
Alterations of Fucosyltransferase Genes and Fucosylated Glycans in Gastric Epithelial Cells Infected with Helicobacter pylori

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Cell Lines and Cell Culture
2.3. Infection Procedures
2.4. RNA Extraction and Quantitative RT-PCR
2.5. Cellular Lysis
2.6. Lectin Microarrays and Data Analysis
2.7. Statistical Analyses
3. Results
3.1. Glycopatterns Changed after Interaction of Gastric Epithelial Cells with H. pylori Strains
3.2. Glycopatterns in GES-1 Cells Infected by H. pylori Isolates from Different Sources
3.3. Time Course of Relative Expression of FUTs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shan, M.; Yang, D.; Dou, H.; Zhang, L. Fucosylation in cancer biology and its clinical applications. Progress Mol. Biol. Transl. Sci. 2019. [Google Scholar] [CrossRef]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef] [PubMed]
- Blanas, A.; Sahasrabudhe, N.M.; Rodríguez, E.; Van Kooyk, Y.; Van Vliet, S.J. Fucosylated Antigens in Cancer: An Alliance toward Tumor Progression, Metastasis, and Resistance to Chemotherapy. Front. Oncol. 2018, 8, 39. [Google Scholar] [CrossRef]
- Moriwaki, K.; Miyoshi, E. Fucosylation and gastrointestinal cancer. World J. Hepatol. 2010, 2, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, E.; Moriwaki, K.; Terao, N.; Tan, C.-C.; Terao, M.; Nakagawa, T.; Matsumoto, H.; Shinzaki, S.; Kamada, Y. Fucosylation Is a Promising Target for Cancer Diagnosis and Therapy. Biomolecules 2012, 2, 34–45. [Google Scholar] [CrossRef]
- Holst, S.; Wuhrer, M.; Rombouts, Y. Glycosylation Characteristics of Colorectal Cancer. Adv. Cancer Res. 2015, 126, 203–256. [Google Scholar] [CrossRef]
- Holst, S.; Wilding, J.; Koprowska, L.; Rombouts, K.; Wuhrer, Y.; Manfred, Y. N-Glycomic and Transcriptomic Changes Associated with CDX1 mRNA Expression in Colorectal Cancer Cell Lines. Cells 2019, 8, 273. [Google Scholar] [CrossRef]
- Breiman, A.; Robles, M.D.L.; Trécesson, S.D.C.; Echasserieau, K.; Bernardeau, K.; Drickamer, K.; Imberty, A.; Barillé-Nion, S.; Altare, F.; Le Pendu, J. Carcinoma-associated fucosylated antigens are markers of the epithelial state and can contribute to cell adhesion through CLEC17A (Prolectin). Oncotarget 2016, 7, 14064–14082. [Google Scholar] [CrossRef]
- Gagneux, P.; Cheriyan, M.; Hurtado-Ziola, N.; Van Der Linden, E.C.M.B.; Anderson, D.C.; McClure, H.M.; Varki, A.; Varki, N. Human-specific Regulation of α2–6-linked Sialic Acids. J. Biol. Chem. 2003, 278, 48245–48250. [Google Scholar] [CrossRef]
- Sugano, K.; Tack, J.; Kuipers, E.J.; Graham, D.Y.; El-Omar, E.M.; Miura, S.; Haruma, K.; Asaka, M.; Uemura, N.; Malfertheiner, P. Kyoto global consensus report on Helicobacter pylori gastritis. Gut 2015, 64, 1353–1367. [Google Scholar] [CrossRef]
- Crowe, S.E. Helicobacter pylori Infection. N. Engl. J. Med. 2019, 380, 1158–1165. [Google Scholar] [CrossRef] [PubMed]
- Cairns, M.; Gupta, A.; A Naughton, J.; Kane, M.; Clyne, M.; Joshi, L. Glycosylation-related gene expression in HT29-MTX-E12 cells upon infection by Helicobacter pylori. World J. Gastroenterol. 2017, 23, 6817–6832. [Google Scholar] [CrossRef]
- Magalhães, A.; Reis, C.A. Helicobacter pylori adhesion to gastric epithelial cells is mediated by glycan receptors. Braz. J. Med. Biol. Res. 2010, 43, 611–618. [Google Scholar]
