Abstract
Babesiosis caused by the Babesia species is a parasitic tick-borne disease. It threatens many mammalian species and is transmitted through infected ixodid ticks. To date, the global occurrence and distribution are poorly understood in questing ticks. Therefore, we performed a meta-analysis to estimate the distribution of the pathogen. A deep search for four electronic databases of the published literature investigating the prevalence of Babesia spp. in questing ticks was undertaken and obtained data analyzed. Our results indicate that in 104 eligible studies dating from 1985 to 2020, altogether 137,364 ticks were screened with 3069 positives with an estimated global pooled prevalence estimates (PPE) of 2.10%. In total, 19 different Babesia species of both human and veterinary importance were detected in 23 tick species, with Babesia microti and Ixodes ricinus being the most widely reported Babesia and tick species, respectively. Regardless of species, adult ticks with 2.60% had the highest infection rates, while larvae had the least with 0.60%. Similarly, female ticks with 4.90% were infected compared to males with 3.80%. Nested-polymerase chain reaction (PCR) 2.80% had the highest prevalence among the molecular techniques employed. In conclusion, results obtained indicate that Babesia species are present in diverse questing tick species at a low prevalence, of which some are competent vectors.
1. Introduction
Both Theileria and Babesia species belong to the order Piroplasmida, are widely distributed and are among the economically important tick-borne hemoparasites of mammals [1]. Babesiosis has been well-known since the 19th century and is distributed worldwide as a disease of veterinary importance in cattle, sheep, pigs, dogs, and horses and in recent times has attracted attention as a zoonotic infection in humans [2,3].
Babesia is second only after Trypanosomes globally as the commonly found hemoparasites in the blood of mammals [4]. In 1888, Victor Babes, a Romanian biologist, was the first to discover the presence of intra-erythrocytic microorganisms in the blood of cattle, and he later observed similar intra-erythrocytic organisms in the blood of sheep [5]. A few years later, these microorganisms, which were later named “Babesia”, were noted in the blood of cattle in the United States [6]. These microorganisms in cattle were named Babesia bovis and B. bigemina, and in sheep, B. ovis [7]. Ever since, different species of Babesia have been observed parasitizing the blood of domestic animals. Over 100 species have been described thanks to the advances in microscopy, cell culture, and molecular techniques [1,3]. The clinical manifestations of babesiosis vary considerably across different animal species, but abortions, decreased milk and meat production, and mortality have been observed [8].
Furthermore, human babesiosis was first documented in the former Yugoslavia republic in 1957 [9]. Babesiosis in humans is becoming a public health concern as several species, including B. microti, B. divergens and B. venatorum, can infect humans accidentally, causing disease [8]. Babesia microti infections are less acute compared with B. divergens, while those due to B. venatorum are milder [10]. Affected persons are often asymptomatic except in immunocompromised individuals where the outcome can be fatal [8,11]. Clinical complications, such as hemolysis, acute respiratory distress and multiorgan malfunctioning leading to death have been observed [12].
Ixodid ticks are obligate hematophagous acarines, which feed on a wide variety of hosts, and over 700 species have been described [13]. To complete their life cycle, ticks must look for suitable hosts. Therefore, newly hatched larvae, nymphs and adults that are unfed need to seek a host for a blood meal for their further development into the next stage [14]. Detection and attachment to potential hosts in Ixodidae can be achieved through three major behavioral patterns: hunting, tick-host cohabitation, and questing [15].
Species of questing ticks within the genera Ixodes, Dermacentor and Haemaphysalis have been described and collected for the detection of tick-borne pathogens. Other species within the genus Rhipicephalus and Hyalomma have also been collected from the environment [16]. Questing ticks can be collected principally by flagging or dragging, among other methods, including trapping using baits (e.g., carbon dioxide) [14]. Ixodid ticks are the primary vectors of Babesia, but the parasites are sustained in a complex system of animal reservoirs and tick vectors [17,18]. In Ixodid ticks, the sexual phase of the life cycle of Babesia typically takes place acquiring and transmitting the parasites during blood meals from their host [19,20]. Transovarial transmission is exclusive within the Babesia sensu stricto evolutionary lineage, thereby allowing the pathogens to perpetuate their long-term persistence in ticks and serving as parasite reservoirs when vertebrate hosts are absent [18,20].
Ixodes ricinus is the most common tick, widely distributed in Europe (Western Palearctic), while the focal distribution of Dermacentor reticulatus has been observed [17,21,22,23,24,25]. Other species like I. scapularis are common in the United States of America [26,27], I. ovatus and Hemaphysalis longicornis in East Asia [28,29,30] and I. persulcatus in Europe (Russia) and parts of Central and Northern Asia [31,32]. Other species of Rhipicephalid ticks have also been reported globally [8,33,34].
Major interest in the role of questing ticks as vectors of pathogens of zoonotic importance began to emerge in the early 2000s. In questing ticks, aside from B. microti, which has been well reported in Europe, Asia, and America with varying infection rates [35,36,37,38], B. divergens and B. venatorum have been exclusively reported in Europe in the last two decades [17,39,40]. Other species of Babesia that infect domestic animals and that have been detected in questing ticks include B. canis [24,41], B. odocoilei [26,42], B. ovata [29,43], B. bigemina [8,43], B. bovis [43,44], B. caballi [41,45], B. capreoli [17,46,47] and many more.
In the last two decades, several individual studies around the world attempted to screen for the presence of Babesia species in questing ticks using molecular techniques, but no attempt has been made to synchronize the results from all these studies. Assessing the global state of the pathogen prevalence in unfed host-seeking ticks is essential to develop effective control measures. Therefore, in this study, we undertook a comprehensive assessment to determine the occurrence of Babesia species in unfed host-seeking ticks collected from vegetation while using globally published epidemiological data. To achieve the above aim, we evaluated prevalence rates according to tick species, region of sampling, life stages of ticks, sex of adult ticks, sampling years and molecular detection techniques.
2. Results
2.1. Literature Search and Eligible Studies
A total of 4359 relevant articles were identified following a search for all four databases using the procedure enumerated in Figure 1. After the removal of duplicates, we were left with 2826 studies for further review. A careful review of the titles and abstracts was done, and a total of 122 full-text articles were downloaded for detailed review. In total, 18 studies were excluded for various reasons. These included (i) the exact number of positive Babesia isolates were not clearly stated (n = 5), (ii) non-separation of the number of positive isolates of Babesia from questing ticks and other vertebrate hosts/feeding ticks (n = 4), (iii) incomplete information on tick collections (n = 3), (iv) lack of delineation of the results of positive Babesia species from other piroplasms (n = 2), and (v) no information on the number of tick DNA used for polymerase chain reaction (PCR) screening (n = 2), (vi) study with samples size below 40 (n = 2). One hundred and four (104) studies were further subjected to the quantitative synthesis. The quality assessment score (QAS) from the Joanna Briggs Institute (JBI) critical appraisal ranges from 6 to 8 out of a possible score of 9, equivalent to 66.7–88.89% in 100 out of the 104 included studies. Only 4 studies had a score of 5 (55.67%) (Table 1; Supplementary Table S2).
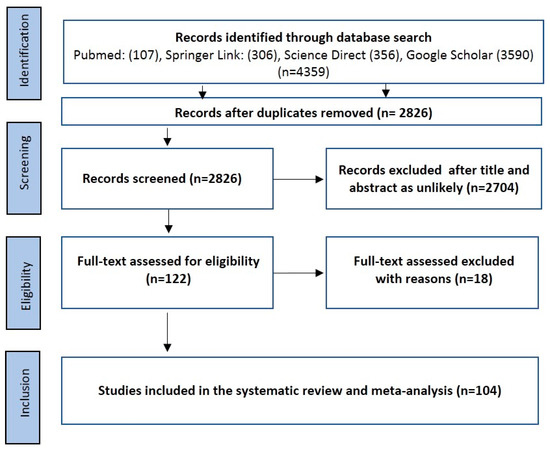
Figure 1.
Preferred reporting items for systematic reviews and meta-analyses (PRISMA) flowchart used in the selection of eligible studies.

Table 1.
Characteristics of all 104 studies used in the meta-analysis of molecular Babesia detection in questing ticks.
2.2. Characteristics of Eligible Studies
The characteristics of all eligible studies comprising of 137,364 ticks from 104 studies across different regions of the world are presented in Table 1. Included studies were from Europe (n = 78), North America (n = 13), Asia (n = 12), and Africa (n = 1). All eligible studies were carried out using molecular techniques to screen for tick-borne pathogens with particular reference to Babesia species. The prevalence for all the individual studies was computed and presented in Table 1. Individually, apart from a few studies, which recorded a 0% prevalence, the majority of the studies ranges from 0.25% to 12.96%, with a median of 1.78%. There were two studies with a prevalence of 20.65% and 21.67% and another two studies with a prevalence of 51.04% and 58.33% (Table 1). The majority of the studies were carried out from the year 2000 onward, with only one study undertaken in 1985.
2.3. Pooling, Heterogeneity and Subgroup Analysis
2.3.1. Prevalence Based on Tick Species, Life Stages, Sex, and Diagnostic Technique
The overall and subgroup prevalence estimates of Babesia spp. based on tick species, life stages, sex and diagnostic technique, including confidence intervals and statistical parameters, are presented in Table 2. Globally, the overall pooled prevalence estimated (PPE) for Babesia species in questing ticks was 2.10% for all studies with 3069 positive cases from a total of 137,364 ticks screened and substantial study heterogeneity was observed (Table 2; Figure 2). Babesia species were detected in 23 different tick species within 4 genera Ixodes (5 species), Dermacentor (4 species), Rhipicephalus (4 species), Haemaphysalis (9 species) and Hyalomma (1 species) (Table 2). Ixodes ricinus was the most collected tick species with over 74,802 ticks in number and 1756 positive cases with PPE at 2.40% (Table 2; Figure 3). Other tick species included: I. persulcatus with PPE at 1.50%, I. scapularis at 4.10%, D. reticulatus at 2.10%, and H. longicornis at 4.30% (Table 2).
Table 2.
Pooled minimum infection rate (MIR) estimates of Babesia spp. in questing ticks based on tick species, life stages, sex, and diagnostic technique.
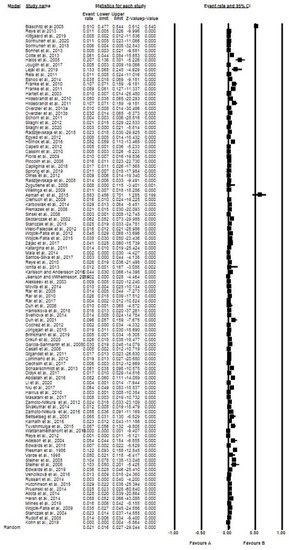
Figure 2.
