
The Ecology and Evolution of Japanese Encephalitis Virus

 , , , , and
, , , , and 
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Classification, Virion and Genome Structure
3. Epidemiology of Japanese Encephalitis
4. Clinical Picture
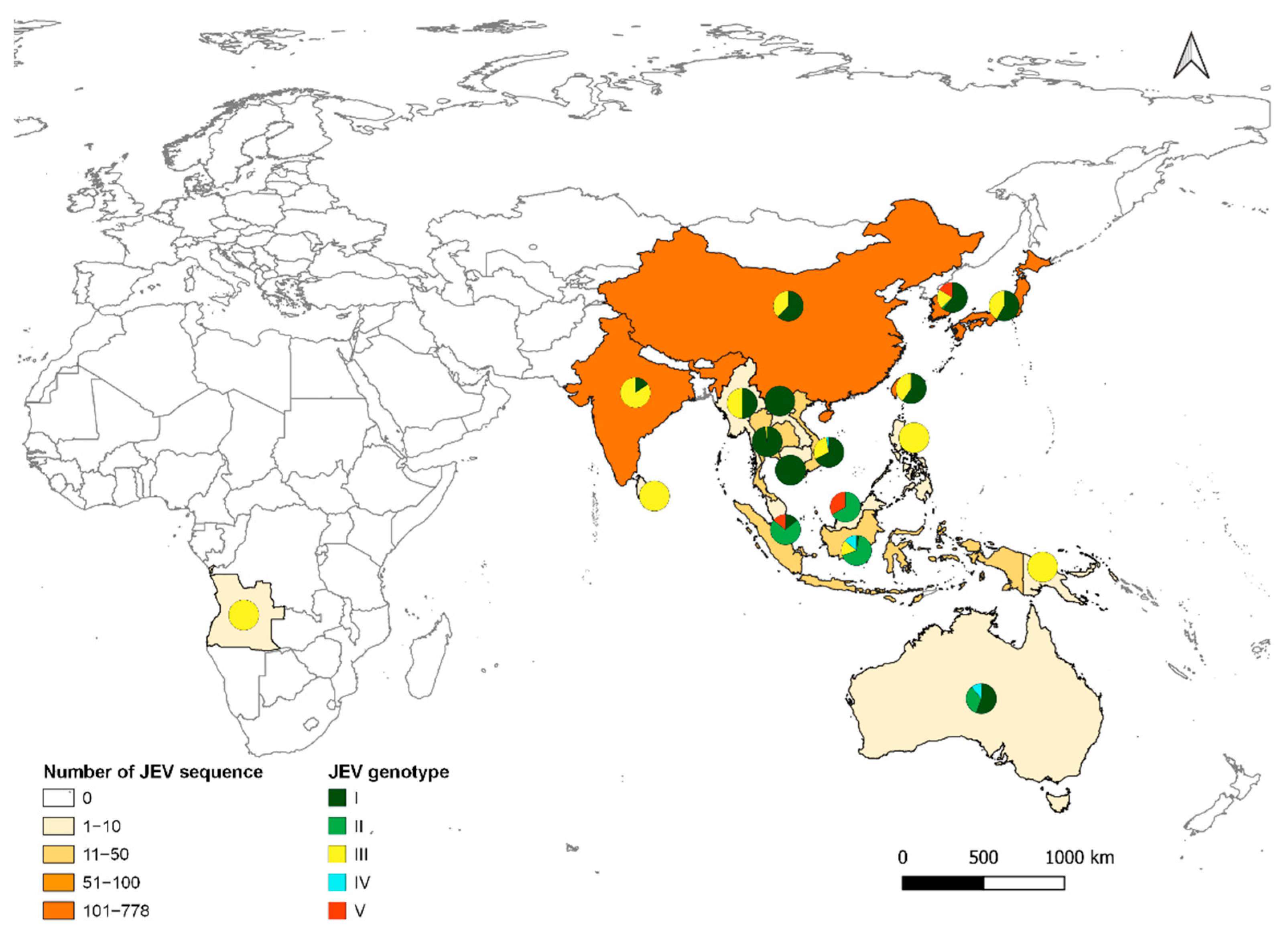
5. Genetic Evolution of JEV
6. JEV and Asian Economic Development
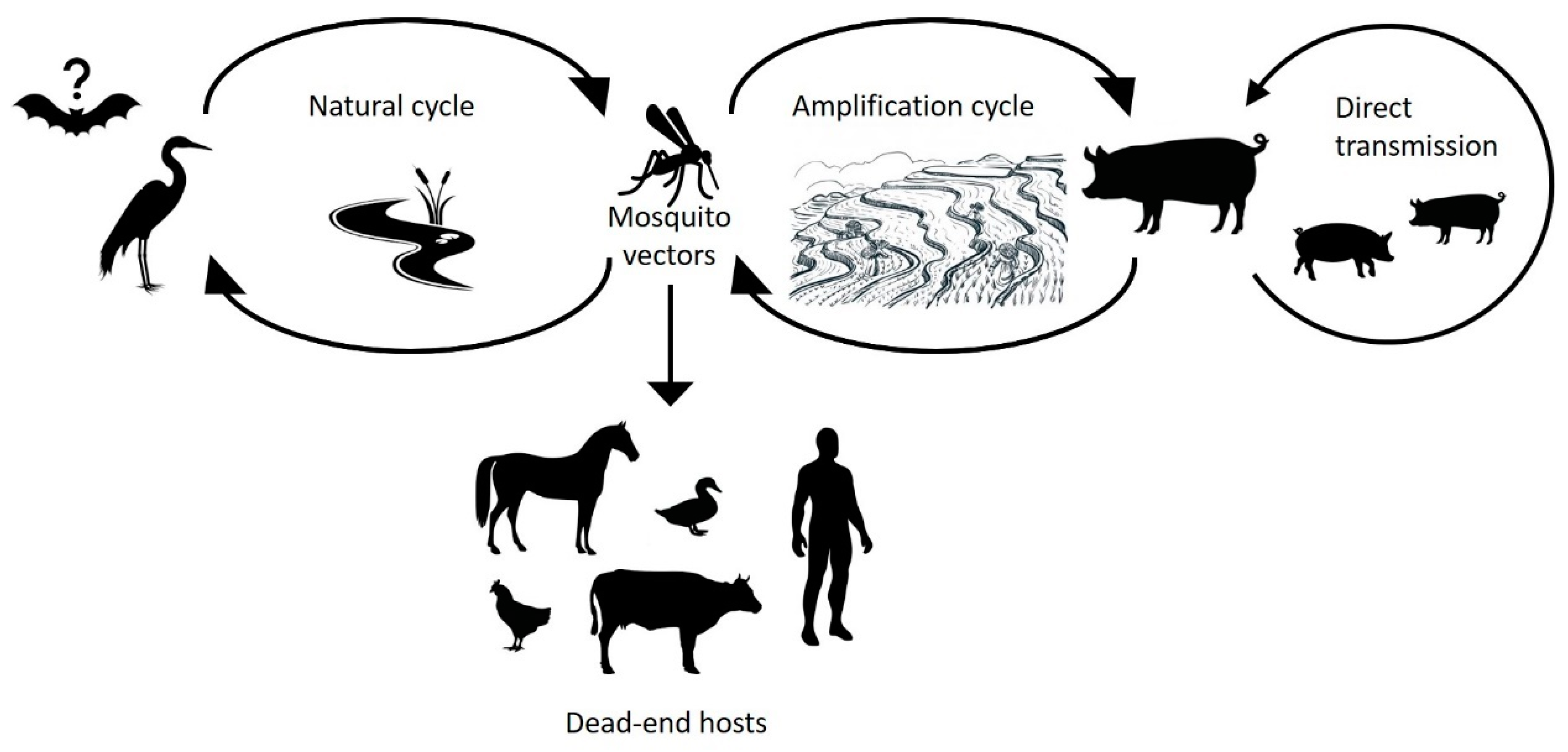
7. The Ecology of JEV
7.1. Mosquitoes
7.2. Birds
7.3. Pigs
7.4. Other Hosts
8. Possible Impacts of Climate Change
9. JEV Vaccination
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Campbell, G.; Hills, S.; Fischer, M.; Jacobson, J.; Hoke, C.; Hombach, J.; Marfin, A.; Solomon, T.; Tsai, T.; Tsu, V.; et al. Estimated global incidence of Japanese encephalitis: A systematic review. Bull. World Health Organ. 2011, 89, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Solomon, T.; Dung, N.M.; Kneen, R.; Gainsborough, M.; Vaughn, D.W.; Khanh, V.T. Japanese encephalitis. J. Neurol. Neurosurg. Psychiatry 2000, 68, 405–415. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Japanese Encephalitis. 2019. Available online: https://www.who.int/news-room/fact-sheets/detail/japanese-encephalitis (accessed on 1 August 2021).