- Stowell, C.P.; Stowell, S.R. Biologic roles of the ABH and Lewis histo-blood group antigens Part I: Infection and immunity. Vox Sang. 2019, 114, 426–442. [Google Scholar] [CrossRef] [PubMed]
- Bugaytsova, J.A.; Björnham, O.; Chernov, Y.A.; Gideonsson, P.; Henriksson, S.; Mendez, M.; Sjöström, R.; Mahdavi, J.; Shevtsova, A.; Ilver, D.; et al. Helicobacter pylori Adapts to Chronic Infection and Gastric Disease via pH-Responsive BabA-Mediated Adherence. Cell Host Microbe 2017, 21, 376–389. [Google Scholar] [CrossRef]
- Ilver, D.; Arnqvist, A.; Ögren, J.; Frick, I.-M.; Kersulyte, D.; Incecik, E.T.; Berg, D.E.; Covacci, A.; Engstrand, L.; Borén, T. Helicobacter pylori Adhesin Binding Fucosylated Histo-Blood Group Antigens Revealed by Retagging. Science 1998, 279, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Oleastro, M.; Ménard, A. The Role of Helicobacter pylori Outer Membrane Proteins in Adherence and Pathogenesis. Biology 2013, 2, 1110–1134. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi, J.; Sondén, B.; Hurtig, M.; Olfat, F.O.; Forsberg, L.; Roche, N.; Ångström, J.; Larsson, T.; Teneberg, S.; Karlsson, K.-A.; et al. Helicobacter pylori SabA Adhesin in Persistent Infection and Chronic Inflammation. Science 2002, 297, 573–578. [Google Scholar] [CrossRef]
- Marcos, N.; Magalhães, A.; Ferreira, B.; Oliveira, M.J.; Carvalho, A.S.; Mendes, N.; Gilmartin, T.; Head, S.R.; Figueiredo, C.; David, L.; et al. Helicobacter pylori induces β3GnT5 in human gastric cell lines, modulating expression of the SabA ligand sialyl–Lewis x. J. Clin. Investig. 2008, 118, 2325–2336. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, A.; Rossez, Y.; Robbe-Masselot, C.; Maes, E.; Gomes, J.; Shevtsova, A.; Bugaytsova, J.; Borén, T.; Reis, C.A. Muc5ac gastric mucin glycosylation is shaped by FUT2 activity and functionally impacts Helicobacter pylori binding. Sci. Rep. 2016, 6, 25575. [Google Scholar] [CrossRef]
- Flach, C.-F.; Svensson, N.; Blomquist, M.; Ekman, A.; Raghavan, S.; Holmgren, J. A truncated form of HpaA is a promising antigen for use in a vaccine against Helicobacter pylori. Vaccine 2011, 29, 1235–1241. [Google Scholar] [CrossRef]
- Roy, B.; Chattopadhyay, G.; Mishra, D.; Das, T.; Chakraborty, S.; Maiti, T.K. On-chip lectin microarray for glycoprofiling of different gastritis types and gastric cancer. Biomicrofluidics 2014, 8, 34107. [Google Scholar] [CrossRef]
- Yang, L.; Yang, Z.; Cheng, L.; Cheng, J.; Cheng, L.; Sun, Y.; Li, W.; Song, K.; Huang, W.; Yin, Y. Lectin Microarray Combined with Mass Spectrometry Identifies Haptoglobin-Related Protein (Hpr) as a Potential Serologic Biomarker for Separating Non-Bacterial Pneumonia from Bacterial Pneumonia in Childhood. Proteom. Clin. Appl. 2018, 1800030. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.-C.; Li, Y.; Zhou, J.; Qian, J.; Schnaar, R.L.; Zhang, Y.; Goldstein, I.J.; Zhu, H.; Schneck, J.P. Lectin microarrays identify cell-specific and functionally significant cell surface glycan markers. Glycobiology 2008, 18, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Hsu, K.-L.; Pilobello, K.; Krishnamoorthy, L.; Mahal, L.K. Ratiometric Lectin Microarray Analysis of the Mammalian Cell Surface Glycome. Adv. Struct. Saf. Stud. 2010, 671, 117–131. [Google Scholar] [CrossRef]