Forest plot showing the pooled prevalence of Babesia species globally. N.B. The squares show the individual point estimate. The diamond at the base indicate the pooled estimates from the total studies. Event rate: the frequency of occurrence of an event in a population, and it takes into account the possibility of an event occurring several times in an individual.
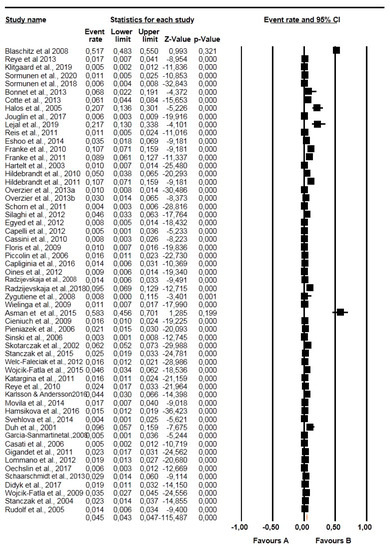
Figure 3.
Forest plot showing the prevalence of Babesia species in questing Ixodes ricinus in Europe. N.B. The squares show the individual point estimate. The diamond at the base indicate the pooled estimates from the total studies. Event rate: is the frequency of occurrence of an event in a population, and it takes into account the possibility of an event occurring several times in an individual.
Other tick species that were reported, but no Babesia species were detected: H. sp. 1 & 2 [8]; H. bispinosa [28]; Hy. spp. [109]; H. hystricis and H. kitaokai [110]; Amblyomma testudinarium [110]; I. nipponensis [110]; I. turdus [37,110]; I. tanuki [37]; H. douglasi [29,37]; H. megaspinosa [29]; H. wellingtoni [112].
With regard to tick life stages, we observed an increasing infection rate from larvae with 0.60% to nymphs with 1.70% and the highest in adults with 2.60% (Table 2). Statistically significant differences (p < 0.0001) were observed across the different life stages. Additionally, the infection rate between the adult and larva was significantly different (p = 0.0033). The PPE was significantly (p = 0.0211) higher in the females with 4.90% compared to the males with 3.60% (Table 2).
Six different molecular diagnostic techniques were employed in all the included studies, with conventional PCR being the most widely utilized in 66 studies with a PPE of 1.90%. Others include nested-PCR with 2.80% and qPCR with 1.70% (Table 2, Figure 4).
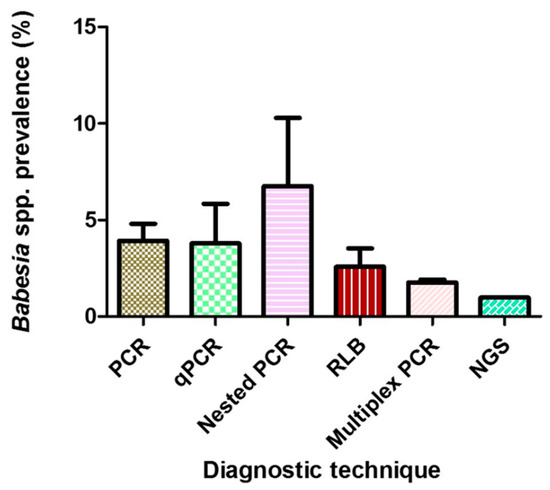
Figure 4.
Mean prevalences of Babesia species globally using different diagnostic techniques. Error bars, standard errors of the means.
2.3.2. Prevalence Based on Babesia Species, Region, and Sampling Periods
Globally, 19 different Babesia species were identified in ticks, with B. microti being the most observed species in 46 studies with a PPE of 1.90% (Table 3; Figure 5). This was followed by B. venatorum with 0.90% and B. divergens with 0.40%, which were exclusively found in ticks from Europe except for one study from Mongolia where B. venatorum DNA was amplified (Table 3). The prevalence of B. ovata was 0.60%, and B. spp. Xinjiang with 6.70% was observed only in ticks collected from Asia (Table 3).
Table 3.
Pooled MIR estimates of Babesia in questing ticks based on Babesia species, region, and sampling periods.
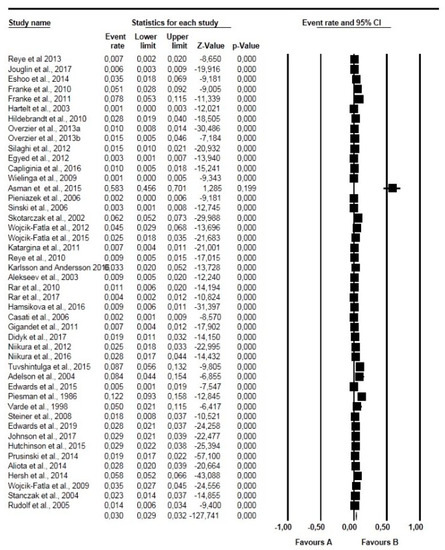
Figure 5.
Forest plot showing the prevalence of Babesia microti globally. N.B. The squares show the individual point estimate. The diamond at the base indicate the pooled estimates from the total studies.
According to region, Europe accounted for the majority of the studies (n = 78) with a PPE of 1.90% compared with Asia (n = 12) with a PPE of 2.00% (Table 3). North America had the highest PPE of 4.30% (Table 3). A single study was eligible from Africa, but none of the ticks was positive for Babesia spp.
We observed a statistically significant (p < 0.001) downward trend with respect to the PPE, with the highest being in period 1 (1992–1997) and the lowest in period 5 (2015–2020) (Table 3).
2.3.3. Species Diversity of Babesia within Different Tick Species
The results of the distribution of different Babesia species according to the different tick species are presented in Figure 6. Ixodes ricinus was associated with 9 different Babesia spp. with B. microti and B. venatorum having the highest number of isolates: 523 and 359, respectively (Figure 6). Furthermore, I. persulcatus and I. scapularis ticks were associated with 5 and 3 different Babesia species, respectively, with a total of 911 Babesia isolates shared between both ticks. Additionally, B. microti accounted for 746 Babesia isolates in I. scapularis. Finally, D. reticulatus was associated with 6 different Babesia species, with B. canis being the highest with 126 isolates (Figure 6).
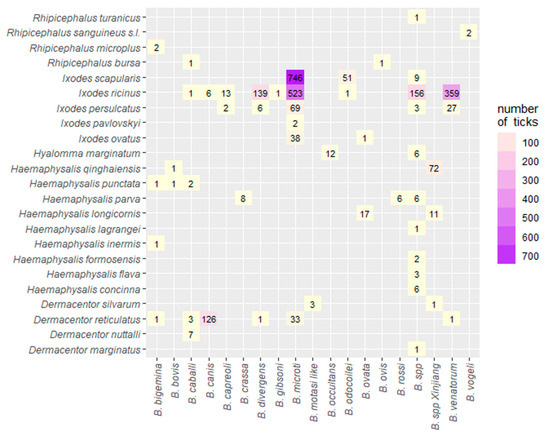
Figure 6.
Heat map of the association of different Babesia species in different tick species globally.
2.4. Spatial Distribution of Eligible Studies
In total, the results for 36 individual countries across four continents are presented in Table 4. In Europe, Poland and Germany had the highest number of eligible studies with 13 and 12 entries, each with PPE of 3.40% and 2.20%, respectively (Table 4). In addition, United States had 12 eligible studies with a PPE of 4.30%. Some other countries, including France, Russia, and Switzerland, have a PPE of 3.30%, 1.20% and 1.50%, respectively. A map with the spatial distribution of Babesia spp. across the different countries in Europe in different tick species is shown in Figure 7.
Table 4.
Prevalence estimates of Babesia in questing ticks based on country.
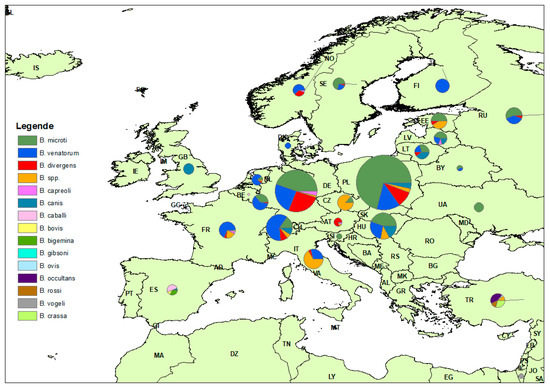
Figure 7.
Distribution of Babesia species in different tick species across Europe.
2.5. Publication Bias
The funnel plots and their corresponding bias coefficient (Begg and Mazumdar rank) for the estimation of the overall pooled MIR for published studies (Z = −48.00, p = 0.446) provides no evidence for the presence of publication bias among the eligible studies globally. For a few subgroup analyses, significant publication bias was observed for studies used for the computation of B. canis (Z = −35.00, p = 0.05), B. divergens (Z = −72.00, p = 0.01) and B. microti (Z = −203.00, p = 0.02). Additionally, mild bias was observed in studies from Asia (Z = −32.00, p = 0.014).
3. Discussion
3.1. Babesia Species in Ticks with Medical Importance
With the dawn of DNA-based techniques, molecular characterization has fostered the description and classification of new Babesia species. Therefore, the list of new species of Babesia continues to increase. In an attempt to synchronize the results from diverse epidemiological surveys for Babesia piroplasms in unfed host-seeking ticks comprising all live stages collected from vegetation across the globe, we undertook a systematic review and meta-analysis to estimate the pooled prevalence using random effect models.
Undoubtedly, Babesia microti was the most prevalent and widespread species of Babesia found in questing ticks in this study. DNA of B. microti has been detected in Europe, North America, and Asia with a PPE of 1.90%. This finding is comparable to the individual prevalence rates reported in previous studies [27,38,107,117]. Higher prevalence rates above 5.00% have also been reported in several other countries like United States [114,121], Poland [79,82] and Mongolia [32].
Babesia microti, B. duncani, B. divergens and B. venatorum are all regarded as zoonotic Babesia species. Clinically, most infected individuals are asymptomatic but could register lethal evolution depending on the species of Babesia and immunocompetence of the patient [18]. It is important to note that B. microti is responsible for most cases of human babesiosis and with great impact in North America but rare in Europe and Asia [18]. In Europe, both B. divergens and B. venatorum (formerly Babesia spp. EU1) are the predominant species causing human babesiosis. Interestingly, no study reported the detection of B. duncani in questing ticks. However, a recent report suggests the possible role of larval forms of D. albipictus as a possible vector of B. duncani transmission [122].