- Tarantola, A.; Goutard, F.; Newton, P.; De Lamballerie, X.; Lortholary, O.; Cappelle, J.; Buchy, P. Estimating the burden of Japanese encephalitis virus and other encephalitides in countries of the Mekong region. PLoS Negl. Trop. Dis. 2014, 8, e2533. [Google Scholar] [CrossRef]
- Quan, T.M.; Thao, T.T.; Duy, N.M.; Nhat, T.M.; Clapham, H. Estimates of the global burden of Japanese encephalitis and the impact of vaccination from 2000–2015. eLife 2020, 9, e51027. [Google Scholar] [CrossRef] [PubMed]
- Lord, J.S.; Gurley, E.S.; Pulliam, J.R. Rethinking Japanese encephalitis virus transmission: A framework for implicating host and vector species. PLoS Negl. Trop. Dis. 2015, 9, e0004074. [Google Scholar] [CrossRef] [PubMed]
- Konno, J.; Endo, K.; Agatsuma, H.; Ishida, N. Cyclic outbreaks of Japanese encephalitis among pigs and humans. Am. J. Epidemiol. 1966, 84, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Dhanda, V.; Banerjee, K.; Deshmukh, P.; IIkal, M. Experimental viraemia and transmission of Japanese encephalitis virus by mosquitoes in domestic ducks. Indian J. Med. Res. 1977, 66, 881–888. [Google Scholar] [PubMed]
- Soman, R.; Rodrigues, F.; Guttikar, S.; Guru, P. Experimental viraemia and transmission of Japanese encephalitis virus by mosquitoes in ardeid birds. Indian J. Med. Res. 1977, 66, 709–718. [Google Scholar]
- Gresser, I.; Hardy, J.L.; Hu, S.M.; Scherer, W.F. Factors influencing transmission of Japanese B encephalitis virus by a colonized strain of Culex tritaeniorhynchus Giles, from infected pigs and chicks to susceptible pigs and birds. Am. J. Trop. Med. Hyg. 1958, 7, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Yap, G.; Lim, X.F.; Chan, S.; How, C.B.; Humaidi, M.; Yeo, G.; Mailepessov, D.; Kong, M.; Lai, Y.L.; Okumura, C.; et al. Serological evidence of continued Japanese encephalitis virus transmission in Singapore nearly three decades after end of pig farming. Parasites Vectors 2019, 12, 244. [Google Scholar] [CrossRef]
- Yang, S.-E.; Pan, M.-J.; Tseng, H.-F.; Liau, M.-Y. The efficacy of mouse-brain inactivated Nakayama strain Japanese encephalitis vaccine—Results from 30 years experience in Taiwan. Vaccine 2006, 24, 2669–2673. [Google Scholar] [CrossRef]
- Hanna, J.N.; Ritchie, S.A.; Hills, S.L.; van den Hurk, A.F.; Phillips, D.A.; Pyke, A.T.; Lee, J.M.; Johansen, C.A.; Mackenzie, J.S. Japanese encephalitis in north Queensland, Australia, 1998. Med. J. Aust. 1999, 170, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Wood, B.L.; Beck, L.R.; Washino, R.K.; Hibbard, K.A.; Salute, J.S. Estimating high mosquito-producing rice fields using spectral and spacial data. Int. J. Remote Sens. 1992, 13, 2813–2826. [Google Scholar] [CrossRef]
- Wood, B.L.; Beck, L.R.; Washino, R.K.; Palchick, S.M.; Sebesta, P.D. 1991. Spectral and spatial characterization of rice field mosquito habitat. Int. J. Remote Sens. 1991, 12, 621–626. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses (ICTV). Genus: Flavivirus. 2021. Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/positive-sense-rna-viruses/w/flaviviridae/360/genus-flavivirus (accessed on 1 August 2021).