- Chen, Y.; Ding, L.; Ju, H. In Situ Cellular Glycan Analysis. Accounts Chem. Res. 2018, 51, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, K.; Kuno, A.; Matsuda, A.; Ikehata, Y.; Katada, N.; Hirabayashi, J.; Narimatsu, H.; Watanabe, M. Lectin microarray technology identifies specific lectins related to lymph node metastasis of advanced gastric cancer. Gastric Cancer 2015, 19, 531–542. [Google Scholar] [CrossRef]
- Nolan, T.; E Hands, R.; A Bustin, S. Quantification of mRNA using real-time RT-PCR. Nat. Protoc. 2006, 1, 1559–1582. [Google Scholar] [CrossRef] [PubMed]
- Nagura-Ikeda, M.; Imai, K.; Tabata, S.; Miyoshi, K.; Murahara, N.; Mizuno, T.; Horiuchi, M.; Kato, K.; Imoto, Y.; Iwata, M.; et al. Clinical Evaluation of Self-Collected Saliva by Quantitative Reverse Transcription-PCR (RT-qPCR), Direct RT-qPCR, Reverse Transcription–Loop-Mediated Isothermal Amplification, and a Rapid Antigen Test To Diagnose COVID-19. J. Clin. Microbiol. 2020, 58. [Google Scholar] [CrossRef]
- Cooke, C.L.; An, H.J.; Kim, J.; Canfield, D.R.; Torres, J.; Lebrilla, C.B.; Solnick, J.V. Modification of Gastric Mucin Oligosaccharide Expression in Rhesus Macaques After Infection With Helicobacter pylori. Gastroenterology 2009, 137, 1061–1071. [Google Scholar] [CrossRef]
- Takeshi, O.; Hironori, Y.; Teruko, N.; In-Dal, P.; Myung-Woong, C.; Hideo, Y.; Kiwamu, O.; Mutsunori, S. A change in PBP1 is involved in amoxicillin resistance of clinical isolates of Helicobacter pylori. J. Antimicrob. Chemother. 2002, 50, 849–856. [Google Scholar]
- Nyström, K.; Biller, M.; Grahn, A.; Lindh, M.; Larson, G.; Olofsson, S. Real time PCR for monitoring regulation of host gene expression in herpes simplex virus type 1-infected human diploid cells. J. Virol. Methods 2004, 118, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Nyström, K.; Grahn, A.; Lindh, M.; Brytting, M.; Mandel, U.; Larson, G.; Olofsson, S. Virus-induced transcriptional activation of host FUT genes associated with neo-expression of Ley in cytomegalovirus-infected and sialyl-Lex in varicella-zoster virus-infected diploid human cells. Glycobiology 2007, 17, 355–366. [Google Scholar] [CrossRef]
- Ying, H.; Qi-Long, W.; Dan-Dan, C.; Wen-Ting, X.; Nong-Hua, L. Adhesion and Invasion of Gastric Mucosa Epithelial Cells by Helicobacter pylori. Front. Cell. Infect. Microbiol. 2016, 6, 159. [Google Scholar]
- Dan, X.; Liu, W.; Ng, T.B. Development and Applications of Lectins as Biological Tools in Biomedical Research. Med. Res. Rev. 2015, 36, 221–247. [Google Scholar] [CrossRef]
- Aspholm-Hurtig, M.; Dailide, G.; Lahmann, M.; Kalia, A.; Ilver, D.; Roche, N.; Vikström, S.; Sjöström, R.; Lindén, S.; Bäckström, A.; et al. Functional Adaptation of BabA, the H. pylori ABO Blood Group Antigen Binding Adhesin. Science 2004, 305, 519–522. [Google Scholar] [CrossRef] [PubMed]
- McGuckin, M.A.; Lindén, S.K.; Sutton, P.; Florin, T.H. Mucin dynamics and enteric pathogens. Nat. Rev. Genet. 2011, 9, 265–278. [Google Scholar] [CrossRef]
- Ansari, S.; Yamaoka, Y. Helicobacter pylori BabA in adaptation for gastric colonization. World J. Gastroenterol. 2017, 4158–4169. [Google Scholar] [CrossRef]
- Grahn, E.M.; Winter, H.C.; Tateno, H.; Goldstein, I.J.; Krengel, U. Structural Characterization of a Lectin from the Mushroom Marasmius oreades in Complex with the Blood Group B Trisaccharide and Calcium. J. Mol. Biol. 2009, 390, 457–466. [Google Scholar] [CrossRef]