With the exception of one study from Mongolia [111], studies reporting the detection of B. divergens and B. venatorum were exclusively found in Europe with a PPE below 1.00%. This finding is comparable to the reports from over 70% of studies reporting the detection of this Babesia species in Europe [17,47,48,65,106]. The widespread presence of these species of Babesia of zoonotic importance in questing ticks has public health implications, especially in recreational parks during the period of tick activity. Therefore, humans could be exposed to pathogens with tick bites. Alternatively, blood transfusion-associated transmission has been reported in endemic areas, and it is regarded as the most common way of transmission in North America [123]. Therefore, Giemsa stained blood, serological testing or the use of PCR may significantly reduce the likelihood for transmission to occur by blood transfusion in endemic areas. Naturally, B. microti and B. divergens parasitize microtine rodents and cattle, respectively, these hosts being regarded as their reservoir [12]. On the other hand, B. venatorum is maintained naturally in wild cervids (deer), while the mule deer (Odocoileu hemionus) and possibly other species of wild ungulates in western North America may be the primary reservoir for B. duncani [122].
3.2. Babesia Species in Ticks with Veterinary Importance
Several species of Babesia are causing babesiosis in animals, including B. bovis, B. bigemina, B. occultans, B. divergens, B. ovata, B. odocoilei and B. capreoli (large ruminants and deer); B. caballi (equines); B. crassa, B. ovis, B. motasi-like and B. spp. Xinjiang (small ruminants), and B. vogeli, B. canis, B. rossi and B. gibsoni (canines). These species were observed in questing ticks across several regions. Of these species, some were observed to be geographically restricted (like B. ovata and B. spp. Xinjiang in Japan and China, respectively), in addition to uncharacterized Babesia species. The PPE for animal babesiosis in questing ticks ranges between 0.30% and 1.50%, with the exception of B. spp. Xinjiang with a PPE of 6.70%. These low prevalences are comparable to the infection rates reported for individual studies [24,29,41,43,47,68,108].
The PPE for B. canis was low, comparable to the prevalence reported in ticks from Slovakia [97], Russia [22] and Germany [24]. Furthermore, we observed that with the exception of B. canis, the agent of canine babesiosis, all other species of Babesia causing babesiosis in dogs were only reported separately, in one study each. Nonetheless, B. canis was reported in 14 studies from Europe. Therefore, B. canis appears to be the principal agent of canine babesiosis in Europe. In autochthonous cases where clinical canine babesiosis was reported, flagged ticks (Dermacentor reticulatus) in surrounding areas were positive to B. canis [23,41]. Additionally, in the majority of the studies (about 78%), B. canis DNA was reported in D. reticulatus tick, which is a competent vector for the protozoan parasite and is frequently found in urban biotypes in Europe [21].
Babesia caballi, one of the etiological agents of equine piroplasmosis, was observed at a low infection rate. The DNA of B. caballi was observed in R. bursa [44], D. nutalli [45] and D. reticulatus [41,44]. In the latter studies, both B. caballi and B. canis were detected in D. reticulatus ticks. Interestingly, both B. canis and B. caballi can be maintained for several generations in D. reticulatus ticks [41].
The PPE for agents of small ruminant’s babesiosis in questing ticks is consistent with reports from other individual studies where they occur at a very low prevalence [43,108]. Unlike B. motasi in Europe, B. spp. Xinjiang is known to principally infect sheep in China. From all available evidence, their presence in questing ticks is very low. Nonetheless, this Babesia spp. (B. spp. Xinjiang) has been amplified from blood samples from sheep and goats in China [28]. Earlier studies reported that Hy anatolicum anatolicum is the principal and competent vector [124]. The detection of B. spp. Xinjiang in H. longicornis and H. qinghaiensis, which are widespread in China, has raised several questions of their potential as vectors, but this remains speculative, and further studies will be required to verify this claim [28]. Additionally, B. crassa was detected in questing H. parva ticks in Turkey [108].
We observed seven species of bovine/cervid Babesia in host-seeking ticks. Unlike the virulent B. bovis and B. bigemina, B. ovata is of lower pathogenicity in cattle [29] and is one of the geographically restricted species of Babesia, similar to B. spp. Xinjiang in China. Babesia ovata is endemic in Japan and principally infects cattle [29]. H. longicornis is a known competent tick vector that can transmit the protozoan parasite transovarially [29,125], and further studies are needed to ascertain the probable role of I. ovatus that was observed to harbor this Babesia species. Additionally, B. occultans DNA was reported in Hy. marginatum, which is a known competent vector with empirical evidence from natural transovarial transmission [108], and transstadial survival [126]. Therefore, transstadial persistence of B. occultans in Hy. marginatum has been attributed to transovarial transmission of the pathogen [108], as only the adult ticks feed on the blood from cattle [127].
3.3. Ticks as Vectors of Babesia Species
Ixodes ricinus was the most abundant tick species in this study. This is not surprising considering that majority of the studies were from Europe, where this tick is predominant and a vector of several pathogens of protozoan, viral and bacterial agents of veterinary and medical importance [128,129]. Reports from various studies indicate that this tick is mostly found in urban and peri-urban areas in city parks, gardens, forest patches and litter layers [129]. Forested areas and particularly mixed and deciduous forests provide a sheltered canopy, and this tick species thrives due to the microclimates provided [128,130]. Due to climate change, current evidence points to the increasing distribution of I. ricinus steadily towards higher latitudes and altitudes. This was obvious in this study as several works were found investigating the presence of Babesia pathogen in questing tick in Sweden [92,93] and Finland [50]. Furthermore, I. ricinus harbors diverse Babesia species, which have been reported in Western Europe [46,52,73], Eastern Europe [98,99], Central Europe [39,104], Scandinavia [70,71,92], Southern Europe [44,64] and Balkan Peninsula [100], with varying prevalence and spread across the continent.
Other species within the genus Ixodes, such as I. persulcatus, I. ovatus and I. pavlovskyi, were reported in the Northern Hemisphere precisely in Russia and parts of southern Eurasia to harbor Babesia spp. at a prevalence ranging from 0.30 to 1.60%. According to [131], I. persulcatus ticks are closely related to I. pavlovskyi. For now, the vector competence of I. pavlovskyi is largely unknown. Nonetheless, I. persulcatus has been implicated as a possible competent vector for B. divergens [132].
Ixodes scapularis is widely distributed in the northeast, upper Midwest, mid-Atlantic and southeast states of the United States as well as in Canada [26,133] and was observed to be the major tick vector reported from North America. The PPE of Babesia spp. in this tick was low at 3.60%, comparable to the prevalence reported from other individual studies [118,120]. Higher prevalence has been reported in other parts of America [113,117]. Both B. microti and B. odocoilei are Babesia spp. found to be associated with this tick species causing human and cervid (white-tailed deer) babesiosis, respectively. The vector competence for B. odocoilei is unknown, but I. scapularis has been involved [42].
Haemaphysalis longicornis was reported in five studies, all from eastern Asia, where this tick species is native and originated from. The PPE was low to moderate at 4.3%. This tick species was observed to harbor B. ovata in Japan [29] and B. spp. Xinjiang in China [28]. Therefore, babesiosis in cattle and sheep, respectively, in that region is believed to be caused by B. ovata and B. spp. Xinjiang is probably transmitted by H. longicornis.
3.4. Association between Ticks and Babesia Including Other Factors
Ixodid tick species play a crucial role in the epidemiology of babesiosis. Reports of the detection of Babesia DNA may not necessarily denote evidence of vector competence, whether in unfed or engorged ticks [134]. In transovarial transmission, most Babesia species invade the tick ovaries and persist in the larvae. Consequently, infection is transmitted vertically. The acquisition of the parasites (Babesia species) from their respective host by either the larvae or nymphs is referred to as transstadial transmission.
Furthermore, of all tick species in this study, I. ricinus had the highest association with several Babesia species with three and six species of human and veterinary importance, respectively. This tick is a known competent vector for 3 Babesia parasites (B. divergens, B. venatorum and B. microti), causing human babesiosis [129]. Since all stages (larvae, nymph, and adult) of I. ricinus can transmit B. divergens and B. venatorum, the risk of infection is high after tick bites in humans during periods of peak tick activity. Detailed review on the association of I. ricinus with Babesia and other tick-borne pathogens can be obtained elsewhere [129,134].
The fact that the adult ticks and, by extension, female ticks were the most predominant with the highest infection rates compared with the nymphs and larvae may have some implications in transmission. In transovarial transmission involving most Babesia species, it has been asserted that only the female ticks can acquire the infection. Immature stages are less likely to become infected due to the smaller blood volumes they ingest. Furthermore, the fewer number and size of the midgut epithelial basophilic cells of immature stages, which play a role in parasite development, are believed to be an important factor as well [134]. Furthermore, evidence of transstadial transmission has been observed for some Babesia spp., but also, not all tick stages are capable of transmitting the parasite as observed for B. bovis, where only the larvae of R. annulatus can transmit. On the other hand, only the nymphal and adult stages of R. annulatus can transmit B. bigemina [134]. Additionally, many Babesia spp., including B. major, B. motasi, B. rossi, B. venatorum, B. vogeli and B. divergens, can persist from larval to their adult stages (transstadial transmission) in their competent vectors without reinfection for a minimum of one generation [134].
Female ticks had higher infection rates compared with their male counterparts. It is well known that female ticks require blood meals to develop their ovaries and lay thousands of eggs to perpetuate their existence. In addition, as earlier mentioned, the transovarial transmission is one of the utmost successful evolutionary strategies among the Apicomplexa and specifically in Babesia sensu stricto [20]. Therefore, female ticks take larger blood meals (high volume of blood) due to prolonged feeding, which may result in higher chances of infection. Furthermore, females require a higher number of blood meals for molting before reaching the adult stage.
The use of molecular-based techniques for the diagnosis and classification of Babesia species has been widely adopted due to greater sensitivity and specificity [18]. All studies used molecular-based techniques. In the various epidemiological investigation of Babesia species in questing ticks as observed in this study, several molecular approaches, including qPCR [49,120], nested-PCR [8,28,74], conventional PCR [47,57,88], reverse line blot hybridization [39,43] and more recently, next-generation sequencing [33] among other methods have been adopted. Despite the observation of differences in the prevalence rates between techniques, no statistical significance was noted. Similar findings were observed in a Euro-wide meta-analysis of Borrelia burgdorferi sensu lato prevalence in questing I. ricinus ticks [135]. The highest in the prevalence rate was nested-PCR, but it is difficult to conclude considering the fact that the number of studies that utilized this technique is comparatively fewer compared with the conventional PCR. The geospatial distribution indicates that extensive studies have been conducted in Germany, Poland, and United States. This observation could be connected with a research interest in those countries with a bias towards tick-borne diseases.