- Yun, S.I.; Lee, Y.M. Japanese encephalitis: The virus and vaccines. Hum. Vaccines Immunother. 2014, 10, 263–279. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, A. Epidemiology and control of Japanese encephalitis. Health Stat. Q. 1992, 45, 299–305. [Google Scholar]
- Huynh, T.T.T.; Aarnink, A.J.A.; Drucker, A.; Verstegen, M.W.A. Pig production in Cambodia, Laos, Philippines, and Vietnam: A review. Asian J. Agric. Dev. 2006, 3, 69–90. [Google Scholar]
- Laborte, A.G.; Gutierrez, M.A.; Balanza, J.G.; Saito, K.; Zwart, S.J.; Boschetti, M.; Murty, M.V.R.; Villano, L.; Aunario, J.K.; Reinke, R.; et al. RiceAtlas, a spatial database of global rice calendars and production. Sci. Data 2017, 4, 170074. [Google Scholar] [CrossRef] [PubMed]
- Yen, N.T.; Duffy, M.R.; Hong, N.M.; Hien, N.T.; Fischer, M.; Hills, S.L. Surveillance for Japanese encephalitis in Vietnam, 1998–2007. Am. J. Trop. Med. Hyg. 2010, 83, 816. [Google Scholar] [CrossRef]
- Ho Dang Trung, N.; Le Thi Phuong, T.; Wolbers, M.; Nguyen Van Minh, H.; Nguyen Thanh, V.; Van, M.P.; Thieu, N.T.; Le Van, T.; Song, D.T.; Le Thi, P.; et al. Aetiologies of central nervous system infection in Viet Nam: A prospective provincial hospital-based descriptive surveillance study. PLoS ONE 2012, 7, e37825. [Google Scholar]
- Horwood, P.F.; Duong, V.; Laurent, D.; Mey, C.; Sothy, H.; Santy, K.; Richner, B.; Heng, S.; Hem, S.; Cheval, J.; et al. Aetiology of acute meningoencephalitis in Cambodian children, 2010–2013. Emerg. Microbes Infect. 2017, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.N.; Ritchie, S.A.; Phillips, D.A.; Shield, J.; Bailey, M.C.; Mackenzie, J.S.; Poidinger, M.; McCall, B.J.; Mills, P.J. An outbreak of Japanese encephalitis in the Torres Strait, Australia, 1995. Med. J. Aust. 1996, 165, 256–260. [Google Scholar] [CrossRef]
- Mackenzie, J.S.; Williams, D.T.; Smith, D.W. Japanese encephalitis virus: The geographic distribution, incidence, and spread of a virus with a propensity to emerge in new areas. Perspect. Med. Virol. 2006, 16, 201–268. [Google Scholar]
- Johansen, C.A.; van den Hurk, A.F.; Pyke, A.T.; Zborowski, P.; Phillips, D.A.; Mackenzie, J.S.; Ritchie, S.A. Entomological investigations of an outbreak of Japanese encephalitis virus in the Torres Strait, Australia, in 1998. J. Med. Entomol. 2001, 38, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Van den Hurk, A.F.; Nisbet, D.J.; Johansen, C.A.; Foley, P.N.; Ritchie, S.A.; Mackenzie, J.S. Japanese encephalitis on Badu Island, Australia: The first isolation of Japanese encephalitis virus from Culex gelidus in the Australasian region and the role of mosquito host-feeding patterns in virus transmission cycles. Trans. R. Soc. Trop. Med. Hyg. 2001, 95, 595–600. [Google Scholar] [CrossRef]
- Van den Hurk, A.F.; Ritchie, S.A.; Johansen, C.A.; Mackenzie, J.S.; Smith, G.A. Domestic pigs and Japanese encephalitis virus infection, Australia. Emerg. Infect. Dis. 2008, 14, 1736. [Google Scholar] [CrossRef] [PubMed]
- Johansen, C.A.; Nisbet, D.J.; Foley, P.N.; van Den Hurk, A.F.; Hall, R.A.; Mackenzie, J.S.; Ritchie, S.A. Flavivirus isolations from mosquitoes collected from Saibai Island in the Torres Strait, Australia, during an incursion of Japanese encephalitis virus. Med. Vet. Entomol. 2004, 18, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Johansen, C.A.; van den Hurk, A.F.; Ritchie, S.A.; Zborowski, P.; Nisbet, D.J.; Paru, R.A.; Bockarie, M.J.; Macdonald, J.; Drew, A.C.; Khromykh, T.I.; et al. Isolation of Japanese encephalitis virus from mosquitoes (Diptera: Culicidae) collected in the Western Province of Papua New Guinea, 1997–1998. Am. J. Trop. Med. Hyg. 2000, 62, 631–638. [Google Scholar] [CrossRef]
- Waller, C.; (Royal Darwin Hospital [RDH], Tiwi, Australia); Currie, B.J.; (RDH, Charles Darwin University, Casuarina, Australia); Baird, R.W.; (RDH, Tiwi, Australia); Van Beek, R.; (RDH, Tiwi, Australia); Krause, V.L.; (Northern Territory Department of Health, Darwin City, Australia). Personal communication, 2021.
