badger (Meles meles) | Stone Marten (Martes foina) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parasites | n | Mean Intensity (Range) | n | Mean Intensity (Range) | n | Mean Intensity (Range) | n | Mean Intensity (Range) | n | Mean Intensity (Range) | n | Mean Intensity (Range) | n | Mean Intensity (Range) |
| Nematodes | ||||||||||||||
| Ancylostomatidae | - | - | 1 | 500 (500) | - | - | - | - | - | - | - | - | - | - |
| Toxocara cati | 3 | 4566.7 (100–11,050) | - | - | - | - | - | - | - | - | - | - | - | - |
| Toxascaris leonina | 1 | 200 (200) | - | - | - | - | - | - | - | - | - | - | - | - |
| Trichuris sp. | 1 | 200 (200) | - | - | - | - | - | - | - | - | - | - | - | - |
| Protozoa | ||||||||||||||
| Cystoisospora spp. | 2 | 1250 (100–2400) * | 1 | 50 (50) * | - | - | - | - | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueiredo, A.M.; de Carvalho, L.M.; González, M.J.P.; Torres, R.T.; Pla, S.; Núñez-Arjona, J.C.; Rueda, C.; Vallverdú-Coll, N.; Silvestre, F.; Peña, J.; et al. Parasites of the Reintroduced Iberian Lynx (Lynx pardinus) and Sympatric Mesocarnivores in Extremadura, Spain. Pathogens 2021, 10, 274. https://doi.org/10.3390/pathogens10030274
Figueiredo AM, de Carvalho LM, González MJP, Torres RT, Pla S, Núñez-Arjona JC, Rueda C, Vallverdú-Coll N, Silvestre F, Peña J, et al. Parasites of the Reintroduced Iberian Lynx (Lynx pardinus) and Sympatric Mesocarnivores in Extremadura, Spain. Pathogens. 2021; 10(3):274. https://doi.org/10.3390/pathogens10030274
Chicago/Turabian StyleFigueiredo, Ana M., Luís Madeira de Carvalho, María J. P. González, Rita T. Torres, Samuel Pla, Juan C. Núñez-Arjona, Carmen Rueda, Núria Vallverdú-Coll, Fernando Silvestre, Jorge Peña, and et al. 2021. "Parasites of the Reintroduced Iberian Lynx (Lynx pardinus) and Sympatric Mesocarnivores in Extremadura, Spain" Pathogens 10, no. 3: 274. https://doi.org/10.3390/pathogens10030274
APA StyleFigueiredo, A. M., de Carvalho, L. M., González, M. J. P., Torres, R. T., Pla, S., Núñez-Arjona, J. C., Rueda, C., Vallverdú-Coll, N., Silvestre, F., Peña, J., Carmena, D., Habela, M. A., Calero-Bernal, R., Fonseca, C., & Nájera, F. (2021). Parasites of the Reintroduced Iberian Lynx (Lynx pardinus) and Sympatric Mesocarnivores in Extremadura, Spain. Pathogens, 10(3), 274. https://doi.org/10.3390/pathogens10030274