- Saboti, J.; Kos, J. CNL–Clitocybe nebularis Lectin—The Fungal GalNAcβ1-4GlcNAc-Binding Lectin. Molecules 2019, 24, 4204. [Google Scholar] [CrossRef]
- Padró, M.; Cobler, L.; Garrido, M.; De Bolós, C. Down-regulation of FUT3 and FUT5 by shRNA alters Lewis antigens expression and reduces the adhesion capacities of gastric cancer cells. Biochim. Biophys. Acta BBA Gen. Subj. 2011, 1810, 1141–1149. [Google Scholar] [CrossRef]
- Cheng, L.; Jin, M.; Zhou, J. FUT family mediates the multidrug resistance of human hepatocellular carcinoma via the PI3K/Akt signaling pathway. Cell Death Disease 2013. [Google Scholar] [CrossRef]
- Magalhães, A.; Gomes, J.; Ismail, M.N.; Haslam, S.M.; Mendes, N.; Osório, H.; David, L.; Le Pendu, J.; Haas, R.; Dell, A.; et al. Fut2-null mice display an altered glycosylation profile and impaired BabA-mediated Helicobacter pylori adhesion to gastric mucosa. Glycobiology 2009, 19, 1525–1536. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Uematsu, S.; Kiyono, H. Epithelial glycosylation in gut homeostasis and inflammation. Nat. Immunol. 2016, 17, 1244–1251. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, Y. Increasing evidence of the role of Helicobacter pylori SabA in the pathogenesis of gastroduodenal disease. J. Infect. Dev. Ctries. 2008, 2, 174–181. [Google Scholar] [CrossRef]
- Lopez-Ferrer, A.; de Bolosa, C.; Barrancob, C.; Garridoa, M.; Iserna, J.; Carlstedtc, I.; Reisd, C.A.; Torradoe, J.; Reala, F.X. Role of fucosyltransferases in the association between apomucin and Lewis antigen expression in normal and malignant gastric epithelium. Gut 2000, 47, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Theodora, d.V.; Holmes, E.H.; Macher, B.A. Fucosyltransferases: Structure/function studies. Glycobiology 2001, 11, 119R. [Google Scholar]
- You, Y.; He, L.; Zhang, M.; Fu, J.; Gu, Y.; Zhang, B.; Tao, X.; Zhang, J. Comparative Genomics of Helicobacter pylori Strains of China Associated with Different Clinical Outcome. PLoS ONE 2012, 7, e38528. [Google Scholar] [CrossRef] [PubMed]

 ), P164 (
), P164 ( ), HLJ011 (
), HLJ011 ( ), HLJ030 (
), HLJ030 ( ), M84 (
), M84 ( ), and YN4-62 (
), and YN4-62 ( ) at 0.5–8 h was analyzed using real-time PCR. The expression quantity of FUT genes in uninfected cells was assigned to 1, and the expression level of FUT genes was considered up-regulated in infected cells when the relative quantity > 1.
), P164 (), HLJ011 (), HLJ030 (), M84 (), and YN4-62 () at 0.5–8 h was analyzed using real-time PCR. The expression quantity of FUT genes in uninfected cells was assigned to 1, and the expression level of FUT genes was considered up-regulated in infected cells when the relative quantity > 1.
) at 0.5–8 h was analyzed using real-time PCR. The expression quantity of FUT genes in uninfected cells was assigned to 1, and the expression level of FUT genes was considered up-regulated in infected cells when the relative quantity > 1.
), P164 (), HLJ011 (), HLJ030 (), M84 (), and YN4-62 () at 0.5–8 h was analyzed using real-time PCR. The expression quantity of FUT genes in uninfected cells was assigned to 1, and the expression level of FUT genes was considered up-regulated in infected cells when the relative quantity > 1.