- Simon-Loriere, E.; Faye, O.; Prot, M.; Casademont, I.; Fall, G.; Fernandez-Garcia, M.D.; Diagne, M.M.; Kipela, J.M.; Fall, I.S.; Holmes, E.C.; et al. Autochthonous Japanese encephalitis with yellow fever coinfection in Africa. N. Engl. J. Med. 2017, 376, 1483–1485. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, K.L.; Hernández-Triana, L.M.; Banyard, A.C.; Fooks, A.R.; Johnson, N. Japanese encephalitis virus infection, diagnosis and control in domestic animals. Vet. Microbiol. 2017, 201, 85–92. [Google Scholar] [CrossRef]
- Turtle, L.; Solomon, T. Japanese encephalitis—the prospects for new treatments. Nat. Rev. Neurol. 2018, 14, 298–313. [Google Scholar] [CrossRef] [PubMed]
- Amicizia, D.; Zangrillo, F.; Lai, P.L.; Iovine, M.; Panatto, D. Overview of Japanese encephalitis disease and its prevention. Focus on IC51 vaccine (IXIARO®). J. Prev. Med. Hyg. 2018, 59, E99. [Google Scholar] [PubMed]
- Vaughn, D.W.; Hoke, C.H., Jr. The epidemiology of Japanese encephalitis: Prospects for prevention. Epidemiol. Rev. 1992, 14, 197–221. [Google Scholar] [CrossRef]
- Ghosh, D.; Basu, A. Japanese encephalitis—A pathological and clinical perspective. PLoS Negl. Trop. Dis. 2009, 3, e437. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.J.; Gurley, E.S.; Montgomery, S.; Petersen, L.; Sejvar, J.; Fischer, M.; Panellam, A.; Powers, A.M.; Nahar, N.; Uddin, A.R.; et al. Hospital-based surveillance for Japanese encephalitis at four sites in Bangladesh, 2003–2005. Am. J. Trop. Med. Hyg. 2010, 82, 344. [Google Scholar] [CrossRef]
- Feng, Y.; Fu, S.; Zhang, H.; Petersen, L.R.; Zhang, B.; Gao, X.; Yang, W.; Zhang, Y.; Dao, B.; Li, K.; et al. High incidence of Japanese encephalitis, southern China. Emerg. Infect. Dis. 2013, 19, 672. [Google Scholar] [CrossRef]
- Solomon, T.; Ni, H.; Beasley, D.W.; Ekkelenkamp, M.; Cardosa, M.J.; Barrett, A.D. Origin and evolution of Japanese encephalitis virus in southeast Asia. J. Virol. 2003, 77, 3091–3098. [Google Scholar] [CrossRef]
- Schuh, A.J.; Guzman, H.; Tesh, R.B.; Barrett, A.D. Genetic diversity of Japanese encephalitis virus isolates obtained from the Indonesian archipelago between 1974 and 1987. Vector Borne Zoonotic Dis. 2013, 13, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Adams, J.; Chen, P.; Guo, Z.Y.; Zhong, X.F.; Fang, W.; Li, N.; Wen, L.; Tao, X.Y.; Yuan, Z.M.; et al. Comparison of genotypes I and III in Japanese encephalitis virus reveals distinct differences in their genetic and host diversity. J. Virol. 2014, 88, 11469–11479. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Liu, H.; Li, M.; Fu, S.; Liang, G. Insights into the evolutionary history of Japanese encephalitis virus (JEV) based on whole-genome sequences comprising the five genotypes. Virol. J. 2015, 12, 43. [Google Scholar] [CrossRef]
- Ma, S.P.; Yoshida, Y.; Makino, Y.; Tadano, M.; Ono, T.; Ogawa, M. Short report: A major genotype of Japanese encephalitis virus currently circulating in Japan. Am. J. Trop. Med. Hyg. 2003, 69, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.H.; Chung, Y.J.; Ban, S.J.; Kim, E.J.; Park, Y.K.; Cho, H.W. Envelope gene sequence variation among Japanese encephalitis viruses isolated in Korea. Acta Virol. 1996, 40, 303–309. [Google Scholar] [PubMed]