This systematic review has spawned data on the prevalence of Babesia species in questing ticks. However, some limitations were observed in our study. First, we excluded articles published in languages other than English, and hence some vital information may have been set aside. Second, our study focused only on questing ticks; therefore, areas without reported Babesia pathogen may still have the pathogen. Third, due to the use of different DNA-based techniques with varying sensitivity, some Babesia species with low detection sensitivity might have been missed. Fourth, the global prevalence was obtained from studies from four continents. Therefore, the global pooled prevalence of Babesia spp. may vary from the actual estimate, but we believe that the apparent prevalence in this study is close to normal. Fifth, the heterogeneity observed could be due to sampling error, sample size, or variation of endemicity and study design. Despite the limitations highlighted above, this study used a large number of eligible studies (n = 104) and ticks screened (137,364) from a global perspective to clearly provide a comprehensive insight and meta-analysis on the distribution of Babesia species in different questing ticks across four continents from published literature. Our results clearly indicate that these ticks harbor potentially disease-causing Babesia parasites of human and veterinary importance.
4. Material and Methods
4.1. Search Strategy
We followed the protocol as outlined by the preferred reporting items for systematic reviews and meta-analyses (PRISMA) in carrying out this systematic review and meta-analysis [136]. We searched for citations with no time restrictions through to 10 July 2020 solely in English databases of Science Direct, Springer Link, PubMed, and Google Scholar. Key operators used in the systematic search were “Babesia”, “questing ticks”, and “tick-borne pathogens”. Key terms used in the search were used individually or in combination with “AND” and/or “OR” operators. Duplicates were removed, and relevant titles and abstracts were scanned, and those articles in line with the aim of the study were downloaded.
4.2. Inclusion and Exclusion Criteria
Selected relevant articles, after the review of titles and abstract, were downloaded for further screening of the full text for eligibility. Included articles for the study must fulfill the following seven criteria, namely (i) the collected ticks must be questing ticks from vegetation, (ii) the total number of ticks screened was stated, (iii) the country of the study was known, (iv) the study screened for the presence of Babesia in questing ticks, and the number of positives/negatives was stated (v) the molecular diagnostic method employed in the study was stated (vi) for a tick species to be included in the result, at least one Babesia spp. DNA must have been amplified for that species (vii) no limit to the minimum sample size of screened ticks, but for statistical reasons, it was set at less than 40 samples. Where the exact number of the respective live stages were not clearly stated, the total number of screened ticks collected for that study was used only in the computation of the overall prevalence. Studies were excluded if (i) the exact number of positive Babesia isolates were not clearly stated, (ii) separation of the number of positive isolates of Babesia from questing ticks and other vertebrate host/feeding ticks was missing, (iii) incomplete information on tick collections (iv) lack of delineation of the results of positive Babesia species from other piroplasms (v) no information on the number of tick DNA samples used for PCR screening (vi) study with sample size below 40.
4.3. Data Cleaning
In most of the studies, the developmental stages (larva and nymphs) were pooled before pathogen detection. Therefore, we calculated the minimum infection rates (MIR) (based on the assumption of a single positive tick per pool) for all included studies to avoid overestimation of a prevalence. Consequently, the prevalence throughout reflects the MIR in ticks. With regard to the years of sampling, where sampling was undertaken over two or more years, and the results were presented separately for each year, we divided the entries accordingly. Similarly, where entries involved different tick species and countries but published on the same articles, the data were separated meticulously. For the calculation of the overall prevalence, we used data from all eligible studies incorporating the total number of ticks screened irrespective of the live stages. Overall, only tick species that showed at least one positivity to Babesia spp. were presented in Table 2. Therefore, tick species reported without any single cumulative positivity to Babesia spp. were not included in the results. Furthermore, the number of positive Babesia spp. isolates that were confirmed by good quality sequences as reported in the articles were used for the subgroup analysis (Babesia species).
4.4. Data Extraction
All studies meeting the inclusion criteria were cataloged, and data were extracted using a charting form developed by the research team. Data extracted from all the eligible studies included all the variables as contained in the inclusion criteria, such as the name of the authors, year of sampling, geographical location, the total number of ticks screened, the molecular diagnostic technique used, the life stages of the ticks, tick species, sex of the ticks, species of Babesia detected as well as the number of positive/negative Babesia isolates. The MIR was calculated according to the various subgroups.
4.5. Quality Assessment of Included Studies
The quality assessment of each article included in the study was undertaken using the Joanna Briggs Institute (JBI) critical appraisal instrument for studies with prevalence data [137]. This JBI instrument consists of nine questions, of which details are available (Supplementary Table S1). Each answer to the individual question was assigned a score of 0 or 1 for no or yes answers. When the question was not applicable to the study, not applicable (NA) was used. Results of Babesia species distribution were summarized on a country level and exported as a CSV file into ArcGis Desktop (Esri, version 10.5.1, Redlands, CA, USA). Data were visualized in pie charts per country.
4.6. Statistical Analysis
All statistical analyses were carried out using Comprehensive Meta-analysis (CMA) Version 3.0 by Biostat (Englewood, NJ, USA) unless otherwise stated. The weighted pooled minimum infection rate (MIR) and 95% confidence interval (CI) were computed. For each individual study, we recalculated the MIR (prevalence) by summing the total number of samples and positive cases irrespective of the number of tick species reported for that study. When the pooled analysis was performed, each logit event estimate undergoes a transformation within the CMA software into proportions with its corresponding 95% CI. We calculated the overall MIR as a percentage. Forest plots were used to visualize the data generated. Cochran’s heterogeneity (Q) among the included studies, as well as the percentage inverse variation (I2), was calculated using the Cochrane Q test. If I2 was ≤25%, 50% or ≥75%, then heterogeneity was described as low, moderate, or high (substantial), respectively [138]. If there was only a single study for a particular category, the positive rate was computed without heterogeneity (Q). All pooled estimates were arrived at using a random-effects model except for sex, where we used the fixed-effect model due to the homogeneity of the data. The chi-squared test was used to test for significance for all the subgroups using GraphPad Prism, version 5.04 (GraphPad Software, Inc, La Jolla, CA, USA, www.graphpad.com). p values of <0.05 were considered statistically significant unless otherwise stated. Funnel plots using visual inspection and the Beg and Mazumdar rank correlation test [139] were used for assessing the publication bias.
5. Conclusions
In this meta-analysis of pooled data on Babesia species in questing ticks from a global perspective, our findings indicate both human and animal Babesia species DNA in a variety of species of questing hard ticks with low to moderate prevalence. We reported the detection of 19 Babesia species in 23 different tick species across four continents. Adult male and female ticks had the highest infection rates compared with immature and male ticks, respectively. Ixodes ricinus was the main tick species of interest, and it is a tick species of economic importance, with B. microti being the most widely detected species of Babesia across the different regions. The information generated from this study will be helpful to the relevant stakeholders in the design and future implementation of programs aimed at controlling competent vectors against Babesia parasites.
Supplementary Materials
The following are available online at https://www.mdpi.com/2076-0817/10/2/230/s1, Table S1: JBI Critical appraisal checklist for studies reporting prevalence data; Table S2: Quality assessment scores for eligible studies.
Author Contributions
Conceptualization, T.E.O. and C.S.; methodology, T.E.O., C.R.; validation, T.E.O., C.R., S.F. and C.S.; data curation, T.E.O., C.R., S.F. and C.S.; writing—original draft preparation, T.E.O.; writing—review and editing, C.R., S.F. and C.S.; visualization T.E.O., S.F.; supervision C.S.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the current article and the supplementary material.
Acknowledgments
We thank Franz J. Conraths and Christoph Staubach (Friedrich-Loeffler-Institut, Greifswald-Insel Riems, Germany) for providing the ArcGIS software licenses and their useful comments on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef]
- Blaschitz, M.; Narodoslavsky-Gfoller, M.; Kanzler, M.; Stanek, G.; Walochnik, J. Babesia species occurring in Austrian Ixodes ricinus ticks. Appl. Environ. Microbiol. 2008, 74, 4841–4846. [Google Scholar] [CrossRef] [PubMed]
- Uilenberg, G. Babesia--a historical overview. Vet. Parasitol. 2006, 138, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Homer, M.J.; Aguilar-Delfin, I.; Telford, S.R., 3rd; Krause, P.J.; Persing, D.H. Babesiosis. Clin. Microbiol. Rev. 2000, 13, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Babes, V. L’etiologie d’une enzootie des moutons, denommee Carceag en Roumanie. (The etiology of an enzootic disease of sheep, called Carceag, in Romania). C. R. Hebd. Acad. Sci. 1892, 115, 359–361. [Google Scholar]
- Smith, T.; Kilbourne, E.L. Investigation into the Nature Causation and Prevention of Southern Cattle Fever; US Department of Agriculture, Bureau of Animal Industry: Washington, DC, USA, 1893; Volume 1, pp. 1–311.