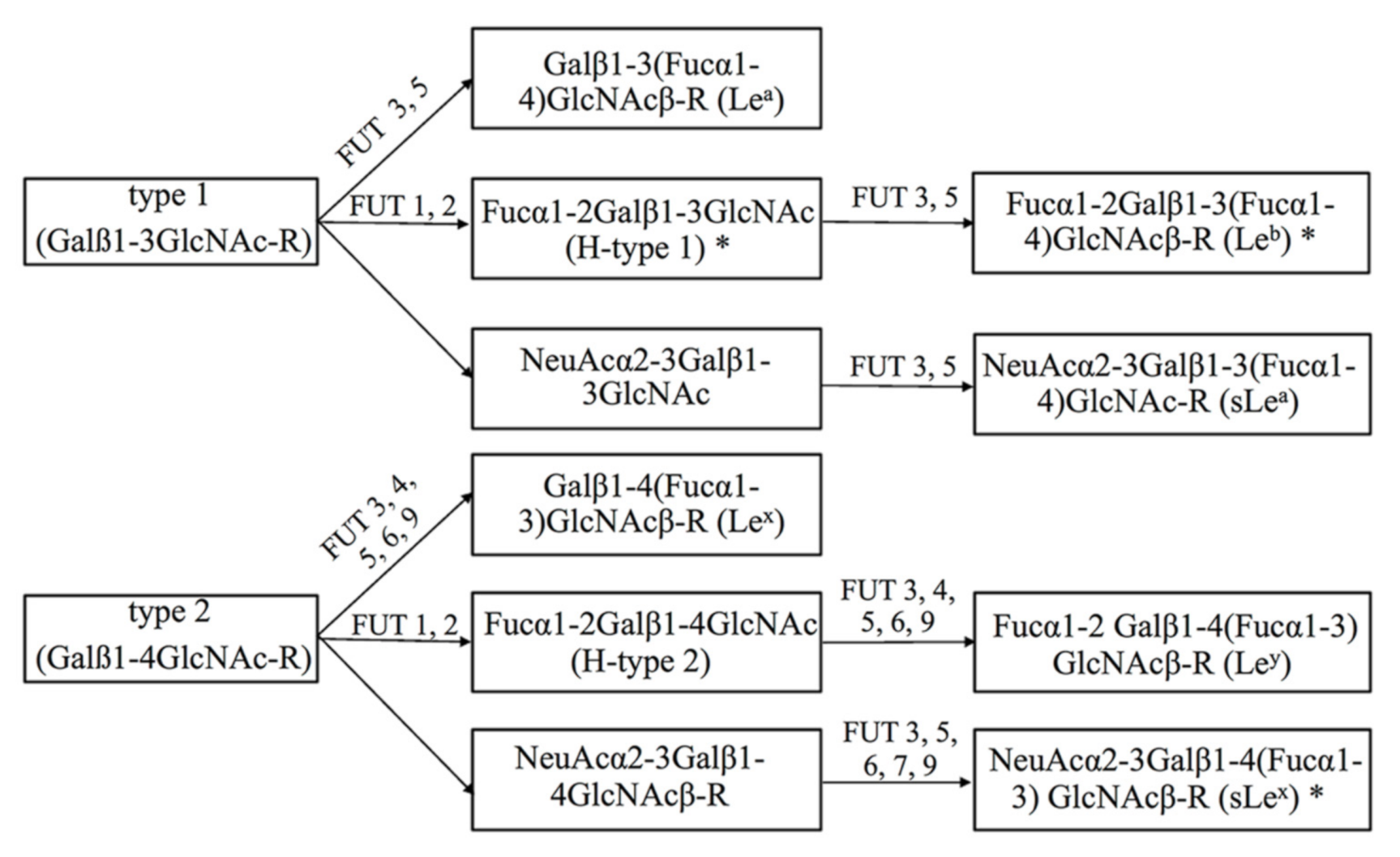{kind=link}
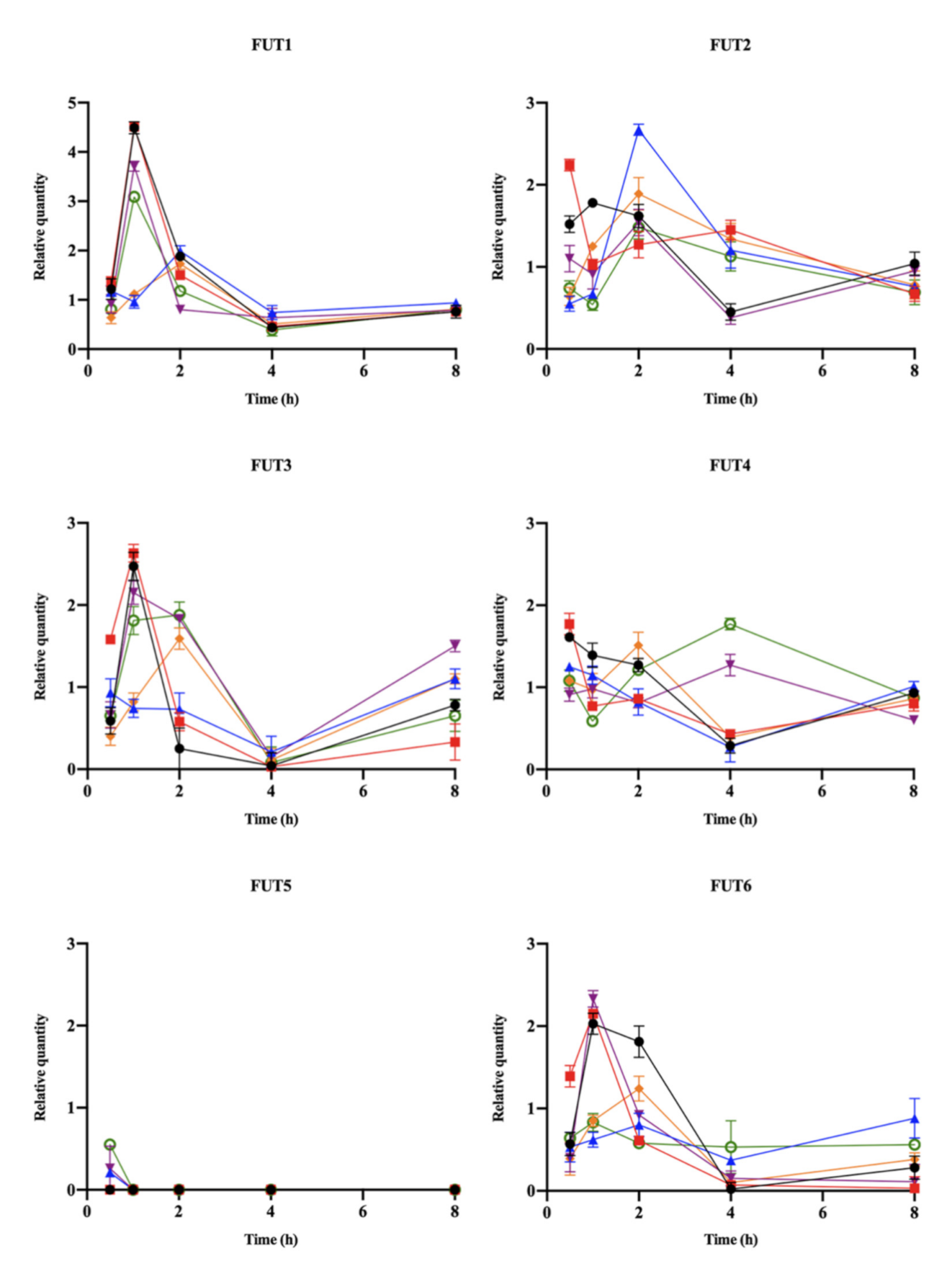{kind=link}
| Gene | Forward Primer | Reverse Primer | Probe a |
|---|---|---|---|
| FUT1 | AGGTATAAACACACCCTCTGTGCTT | GAGTTCAGGGACAGACAGTGGTT | AAACTGGCAGGTACCGTGCTCATTGC |
| FUT2 | CTCGCTACAGCTCCCTCATCTT | CGTGGGAGGTGTCAATGTTCT | TGGTCACCAGTAATGGCATGGCCTG |
| FUT3 | GGGATCCCTTTTCGTCACACT | CGAACTGGTCTAAGCCTTGCA | AGGTGACCTACAGGCTCCGCTCGA |
| FUT4 | AATTGGGCTCCTGCACAC | CCAGGTGCTGCGAGTTCT | TGGCCCGCTACAAGTTCTACCTGG |
| FUT5 | CGCTGGATCTGGTTCAGCAT | CAGCCGTAGGGCGTGAG | CCCCCAGCAACTCCGGC |
| FUT6 | GCATCCAGACGGGATCCA | ACTGCTGCGTCTTGACACCTT | CCAGGTCCCCGATCCCTCTAGCAT |
| GAPDH | GAGAAGGCTGGGGCTCAT | TGCTGATGATCTTGAGGCTG | CTCTGCTGATGCCCCCATGTTCGT |
| Lectin | Carbohydrate Specificity | ATCC 43504-GES-1 | P164-GES-1 | M84-GES-1 | YN4-62-GES-1 | HLJ011-GES-1 | HLJ030-GES-1 |
|---|---|---|---|---|---|---|---|
| Lotus | αFuc | 1.0 | 1.7 | 1.6 | 1.1 | 1.4 | 1.0 |