- Fulmali, P.V.; Sapkal, G.N.; Athawale, S.; Gore, M.M.; Mishra, A.C.; Bondre, V.P. Introduction of Japanese encephalitis virus genotype I., India. Emerg. Infect. Dis. 2011, 17, 319–321. [Google Scholar] [CrossRef]
- Wang, H.Y.; Takasaki, T.; Fu, S.H.; Sun, X.H.; Zhang, H.L.; Wang, Z.X.; Hao, Z.Y.; Zhang, J.K.; Tang, Q.; Kotaki, A.; et al. Molecular epidemiological analysis of Japanese encephalitis virus in China. J. Gen. Virol. 2007, 88, 885–894. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Fan, Y.-C.; Tu, W.-C.; Chang, R.-Y.; Shih, C.-C.; Lu, I.-H.; Chien, M.-S.; Lee, W.-C.; Chen, T.-H.; Chang, G.-J.; et al. Japanese encephalitis virus genotype replacement, Taiwan, 2009–2010. Emerg. Infect. Dis. 2011, 17, 2354–2356. [Google Scholar] [CrossRef]
- Nga, P.T.; Parquet, M.C.; Cuong, V.D.; Ma, S.-P.; Hasebe, F.; Inoue, S.; Makino, Y.; Takagi, M.; Nam, V.S.; Morita, K. Shift in Japanese encephalitis virus (JEV) genotype circulating in northern Vietnam: Implications for frequent introductions of JEV from Southeast Asia to East Asia. J. Gen. Virol. 2004, 85, 1625–1631. [Google Scholar] [CrossRef] [PubMed]
- Nitatpattana, N.; Dubot-Pérès, A.; Gouilh, M.A.; Souris, M.; Barbazan, P.; Yoksan, S.; De Lamballerie, X.; Gonzalez, J.P. Change in Japanese encephalitis virus distribution, Thailand. Emerg. Infect. Dis. 2008, 14, 1762. [Google Scholar] [CrossRef] [PubMed]
- Tsuchie, H.; Oda, K.; Vythilingam, I.; Thayan, R.; Vijayamalar, B.; Sinniah, M.; Singh, J.; Wada, T.; Tanaka, H.; Kurimura, T.; et al. Genotypes of Japanese encephalitis virus isolated in three states in Malaysia. Am. J. Trop. Med. Hyg. 1997, 56, 153–158. [Google Scholar] [CrossRef][Green Version]
- Duong, V.; Choeung, R.; Gorman, C.; Laurent, D.; Crabol, Y.; Mey, C.; Peng, B.; Di Francesco, J.; Hul, V.; Sothy, H.; et al. Isolation and full genome sequence of Japanese encephalitis virus genotype I strains from Cambodian human patients, mosquitoes and pigs. J. Gen. Virol. 2017, 98, 2287–2296. [Google Scholar] [CrossRef] [PubMed]
- Pyke, A.T.; Williams, D.T.; Nisbet, D.J.; Van Den Hurk, A.F.; Taylor, C.T.; Johansen, C.A.; Macdonald, J.; Hall, R.A.; Simmons, R.J.; Mason, R.J.; et al. The appearance of a second genotype of Japanese encephalitis virus in the Australasian region. Am. J. Trop. Med. Hyg. 2001, 65, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Schuh, A.J.; Ward, M.J.; Leigh Brown, A.J.; Barrett, A.D.T. Dynamics of the emergence and establishment of a newly dominant genotype of Japanese encephalitis virus throughout Asia. J. Virol. 2014, 88, 4522–4532. [Google Scholar] [CrossRef] [PubMed]
- Do, L.P.; Bui, T.M.; Phan, N.T. Mechanism of Japanese encephalitis virus genotypes replacement based on human, porcine and mosquito-originated cell lines model. Asian Pac. J. Trop. Med. 2016, 9, 333–336. [Google Scholar] [CrossRef] [PubMed]
- United Nations. 2012. World Urbanization Prospects, the 2011 Revision. Available online: https://www.un.org/en/development/desa/population/publications/pdf/urbanization/WUP2011_Report.pdf (accessed on 1 August 2021).