- Starcovici, C. Bemerkungen über den durch Babes entdeckten Blutparasiten und die durch denselben hervorgebrachten Krankheiten, die seuchenhafte Hämoglobinurie des Rindes (Babes), des Texasfieber (Th. Smith) und der Carceag der Schafe (Babes). Zbl. Bakt. I. Abt. 1893, 14, 11–18. [Google Scholar]
- Li, L.H.; Wang, J.Z.; Zhu, D.; Li, X.S.; Lu, Y.; Yin, S.Q.; Li, S.G.; Zhang, Y.; Zhou, X.N. Detection of novel piroplasmid species and Babesia microti and Theileria orientalis genotypes in hard ticks from Tengchong County, Southwest China. Parasitol. Res. 2020, 119, 1259–1269. [Google Scholar] [CrossRef]
- Skrabalo, Z.; Deanovic, Z. Piroplasmosis in man; report of a case. Doc. Med. Geogr. Trop. 1957, 9, 11–16. [Google Scholar] [PubMed]
- Hunfeld, K.P.; Hildebrandt, A.; Gray, J.S. Babesiosis: Recent insights into an ancient disease. Int. J. Parasitol. 2008, 38, 1219–1237. [Google Scholar] [CrossRef] [PubMed]
- Moritz, E.D.; Winton, C.S.; Tonnetti, L.; Townsend, R.L.; Berardi, V.P.; Hewins, M.E.; Weeks, K.E.; Dodd, R.Y.; Stramer, S.L. Screening for Babesia microti in the U.S. Blood Supply. N. Engl. J. Med. 2016, 375, 2236–2245. [Google Scholar] [CrossRef]
- Gray, J.; Zintl, A.; Hildebrandt, A.; Hunfeld, K.P.; Weiss, L. Zoonotic babesiosis: Overview of the disease and novel aspects of pathogen identity. Ticks Tick Borne Dis. 2010, 1, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Guglielmone, A.A.; Robbins, R.G.; Apanaskevich, D.A.; Petney, T.N.; Estrada-Peña, A.; Horak, I. The Hard Ticks of the World (Acari, Ixodida, Ixodidae); Springer: New York, NY, USA, 2014; pp. 978–994. [Google Scholar]
- Gherman, C.M.; Mihalca, A.D.; Dumitrache, M.O.; Gyorke, A.; Oroian, I.; Sandor, M.; Cozma, V. CO2 flagging—An improved method for the collection of questing ticks. Parasit. Vectors 2012, 5, 125. [Google Scholar] [CrossRef] [PubMed]
- Petney, T.N.; Robbins, R.G.; Guglielmone, A.A.; Apanaskevich, D.A.; Estrada-Pena, A.; Horak, I.G.; Shao, R. A Look at the World of Ticks. In Progress in Parasitology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 283–296. [Google Scholar]
- Leonovich, S.A. Ontogenesis of the questing behavior of hard ticks (Ixodidae). Entomol. Rev. 2015, 95, 795–804. [Google Scholar] [CrossRef]
- Silaghi, C.; Woll, D.; Hamel, D.; Pfister, K.; Mahling, M.; Pfeffer, M. Babesia spp. and Anaplasma phagocytophilum in questing ticks, ticks parasitizing rodents and the parasitized rodents-analyzing the host-pathogen-vector interface in a metropolitan area. Parasit Vectors 2012, 5, 191. [Google Scholar] [CrossRef] [PubMed]
- Yabsley, M.J.; Shock, B.C. Natural history of Zoonotic Babesia: Role of wildlife reservoirs. Int. J. Parasitol. Parasites Wildl. 2013, 2, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.S.; Weiss, L.M. Babesia microti. In Emerging Protozoan Pathogens; Khan, N., Ed.; Taylor and Francis: Abingdon, UK, 2008; pp. 303–349. [Google Scholar]
- Jalovecka, M.; Sojka, D.; Ascencio, M.; Schnittger, L. Babesia Life Cycle—When Phylogeny Meets Biology. Trends Parasitol. 2019, 35, 356–368. [Google Scholar] [CrossRef]
- Hornok, S.; Kartali, K.; Takacs, N.; Hofmann-Lehmann, R. Uneven seasonal distribution of Babesia canis and its two 18S rDNA genotypes in questing Dermacentor reticulatus ticks in urban habitats. Ticks Tick Borne Dis. 2016, 7, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Rar, V.A.; Fomenko, N.V.; Dobrotvorsky, A.K.; Livanova, N.N.; Rudakova, S.A.; Fedorov, E.G.; Astanin, V.B.; Morozova, O.V. Tickborne pathogen detection, Western Siberia, Russia. Emerg. Infect. Dis. 2005, 11, 1708–1715. [Google Scholar] [CrossRef]
- Schaarschmidt, D.; Gilli, U.; Gottstein, B.; Marreros, N.; Kuhnert, P.; Daeppen, J.A.; Rosenberg, G.; Hirt, D.; Frey, C.F. Questing Dermacentor reticulatus harbouring Babesia canis DNA associated with outbreaks of canine babesiosis in the Swiss Midlands. Ticks Tick Borne Dis. 2013, 4, 334–340. [Google Scholar] [CrossRef][Green Version]
- Silaghi, C.; Weis, L.; Pfister, K. Dermacentor reticulatus and Babesia canis in Bavaria (Germany)-A Georeferenced Field Study with Digital Habitat Characterization. Pathogens 2020, 9, 541. [Google Scholar] [CrossRef]
- Sprong, H.; Fonville, M.; Docters van Leeuwen, A.; Devillers, E.; Ibanez-Justicia, A.; Stroo, A.; Hansford, K.; Cull, B.; Medlock, J.; Heyman, P.; et al. Detection of pathogens in Dermacentor reticulatus in northwestern Europe: Evaluation of a high-throughput array. Heliyon 2019, 5, e01270. [Google Scholar] [CrossRef]
- Milnes, E.L.; Thornton, G.; Leveille, A.N.; Delnatte, P.; Barta, J.R.; Smith, D.A.; Nemeth, N. Babesia odocoilei and zoonotic pathogens identified from Ixodes scapularis ticks in southern Ontario, Canada. Ticks Tick Borne Dis. 2019, 10, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Prusinski, M.A.; Kokas, J.E.; Hukey, K.T.; Kogut, S.J.; Lee, J.; Backenson, P.B. Prevalence of Borrelia burgdorferi (Spirochaetales: Spirochaetaceae), Anaplasma phagocytophilum (Rickettsiales: Anaplasmataceae), and Babesia microti (Piroplasmida: Babesiidae) in Ixodes scapularis (Acari: Ixodidae) collected from recreational lands in the Hudson Valley Region, New York State. J. Med. Entomol. 2014, 51, 226–236. [Google Scholar] [PubMed]
- Niu, Q.; Liu, Z.; Yang, J.; Gao, S.; Pan, Y.; Guan, G.; Luo, J.; Yin, H. Genetic characterization and molecular survey of Babesia sp. Xinjiang infection in small ruminants and ixodid ticks in China. Infect. Genet. Evol. 2017, 49, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, T.; Tattiyapong, M.; Okubo, K.; Suganuma, K.; Hayashida, K.; Igarashi, I.; Zakimi, S.; Matsumoto, K.; Inokuma, H.; Yokoyama, N. PCR detection of Babesia ovata from questing ticks in Japan. Ticks Tick Borne Dis. 2014, 5, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Zamoto-Niikura, A.; Morikawa, S.; Hanaki, K.I.; Holman, P.J.; Ishihara, C. Ixodes persulcatus Ticks as Vectors for the Babesia microti U.S. Lineage in Japan. Appl. Environ. Microbiol. 2016, 82, 6624–6632. [Google Scholar] [CrossRef]
- Rar, V.; Livanova, N.; Tkachev, S.; Kaverina, G.; Tikunov, A.; Sabitova, Y.; Igolkina, Y.; Panov, V.; Livanov, S.; Fomenko, N.; et al. Detection and genetic characterization of a wide range of infectious agents in Ixodes pavlovskyi ticks in Western Siberia, Russia. Parasit. Vectors 2017, 10, 258. [Google Scholar] [CrossRef] [PubMed]
- Tuvshintulga, B.; Sivakumar, T.; Battsetseg, B.; Narantsatsaral, S.O.; Enkhtaivan, B.; Battur, B.; Hayashida, K.; Okubo, K.; Ishizaki, T.; Inoue, N.; et al. The PCR detection and phylogenetic characterization of Babesia microti in questing ticks in Mongolia. Parasitol. Int. 2015, 64, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, A.; Hekimoglu, O.; Dincer, E.; Hagedorn, P.; Nitsche, A.; Ergunay, K. A cross-sectional screening by next-generation sequencing reveals Rickettsia, Coxiella, Francisella, Borrelia, Babesia, Theileria and Hemolivia species in ticks from Anatolia. Parasit. Vectors 2019, 12, 26. [Google Scholar] [CrossRef]
- Reye, A.L.; Arinola, O.G.; Hubschen, J.M.; Muller, C.P. Pathogen prevalence in ticks collected from the vegetation and livestock in Nigeria. Appl. Environ. Microbiol. 2012, 78, 2562–2568. [Google Scholar] [CrossRef] [PubMed]
- Aliota, M.T.; Dupuis, A.P., 2nd; Wilczek, M.P.; Peters, R.J.; Ostfeld, R.S.; Kramer, L.D. The prevalence of zoonotic tick-borne pathogens in Ixodes scapularis collected in the Hudson Valley, New York State. Vector Borne Zoonotic Dis. 2014, 14, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Franke, J.; Fritzsch, J.; Tomaso, H.; Straube, E.; Dorn, W.; Hildebrandt, A. Coexistence of pathogens in host-seeking and feeding ticks within a single natural habitat in Central Germany. Appl. Environ. Microbiol. 2010, 76, 6829–6836. [Google Scholar] [CrossRef] [PubMed]
- Zamoto-Niikura, A.; Tsuji, M.; Qiang, W.; Nakao, M.; Hirata, H.; Ishihara, C. Detection of two zoonotic Babesia microti lineages, the Hobetsu and U.S. lineages, in two sympatric tick species, Ixodes ovatus and Ixodes persulcatus, respectively, in Japan. Appl. Environ. Microbiol. 2012, 78, 3424–3430. [Google Scholar] [CrossRef] [PubMed]
- Overzier, E.; Pfister, K.; Herb, I.; Mahling, M.; Bock, G., Jr.; Silaghi, C. Detection of tick-borne pathogens in roe deer (Capreolus capreolus), in questing ticks (Ixodes ricinus), and in ticks infesting roe deer in southern Germany. Ticks Tick Borne Dis. 2013, 4, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Lommano, E.; Bertaiola, L.; Dupasquier, C.; Gern, L. Infections and coinfections of questing Ixodes ricinus ticks by emerging zoonotic pathogens in Western Switzerland. Appl. Environ. Microbiol. 2012, 78, 4606–4612. [Google Scholar] [CrossRef]