| RS-Fuc | αFuc | 0.5 | 4.2 | 5.1 | 3.3 | 5.8 | 0.9 |
| UEA-I | αFuc | 0.9 | 1.1 | 1.2 | 1.2 | 1.2 | 0.9 |
| PA-IIL b | Fuc, Man | 1.0 | 1.5 | 1.6 | 1.1 | 2.2 | 1.0 |
| AAA | Fucα1-6GlcNAc, Fucα1-3GlcNAc | 0.8 | 1.8 | 1.2 | 2.0 | 1.2 | 1.0 |
| AAL | Fucα1-6GlcNAc, Fucα1-3LacNAc | 0.6 | 1.2 | 1.2 | 1.8 | 1.3 | 1.0 |
| BC2LCN | Fucα1-2Galβ1-3GalNAc, Fucα1-2Galβ1-3GlcNAc | 1.0 | 1.8 | 1.9 | 1.1 | 2.0 | 1.0 |
| CNL | GalNAcβ1-4GlcNAc, GalNAcα1-3(Fucα1-2)Galβ1-4GlcNAc | 1.0 | 1.4 | 1.4 | 1.1 | 1.4 | 1.0 |
| MOA | Galα1-3Galβ1-4GlcNAc, Galα1-3(Fucα1-2)Gal | 0.6 | 1.9 | 1.8 | 2.9 | 2.2 | 0.8 |
| ACG | α2-3 Sia | 0.9 | 2.9 | 2.2 | 2.8 | 3.2 | 0.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, R.; Han, X.; Gong, Y.; He, L.; Xue, Z.; Yang, Y.; Sun, L.; Fan, D.; You, Y.; Meng, F.; et al. Alterations of Fucosyltransferase Genes and Fucosylated Glycans in Gastric Epithelial Cells Infected with Helicobacter pylori. Pathogens 2021, 10, 168. https://doi.org/10.3390/pathogens10020168