- Keiser, J.; Maltese, M.F.; Erlanger, T.E.; Bos, R.; Tanner, M.; Singer, B.H.; Utzinger, J. Effect of irrigated rice agriculture on Japanese encephalitis, including challenges and opportunities for integrated vector management. Acta Trop. 2005, 95, 40–57. [Google Scholar] [CrossRef]
- Erlanger, T.E.; Weiss, S.; Keiser, J.; Utzinger, J.; Wiedenmayer, K. Past, present, and future of Japanese encephalitis. Emerg. Infect. Dis. 2009, 15, 1–7. [Google Scholar] [CrossRef]
- Auerswald, H.; Maquart, P.O.; Chevalier, V.; Boyer, S. Mosquito vector competence for Japanese encephalitis virus. Viruses 2021, 13, 1154. [Google Scholar] [CrossRef] [PubMed]
- Simpson, D.I.H.; Smith, C.E.G.; Bowen, E.T.W.; Platt, G.S.; Way, H.; McMahon, D.; Bright, W.F.; Hill, M.N.; Mahadevan, S.; Macdonald, W.W. Arbovirus infections in Sarawak: Virus isolations from mosquitoes. Ann. Trop. Med. Parasitol. 1970, 64, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Gould, D.J.; Edelman, R.; Grossman, R.A.; Nisalak, A.; Sullivan, M.F. Study of Japanese encephalitis virus in Chiangmai Valley, Thailand IV. Vector studies 1. Am. J. Epidemiol. 1974, 100, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Longbottom, J.; Browne, A.J.; Pigott, D.M.; Sinka, M.E.; Golding, N.; Hay, S.I.; Moyes, C.L.; Shearer, F.M. Mapping the spatial distribution of the Japanese encephalitis vector, Culex tritaeniorhynchus Giles, 1901 (Diptera: Culicidae) within areas of Japanese encephalitis risk. Parasites Vectors 2017, 10, 148. [Google Scholar] [CrossRef]
- Samanidou, A.; Harbach, R. Culex (Culex) tritaeniorhynchus Giles, a newly discovered potential vector of arboviruses in Greece. Eur. Mosq. Bull. 2003, 16, 15–17. [Google Scholar]
- Gunay, F.; Alten, B.; Simsek, F.; Aldemir, A.; Linton, Y.-M. Barcoding Turkish Culex mosquitoes to facilitate arbovirus vector incrimination studies reveals hidden diversity and new potential vectors. Acta Trop. 2015, 143, 112–120. [Google Scholar] [CrossRef]
- Lessard, B.D.; Kurucz, N.; Rodriguez, J.; Carter, J.; Hardy, C.M. Detection of the Japanese encephalitis vector mosquito Culex tritaeniorhynchus in Australia using molecular diagnostics and morphology. Parasites Vectors 2021, 14, 411. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.J.; Montgomery, B.L.; Ingram, A.; Ritchie, S.A. First records of Culex gelidus from Australia. J. Am. Mosq. Control Assoc. 2001, 17, 79–80. [Google Scholar]
- Samy, A.M.; Elaagip, A.H.; Kenawy, M.A.; Ayres, C.F.; Peterson, A.T.; Soliman, D.E. Climate change influences on the global potential distribution of the mosquito Culex quinquefasciatus, vector of West Nile virus and lymphatic filariasis. PLoS ONE 2016, 11, e0163863. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Tuni, N.; Yen, N.; Nam, V.; Takagi, M. Influence of the distribution of host species on adult abundance of Japanese encephalitis vectors—Culex vishnui subgroup and Culex gelidus—in a rice-cultivating village in northern Vietnam. Am. J. Trop. Med. Hyg. 2008, 78, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Mwandawiro, C.; Boots, M.; Tuno, N.; Suwonkerd, W.; Tsuda, Y.; Takagi, M. Heterogeneity in the host preference of Japanese encephalitis vectors in Chiang Mai, northern Thailand. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 238–242. [Google Scholar] [CrossRef]
- Boyer, S.; Durand, B.; Yean, S.; Brengues, C.; Maquart, P.-O.; Fontenille, D.; Chevalier, V. Host-Feeding Preference and Diel Activity of Mosquito Vectors of the Japanese Encephalitis Virus in Rural Cambodia. Pathogens 2021, 10, 376. [Google Scholar] [CrossRef]