- Reis, C.; Cote, M.; Paul, R.E.; Bonnet, S. Questing ticks in suburban forest are infected by at least six tick-borne pathogens. Vector Borne Zoonotic Dis. 2011, 11, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Jongejan, F.; Ringenier, M.; Putting, M.; Berger, L.; Burgers, S.; Kortekaas, R.; Lenssen, J.; van Roessel, M.; Wijnveld, M.; Madder, M. Novel foci of Dermacentor reticulatus ticks infected with Babesia canis and Babesia caballi in the Netherlands and in Belgium. Parasit. Vectors 2015, 8, 232. [Google Scholar] [CrossRef]
- Steiner, F.E.; Pinger, R.R.; Vann, C.N.; Abley, M.J.; Sullivan, B.; Grindle, N.; Clay, K.; Fuqua, C. Detection of Anaplasma phagocytophilum and Babesia odocoilei DNA in Ixodes scapularis (Acari: Ixodidae) collected in Indiana. J. Med. Entomol. 2006, 43, 437–442. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Omar Abdallah, M.; Niu, Q.; Yu, P.; Guan, G.; Yang, J.; Chen, Z.; Liu, G.; Wei, Y.; Luo, J.; Yin, H. Identification of piroplasm infection in questing ticks by RLB: A broad range extension of tick-borne piroplasm in China? Parasitol. Res. 2016, 115, 2035–2044. [Google Scholar] [CrossRef]
- Garcia-Sanmartin, J.; Barandika, J.F.; Juste, R.A.; Garcia-Perez, A.L.; Hurtado, A. Distribution and molecular detection of Theileria and Babesia in questing ticks from northern Spain. Med. Vet. Entomol. 2008, 22, 318–325. [Google Scholar] [CrossRef]
- Battsetseg, B.; Xuan, X.N.; Ikadai, H.; Bautista, J.L.R.; Byambaa, B.; Boldbaatar, D.; Battur, B.; Battsetseg, G.; Batsukh, Z.; Igarashi, I.; et al. Detection of Babesia caballi and Babesia equi in Dermacentor nuttalli adult ticks. Int. J. Parasitol. 2001, 31, 384–386. [Google Scholar] [CrossRef]
- Overzier, E.; Pfister, K.; Thiel, C.; Herb, I.; Mahling, M.; Silaghi, C. Diversity of Babesia and Rickettsia species in questing Ixodes ricinus: A longitudinal study in urban, pasture, and natural habitats. Vector Borne Zoonotic Dis. 2013, 13, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Schorn, S.; Pfister, K.; Reulen, H.; Mahling, M.; Silaghi, C. Occurrence of Babesia spp., Rickettsia spp. and Bartonella spp. in Ixodes ricinus in Bavarian public parks, Germany. Parasit. Vectors 2011, 4, 135. [Google Scholar] [CrossRef] [PubMed]
- Reye, A.L.; Stegniy, V.; Mishaeva, N.P.; Velhin, S.; Hubschen, J.M.; Ignatyev, G.; Muller, C.P. Prevalence of tick-borne pathogens in Ixodes ricinus and Dermacentor reticulatus ticks from different geographical locations in Belarus. PLoS ONE 2013, 8, e54476. [Google Scholar]
- Klitgaard, K.; Kjaer, L.J.; Isbrand, A.; Hansen, M.F.; Bodker, R. Multiple infections in questing nymphs and adult female Ixodes ricinus ticks collected in a recreational forest in Denmark. Ticks Tick Borne Dis. 2019, 10, 1060–1065. [Google Scholar] [CrossRef]
- Sormunen, J.J.; Andersson, T.; Aspi, J.; Back, J.; Cederberg, T.; Haavisto, N.; Halonen, H.; Hanninen, J.; Inkinen, J.; Kulha, N.; et al. Monitoring of ticks and tick-borne pathogens through a nationwide research station network in Finland. Ticks Tick Borne Dis. 2020, 11, 101449. [Google Scholar] [CrossRef] [PubMed]
- Sormunen, J.J.; Klemola, T.; Hanninen, J.; Makela, S.; Vuorinen, I.; Penttinen, R.; Saaksjarvi, I.E.; Vesterinen, E.J. The importance of study duration and spatial scale in pathogen detection-evidence from a tick-infested island. Emerg. Microbes Infect. 2018, 7, 189. [Google Scholar] [CrossRef]
- Bonnet, S.; de la Fuente, J.; Nicollet, P.; Liu, X.; Madani, N.; Blanchard, B.; Maingourd, C.; Alongi, A.; Torina, A.; Fernandez de Mera, I.G.; et al. Prevalence of tick-borne pathogens in adult Dermacentor spp. ticks from nine collection sites in France. Vector Borne Zoonotic Dis. 2013, 13, 226–236. [Google Scholar] [CrossRef]
- Cotte, V.; Bonnet, S.; Cote, M.; Vayssier-Taussat, M. Prevalence of five pathogenic agents in questing Ixodes ricinus ticks from western France. Vector Borne Zoonotic Dis. 2010, 10, 723–730. [Google Scholar] [CrossRef]
- Halos, L.; Jamal, T.; Maillard, R.; Beugnet, F.; Le Menach, A.; Boulouis, H.J.; Vayssier-Taussat, M. Evidence of Bartonella sp. in questing adult and nymphal Ixodes ricinus ticks from France and co-infection with Borrelia burgdorferi sensu lato and Babesia sp. Vet. Res. 2005, 36, 79–87. [Google Scholar] [CrossRef]
- Jouglin, M.; Perez, G.; Butet, A.; Malandrin, L.; Bastian, S. Low prevalence of zoonotic Babesia in small mammals and Ixodes ricinus in Brittany, France. Vet. Parasitol. 2017, 238, 58–60. [Google Scholar] [CrossRef]
- Lejal, E.; Moutailler, S.; Simo, L.; Vayssier-Taussat, M.; Pollet, T. Tick-borne pathogen detection in midgut and salivary glands of adult Ixodes ricinus. Parasit. Vectors 2019, 12, 152. [Google Scholar] [CrossRef]
- Eshoo, M.W.; Crowder, C.D.; Carolan, H.E.; Rounds, M.A.; Ecker, D.J.; Haag, H.; Mothes, B.; Nolte, O. Broad-range survey of tick-borne pathogens in Southern Germany reveals a high prevalence of Babesia microti and a diversity of other tick-borne pathogens. Vector Borne Zoonotic Dis. 2014, 14, 584–591. [Google Scholar] [CrossRef]
- Franke, J.; Hildebrandt, A.; Meier, F.; Straube, E.; Dorn, W. Prevalence of Lyme disease agents and several emerging pathogens in questing ticks from the German Baltic coast. J. Med. Entomol. 2011, 48, 441–444. [Google Scholar] [CrossRef]
- Hartelt, K.; Oehme, R.; Frank, H.; Brockmann, S.O.; Hassler, D.; Kimmig, P. Pathogens and symbionts in ticks: Prevalence of Anaplasma phagocytophilum (Ehrlichia sp.), Wolbachia sp., Rickettsia sp., and Babesia sp in Southern Germany. Int. J. Med. Microbiol. 2004, 293, 86–92. [Google Scholar] [CrossRef]
- Hildebrandt, A.; Pauliks, K.; Sachse, S.; Straube, E. Coexistence of Borrelia spp. and Babesia spp. in Ixodes ricinus ticks in Middle Germany. Vector Borne Zoonotic Dis. 2010, 10, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, A.; Fritzsch, J.; Franke, J.; Sachse, S.; Dorn, W.; Straube, E. Co-circulation of emerging tick-borne pathogens in Middle Germany. Vector Borne Zoonotic Dis. 2011, 11, 533–537. [Google Scholar] [CrossRef]
- Kohn, M.; Krucken, J.; McKay-Demeler, J.; Pachnicke, S.; Krieger, K.; von Samson-Himmelstjerna, G. Dermacentor reticulatus in Berlin/Brandenburg (Germany): Activity patterns and associated pathogens. Ticks Tick Borne Dis. 2019, 10, 191–206. [Google Scholar] [CrossRef]
- Egyed, L.; Elo, P.; Sreter-Lancz, Z.; Szell, Z.; Balogh, Z.; Sreter, T. Seasonal activity and tick-borne pathogen infection rates of Ixodes ricinus ticks in Hungary. Ticks Tick Borne Dis. 2012, 3, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Capelli, G.; Ravagnan, S.; Montarsi, F.; Ciocchetta, S.; Cazzin, S.; Porcellato, E.; Babiker, A.M.; Cassini, R.; Salviato, A.; Cattoli, G.; et al. Occurrence and identification of risk areas of Ixodes ricinus-borne pathogens: A cost-effectiveness analysis in north-eastern Italy. Parasit. Vector 2012, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Cassini, R.; Bonoli, C.; Montarsi, F.; Tessarin, C.; Marcer, F.; Galuppi, R. Detection of Babesia EU1 in Ixodes ricinus ticks in northern Italy. Vet. Parasitol. 2010, 171, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Floris, R.; Cecco, P.; Mignozzi, K.; Boemo, B.; Cinco, M. First detection of Babesia EU1 and Babesia divergens-like in Ixodes ricinus ticks in north-eastern Italy. Parassitologia 2009, 51, 23–28. [Google Scholar]
- Piccolin, G.; Benedetti, G.; Doglioni, C.; Lorenzato, C.; Mancuso, S.; Papa, N.; Pttton, L.; Ramon, M.C.; Zasio, C.; Bertiato, G. A study of the presence of Borrelia burgdorferi, Anaplasma (previously Ehrlichia) phagocytophilum, Rickettsia, and Babesia in Ixodes ricinus collected within the territory of Belluno, Italy. Vector-Borne Zoonot. 2006, 6, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Capligina, V.; Berzina, I.; Bormane, A.; Salmane, I.; Vilks, K.; Kazarina, A.; Bandere, D.; Baumanis, V.; Ranka, R. Prevalence and phylogenetic analysis of Babesia spp. in Ixodes ricinus and Ixodes persulcatus ticks in Latvia. Exp. Appl. Acarol. 2016, 68, 325–336. [Google Scholar] [CrossRef]
- Radzijevskaja, J.; Mardosaite-Busaitiene, D.; Aleksandraviciene, A.; Paulauskas, A. Investigation of Babesia spp. in sympatric populations of Dermacentor reticulatus and Ixodes ricinus ticks in Lithuania and Latvia. Ticks Tick-Borne Dis. 2018, 9, 270–274. [Google Scholar] [CrossRef]
- Oines, O.; Radzijevskaja, J.; Paulauskas, A.; Rosef, O. Prevalence and diversity of Babesia spp. in questing Ixodes ricinus ticks from Norway. Parasit. Vectors 2012, 5, 156. [Google Scholar] [CrossRef]
- Radzijevskaja, J.; Paulauskas, A.; Rosef, O. Prevalence of Anaplasma phagocytophilum and Babesia divergens in Ixodes ricinus ticks from Lithuania and Norway. Int. J. Med. Microbiol. 2008, 298, 218–221. [Google Scholar] [CrossRef]
- Žygutienė, M.; Alekseev, A.; Dubinina, H.; Kazlauskienė, R. Evidence for a risk of tick-borne infection in the city parks of Vilnius, Lithuania. EKOLOGIJA 2008, 54, 40–43. [Google Scholar] [CrossRef]