Fan R, Han X, Gong Y, He L, Xue Z, Yang Y, Sun L, Fan D, You Y, Meng F, et al. Alterations of Fucosyltransferase Genes and Fucosylated Glycans in Gastric Epithelial Cells Infected with Helicobacter pylori. Pathogens. 2021; 10(2):168. https://doi.org/10.3390/pathogens10020168
Chicago/Turabian StyleFan, Ruyue, Xiurui Han, Yanan Gong, Lihua He, Zhijing Xue, Yaming Yang, Lu Sun, Dongjie Fan, Yuanhai You, Fanliang Meng, and et al. 2021. "Alterations of Fucosyltransferase Genes and Fucosylated Glycans in Gastric Epithelial Cells Infected with Helicobacter pylori" Pathogens 10, no. 2: 168. https://doi.org/10.3390/pathogens10020168
APA StyleFan, R., Han, X., Gong, Y., He, L., Xue, Z., Yang, Y., Sun, L., Fan, D., You, Y., Meng, F., Yan, X., Zhang, M., & Zhang, J. (2021). Alterations of Fucosyltransferase Genes and Fucosylated Glycans in Gastric Epithelial Cells Infected with Helicobacter pylori. Pathogens, 10(2), 168. https://doi.org/10.3390/pathogens10020168