- Wielinga, P.R.; Fonville, M.; Sprong, H.; Gaasenbeek, C.; Borgsteede, F.; van der Giessen, J.W.B. Persistent Detection of Babesia EU1 and Babesia microti in Ixodes ricinus in The Netherlands During a 5-Year Surveillance: 2003-2007. Vector-Borne Zoonot. 2009, 9, 119–121. [Google Scholar] [CrossRef]
- Asman, M.; Solarz, K.; Cuber, P.; Gasior, T.; Szilman, P.; Szilman, E.; Tondas, E.; Matzullok, A.; Kusion, N.; Florek, K. Detection of protozoans Babesia microti and Toxoplasma gondii and their co-existence in ticks (Acari: Ixodida) collected in Tarnogorski district (Upper Silesia, Poland). Ann. Agric. Environ. Med. 2015, 22, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Cieniuch, S.; Stanczak, J.; Ruczaj, A. The First Detection of Babesia EU1 and Babesia canis canis in Ixodes ricinus Ticks (Acari, Ixodidae) Collected in Urban and Rural Areas in Northern Poland. Pol. J. Microbiol. 2009, 58, 231–236. [Google Scholar]
- Karbowiak, G.; Vichova, B.; Slivinska, K.; Werszko, J.; Didyk, J.; Pet’ko, B.; Stanko, M.; Akimov, I. The infection of questing Dermacentor reticulatus ticks with Babesia canis and Anaplasma phagocytophilum in the Chernobyl exclusion zone. Vet. Parasitol. 2014, 204, 372–375. [Google Scholar] [CrossRef] [PubMed]
- Pieniazek, N.; Sawczuk, M.; Skotarczak, B. Molecular identification of Babesia parasites isolated from Ixodes ricinus ticks collected in northwestern Poland. J. Parasitol. 2006, 92, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Sinski, E.; Bajer, A.; Welc, R.; Pawelczyk, A.; Ogrzewalska, M.; Behnke, J.M. Babesia microti: Prevalence in wild rodents and Ixodes ricinus ticks from the Mazury Lakes District of north-eastern Poland. Int. J. Med. Microbiol. 2006, 296, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Skotarczak, B.; Wodecka, B.; Cichocka, A. Coexistence DNA of Borrelia burgdorferi sensu lato and Babesia microti in Ixodes ricinus ticks from north-western Poland. Ann. Agric. Environ. Med. 2002, 9, 25–28. [Google Scholar]
- Stanczak, J.; Cieniuch, S.; Lass, A.; Biernat, B.; Racewicz, M. Detection and quantification of Anaplasma phagocytophilum and Babesia spp. in Ixodes ricinus ticks from urban and rural environment, northern Poland, by real-time polymerase chain reaction. Exp. Appl. Acarol. 2015, 66, 63–81. [Google Scholar] [CrossRef] [PubMed]
- Welc-Faleciak, R.; Bajer, A.; Paziewska-Harris, A.; Baumann-Popczyk, A.; Sinski, E. Diversity of Babesia in Ixodes ricinus ticks in Poland. Adv. Med. Sci. Pol. 2012, 57, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Wojcik-Fatla, A.; Bartosik, K.; Buczek, A.; Dutkiewicz, J. Babesia microti in adult Dermacentor reticulatus ticks from Eastern Poland. Vector-Borne Zoonot. 2012, 12, 841–843. [Google Scholar] [CrossRef] [PubMed]
- Wojcik-Fatla, A.; Zajac, V.; Sawczyn, A.; Cisak, E.; Dutkiewicz, J. Babesia spp. in questing ticks from eastern Poland: Prevalence and species diversity. Parasitol. Res. 2015, 114, 3111–3116. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zajac, V.; Wojcik-Fatla, A.; Sawczyn, A.; Cisak, E.; Sroka, J.; Kloc, A.; Zajac, Z.; Buczek, A.; Dutkiewicz, J.; Bartosik, K. Prevalence of infections and co-infections with 6 pathogens in Dermacentor reticulatus ticks collected in eastern Poland. Ann. Agric. Environ. Med. 2017, 24, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Wojcik-Fatla, A.; Szymanska, J.; Wdowiak, L.; Buczek, A.; Dutkiewicz, J. Coincidence of three pathogens (Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum and Babesia microti) in Ixodes ricinus ticks in the Lublin Macroregion. Ann. Agric. Environ. Med. 2009, 16, 151–158. [Google Scholar] [PubMed]
- Stanczak, J.; Gabre, R.M.; Kruminis-Lozowska, W.; Racewicz, M.; Kubica-Biernat, B. Ixodes ricinus as a vector of Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum and Babesia microti in urban and suburban forests. Ann. Agric. Environ. Med. 2004, 11, 109–114. [Google Scholar] [PubMed]
- Katargina, O.; Geller, J.; Vasilenko, V.; Kuznetsova, T.; Jarvekulg, L.; Vene, S.; Lundkvist, A.; Golovljova, I. Detection and characterization of Babesia species in Ixodes ticks in Estonia. Vector Borne Zoonotic Dis. 2011, 11, 923–928. [Google Scholar] [CrossRef]
- Maia, C.; Ferreira, A.; Nunes, M.; Vieira, M.L.; Campino, L.; Cardoso, L. Molecular detection of bacterial and parasitic pathogens in hard ticks from Portugal. Ticks Tick Borne Dis. 2014, 5, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Santos-Silva, M.M.; Melo, P.; Santos, N.; Antunes, S.; Duarte, L.R.; Ferrolho, J.; Milhano, N.; Santos, P.T.; Domingos, A.; Santos, A.S. PCR screening of tick-borne agents in sensitive conservation areas, Southeast Portugal. Mol. Cell. Probes 2017, 31, 42–45. [Google Scholar] [CrossRef]
- Reye, A.L.; Hubschen, J.M.; Sausy, A.; Muller, C.P. Prevalence and seasonality of tick-borne pathogens in questing Ixodes ricinus ticks from Luxembourg. Appl. Environ. Microbiol. 2010, 76, 2923–2931. [Google Scholar] [CrossRef] [PubMed]
- Ionita, M.; Mitrea, I.L.; Pfister, K.; Hamel, D.; Silaghi, C. Molecular evidence for bacterial and protozoan pathogens in hard ticks from Romania. Vet. Parasitol. 2013, 196, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, M.E.; Andersson, M.O. Babesia species in questing Ixodes ricinus, Sweden. Ticks Tick Borne Dis. 2016, 7, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Jaenson, T.G.T.; Wilhelmsson, P. First records of tick-borne pathogens in populations of the taiga tick Ixodes persulcatus in Sweden. Parasit. Vectors 2019, 12, 559. [Google Scholar] [CrossRef]
- Alekseev, A.N.; Semenov, A.V.; Dubinina, H.V. Evidence of Babesia microti infection in multi-infected Ixodes persulcatus ticks in Russia. Exp. Appl. Acarol. 2003, 29, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Movila, A.; Dubinina, H.V.; Sitnicova, N.; Bespyatova, L.; Uspenskaia, I.; Efremova, G.; Toderas, I.; Alekseev, A.N. Comparison of tick-borne microorganism communities in Ixodes spp. of the Ixodes ricinus species complex at distinct geographical regions. Exp. Appl. Acarol. 2014, 63, 65–76. [Google Scholar] [CrossRef]
- Rar, V.A.; Epikhina, T.I.; Livanova, N.N.; Panov, V.V. Genetic diversity of Babesia in Ixodes persulcatus and small mammals from North Ural and West Siberia, Russia. Parasitology 2011, 138, 175–182. [Google Scholar] [CrossRef]
- Duh, D.; Slovak, M.; Saksida, A.; Stramsek, K.; Petrovec, M.; Avsic-Zupanc, T. Molecular detection of Babesia canis in Dermacentor reticulatus ticks collected in Slovakia. Biologia 2006, 61, 231–233. [Google Scholar] [CrossRef]
- Hamsikova, Z.; Kazimirova, M.; Harustiakova, D.; Mahrikova, L.; Slovak, M.; Berthova, L.; Kocianova, E.; Schnittger, L. Babesia spp. in ticks and wildlife in different habitat types of Slovakia. Parasit. Vectors 2016, 9, 292. [Google Scholar] [CrossRef]
- Svehlova, A.; Berthova, L.; Sallay, B.; Boldis, V.; Sparagano, O.A.; Spitalska, E. Sympatric occurrence of Ixodes ricinus, Dermacentor reticulatus and Haemaphysalis concinna ticks and Rickettsia and Babesia species in Slovakia. Ticks Tick Borne Dis. 2014, 5, 600–605. [Google Scholar] [CrossRef]
- Duh, D.; Petrovec, M.; Avsic-Zupanc, T. Diversity of Babesia infecting European sheep ticks (Ixodes ricinus). J. Clin. Microbiol. 2001, 39, 3395–3397. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, I.; Golovchenko, M.; Sikutova, S.; Rudenko, N.; Grubhoffer, L.; Hubalek, Z. Babesia microti (Piroplasmida: Babesiidae) in nymphal Ixodes ricinus (Acari: Ixodidae) in the Czech Republic. Folia. Parasitol. (Praha) 2005, 52, 274–276. [Google Scholar] [CrossRef] [PubMed]
- Venclikova, K.; Mendel, J.; Betasova, L.; Blazejova, H.; Jedlickova, P.; Strakova, P.; Hubalek, Z.; Rudolf, I. Neglected tick-borne pathogens in the Czech Republic, 2011–2014. Ticks Tick Borne Dis. 2016, 7, 107–112. [Google Scholar] [CrossRef]
- Cochez, C.; Lempereur, L.; Madder, M.; Claerebout, E.; Simons, L.; De Wilde, N.; Linden, A.; Saegerman, C.; Heyman, P.; Losson, B. Foci report on indigenous Dermacentor reticulatus populations in Belgium and a preliminary study of associated babesiosis pathogens. Med. Vet. Entomol. 2012, 26, 355–358. [Google Scholar] [CrossRef] [PubMed]
- Casati, S.; Sager, H.; Gern, L.; Piffaretti, J.C. Presence of potentially pathogenic Babesia sp. for human in Ixodes ricinus in Switzerland. Ann. Agric. Environ. Med. 2006, 13, 65–70. [Google Scholar]
- Gigandet, L.; Stauffer, E.; Douet, V.; Rais, O.; Moret, J.; Gern, L. Prevalence of three zoonotic Babesia species in Ixodes ricinus (Linne, 1758) nymphs in a suburban forest in Switzerland. Vector Borne Zoonotic Dis. 2011, 11, 363–366. [Google Scholar] [CrossRef]
- Oechslin, C.P.; Heutschi, D.; Lenz, N.; Tischhauser, W.; Peter, O.; Rais, O.; Beuret, C.M.; Leib, S.L.; Bankoul, S.; Ackermann-Gaumann, R. Prevalence of tick-borne pathogens in questing Ixodes ricinus ticks in urban and suburban areas of Switzerland. Parasit. Vectors 2017, 10, 558. [Google Scholar] [CrossRef] [PubMed]
- Didyk, Y.M.; Blanarova, L.; Pogrebnyak, S.; Akimov, I.; Petko, B.; Vichova, B. Emergence of tick-borne pathogens (Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum, Ricketsia raoultii and Babesia microti) in the Kyiv urban parks, Ukraine. Ticks Tick Borne Dis. 2017, 8, 219–225. [Google Scholar] [CrossRef]
- Orkun, O.; Cakmak, A.; Nalbantoglu, S.; Karaer, Z. Turkey tick news: A molecular investigation into the presence of tick-borne pathogens in host-seeking ticks in Anatolia; Initial evidence of putative vectors and pathogens, and footsteps of a secretly rising vector tick, Haemaphysalis parva. Ticks Tick Borne Dis. 2020, 11, 101373. [Google Scholar] [CrossRef]
- Harrus, S.; Perlman-Avrahami, A.; Mumcuoglu, K.Y.; Morick, D.; Eyal, O.; Baneth, G. Molecular detection of Ehrlichia canis, Anaplasma bovis, Anaplasma platys, Candidatus Midichloria mitochondrii and Babesia canis vogeli in ticks from Israel. Clin. Microbiol. Infect. 2011, 17, 459–463. [Google Scholar] [CrossRef]
- Masatani, T.; Hayashi, K.; Andoh, M.; Tateno, M.; Endo, Y.; Asada, M.; Kusakisako, K.; Tanaka, T.; Gokuden, M.; Hozumi, N.; et al. Detection and molecular characterization of Babesia, Theileria, and Hepatozoon species in hard ticks collected from Kagoshima, the southern region in Japan. Ticks Tick Borne Dis. 2017, 8, 581–587. [Google Scholar] [CrossRef]
- Karnath, C.; Obiegala, A.; Speck, S.; Essbauer, S.; Derschum, H.; Scholz, H.; Kiefer, D.; Tserennorov, D.; Dashdavaa, O.; Tsogbadrakh, N.; et al. Detection of Babesia venatorum, Anaplasma phagocytophilum and Candidatus Neoehrlichia mikurensis in Ixodes persulcatus ticks from Mongolia. Ticks Tick-Borne Dis. 2016, 7, 357–360. [Google Scholar] [CrossRef]
- Wattanamethanont, J.; Kaewthamasorn, M.; Tiawsirisup, S. Natural infection of questing ixodid ticks with protozoa and bacteria in Chonburi Province, Thailand. Ticks Tick Borne Dis. 2018, 9, 749–758. [Google Scholar] [CrossRef]
- Adelson, M.E.; Rao, R.V.; Tilton, R.C.; Cabets, K.; Eskow, E.; Fein, L.; Occi, J.L.; Mordechai, E. Prevalence of Borrelia burgdorferi, Bartonella spp., Babesia microti, and Anaplasma phagocytophila in Ixodes scapularis ticks collected in Northern New Jersey. J. Clin. Microbiol. 2004, 42, 2799–2801. [Google Scholar] [CrossRef]
- Edwards, M.J.; Barbalato, L.A.; Makkapati, A.; Pham, K.D.; Bugbee, L.M. Relatively low prevalence of Babesia microti and Anaplasma phagocytophilum in Ixodes scapularis ticks collected in the Lehigh Valley region of eastern Pennsylvania. Ticks Tick Borne Dis. 2015, 6, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Piesman, J.; Mather, T.N.; Telford, S.R., 3rd; Spielman, A. Concurrent Borrelia burgdorferi and Babesia microti infection in nymphal Ixodes dammini. J. Clin. Microbiol. 1986, 24, 446–447. [Google Scholar] [CrossRef]
- Varde, S.; Beckley, J.; Schwartz, I. Prevalence of tick-borne pathogens in Ixodes scapularis in a rural New Jersey County. Emerg. Infect. Dis. 1998, 4, 97–99. [Google Scholar] [CrossRef]
- Steiner, F.E.; Pinger, R.R.; Vann, C.N.; Grindle, N.; Civitello, D.; Clay, K.; Fuqua, C. Infection and co-infection rates of Anaplasma phagocytophilum variants, Babesia spp., Borrelia burgdorferi, and the rickettsial endosymbiont in Ixodes scapularis (Acari: Ixodidae) from sites in Indiana, Maine, Pennsylvania, and Wisconsin. J. Med. Entomol. 2008, 45, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.J.; Russell, J.C.; Davidson, E.N.; Yanushefski, T.J.; Fleischman, B.L.; Heist, R.O.; Leep-Lazar, J.G.; Stuppi, S.L.; Esposito, R.A.; Suppan, L.M. A 4-Yr Survey of the Range of Ticks and Tick-Borne Pathogens in the Lehigh Valley Region of Eastern Pennsylvania. J. Med. Entomol. 2019, 56, 1122–1134. [Google Scholar] [CrossRef]
- Russart, N.M.; Dougherty, M.W.; Vaughan, J.A. Survey of ticks (Acari: Ixodidae) and tick-borne pathogens in North Dakota. J. Med. Entomol. 2014, 51, 1087–1090. [Google Scholar] [CrossRef]
- Hutchinson, M.L.; Strohecker, M.D.; Simmons, T.W.; Kyle, A.D.; Helwig, M.W. Prevalence Rates of Borrelia burgdorferi (Spirochaetales: Spirochaetaceae), Anaplasma phagocytophilum (Rickettsiales: Anaplasmataceae), and Babesia microti (Piroplasmida: Babesiidae) in host-seeking Ixodes scapularis (Acari: Ixodidae) from Pennsylvania. J. Med. Entomol. 2015, 52, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Hersh, M.H.; Ostfeld, R.S.; McHenry, D.J.; Tibbetts, M.; Brunner, J.L.; Killilea, M.E.; LoGiudice, K.; Schmidt, K.A.; Keesing, F. Co-infection of blacklegged ticks with Babesia microti and Borrelia burgdorferi is higher than expected and acquired from small mammal hosts. PLoS ONE 2014, 9, e99348. [Google Scholar] [CrossRef]
- Swei, A.; O’Connor, K.E.; Couper, L.I.; Thekkiniath, J.; Conrad, P.A.; Padgett, K.A.; Burns, J.; Yoshimizu, M.H.; Gonzales, B.; Munk, B.; et al. Evidence for transmission of the zoonotic apicomplexan parasite Babesia duncani by the tick Dermacentor albipictus. Int. J. Parasitol. 2019, 49, 95–103. [Google Scholar] [CrossRef]
- Young, C.; Chawla, A.; Berardi, V.; Padbury, J.; Skowron, G.; Krause, P.J.; Containment, B.T.I. Preventing transfusion-transmitted babesiosis: Preliminary experience of the first laboratory-based blood donor screening program. Transfusion 2012, 52, 1523–1529. [Google Scholar] [CrossRef]
- Guan, G.; Ma, M.; Moreau, E.; Liu, J.; Lu, B.; Bai, Q.; Luo, J.; Jorgensen, W.; Chauvin, A.; Yin, H. A new ovine Babesia species transmitted by Hyalomma anatolicum anatolicum. Exp. Parasitol. 2009, 122, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Bai, Q.; Liu, G.Y.; Zhang, L.; Zhou, J.Y. Studies on the isolation and preservation of a single species of bovine haematocytozoon: The finding and isolation of Babesia ovata in China. Chin. J. Vet. Med. 1990, 16, 2–4. [Google Scholar]
- Aktas, M. A survey of ixodid tick species and molecular identification of tick-borne pathogens. Vet. Parasitol. 2014, 200, 276–283. [Google Scholar] [CrossRef]
- Hoogstraal, H. The epidemiology of tick-borne Crimean-Congo hemorrhagic fever in Asia, Europe, and Africa. J. Med. Entomol. 1979, 15, 307–417. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Pena, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasit. Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubalek, Z.; Foldvari, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Spitalska, E.; et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: New hazards and relevance for public health. Front. Public Health 2014, 2, 251. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.; Eze, J.I.; Gilbert, L.; Wint, G.R.W.; Gunn, G.; Macrae, A.; Medlock, J.M.; Auty, H. Using imperfect data in predictive mapping of vectors: A regional example of Ixodes ricinus distribution. Parasit. Vector 2019, 12, 536. [Google Scholar] [CrossRef] [PubMed]
- Livanova, N.N.; Tikunov, A.Y.; Kurilshikov, A.M.; Livanov, S.G.; Fomenko, N.V.; Taranenko, D.E.; Kvashnina, A.E.; Tikunova, N.V. Genetic diversity of Ixodes pavlovskyi and I. persulcatus (Acari: Ixodidae) from the sympatric zone in the south of Western Siberia and Kazakhstan. Exp. Appl. Acarol. 2015, 67, 441–456. [Google Scholar] [CrossRef] [PubMed]
- Friedhoff, K.T. Transmission of Babesia. In Babesiosis of Domestic Animals and Man; Ristic, M., Ed.; CRC Press: Boca Raton, FL, USA, 1988. [Google Scholar] [CrossRef]
- Eisen, R.J.; Eisen, L. The Blacklegged Tick, Ixodes scapularis: An increasing public health concern. Trends Parasitol. 2018, 34, 295–309. [Google Scholar] [CrossRef]
- Gray, J.S.; Estrada-Pena, A.; Zintl, A. Vectors of Babesiosis. Annu. Rev. Entomol. 2019, 64, 149–165. [Google Scholar] [CrossRef]
- Strnad, M.; Honig, V.; Ruzek, D.; Grubhoffer, L.; Rego, R.O.M. Europe-Wide Meta-Analysis of Borrelia burgdorferi sensu lato prevalence in questing Ixodes ricinus Ticks. Appl. Environ. Microbiol. 2017, 83, e00609-17. [Google Scholar] [CrossRef]
- Moher, D.; Shamseer, L.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A.; Group, P.-P. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015 statement. Syst. Rev. 2015, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Munn, Z.; Moola, S.; Lisy, K.; Riitano, D.; Tufanaru, C. Methodological guidance for systematic reviews of observational epidemiological studies reporting prevalence and cumulative incidence data. Int. J. Evid. Based Healthc. 2015, 13, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.T.; Thompson, S.G. Quantifying heterogeneity in a meta-analysis. Stat. Med. 2002, 21, 1539–1558. [Google Scholar] [CrossRef] [PubMed]
- Begg, C.B.; Mazumdar, M. Operating characteristics of a rank correlation test for publication bias. Biometrics 1994, 50, 1088–1101. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).