- Kay, B.H.; Boreham, P.F.; Williams, G.M. Host preferences and feeding patterns of mosquitoes (Diptera: Culicidae) at Kowanyama, Cape York Peninsula, northern Queensland. Bull. Entomol. Res. 1979, 69, 441–457. [Google Scholar] [CrossRef]
- Van Den Hurk, A.F.; Johansen, C.A.; Zborowski, P.; Paru, R.; Foley, P.N.; Beebe, N.W.; Mackenzie, J.S.; Ritchie, S.A. Mosquito host-feeding patterns and implications for Japanese encephalitis virus transmission in northern Australia and Papua New Guinea. Med. Vet. Entomol. 2003, 17, 403–411. [Google Scholar] [CrossRef]
- Boyle, D.B.; Dickerman, R.W.; Marshall, I.D. Primary viraemia responses of herons to experimental infection with Murray Valley encephalitis, Kunjin and Japanese encephalitis viruses. Aust. J. Exp. Biol. Med. Sci. 1983, 61, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Mani, T.R.; Rao, C.V.R.M.; Rajendran, R.; Devaputra, M.; Prasanna, Y.; Gajanana, A.; Reuben, R. Surveillance for Japanese encephalitis in villages near Madurai, Tamil Nadu, India. Trans. R. Soc. Trop. Med. Hyg. 1991, 85, 287–291. [Google Scholar] [CrossRef]
- Scherer, W.F.; Moyer, J.T.; Izumi, T. Immunologic studies of Japanese encephalitis virus in Japan. J. Immunol. 1959, 83, 620–626. [Google Scholar] [PubMed]
- Cleton, N.B.; Bosco-Lauth, A.; Page, M.J.; Bowen, R.A. Age-related susceptibility to Japanese encephalitis virus in domestic ducklings and chicks. Am. J. Trop. Med. Hyg. 2014, 90, 242–246. [Google Scholar] [CrossRef]
- Scherer, W.F. Ecological studies of Japanese encephalitis virus in Japan. Parts I–IX. Am. J. Trop. Med. Hyg. 1959, 8, 644–722. [Google Scholar] [CrossRef]
- Ladreyt, H.; Durand, B.; Dussart, P.; Chevalier, V. How central is the domestic pig in the epidemiological cycle of Japanese encephalitis virus? A review of scientific evidence and implications for disease control. Viruses 2019, 11, 949. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Fu, S.; Dai, P.; Wang, H.; Li, Y.; Li, X.; Lei, W.; Gao, X.; He, Y.; Lv, Z.; et al. Pigsties near dwellings as a potential risk factor for the prevalence of Japanese encephalitis virus in adult in Shanxi, China. Infect. Dis. Poverty 2017, 6, 100. [Google Scholar] [CrossRef] [PubMed]
- Le Flohic, G.; Porphyre, V.; Barbazan, P.; Gonzalez, J.P. Review of climate, landscape, and viral genetics as drivers of the Japanese encephalitis virus ecology. PLoS Negl. Trop. Dis. 2013, 7, e2208. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.T.; Mackenzie, J.S.; Bingham, J. Flaviviruses. In Diseases of Swine, 11th ed.; Zimmerman, J.J., Karriker, L., Ramirez, A., Schwartz, K., Stevenson, G., Zhang, J., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 530–543. [Google Scholar]
- Ricklin, M.E.; Garcìa-Nicolàs, O.; Brechbühl, D.; Python, S.; Zumkehr, B.; Posthaus, H.; Oevermann, A.; Summerfield, A. Japanese encephalitis virus tropism in experimentally infected pigs. Vet. Res. 2016, 47, 34. [Google Scholar] [CrossRef] [PubMed]
- Takashima, I.; Wantanabe, T.; Ouchi, N.; Hashimoto, N. Ecological studies of Japanese encephalitis virus in Hokkaido: Interepidemic outbreaks of swine abortion and evidence for the virus to overwinter locally. Am. J. Trop. Med. Hyg. 1988, 38, 420–427. [Google Scholar] [CrossRef]
- Lindahl, J.; Boqvist, S.; Ståhl, K.; Thu, H.T.V.; Magnusson, U. Reproductive performance in sows in relation to Japanese encephalitis virus seropositivity in an endemic area. Trop. Anim. Health Prod. 2012, 44, 239–245. [Google Scholar] [CrossRef]
- Ogasa, A.; Yokoki, Y.; Fujisaki, Y.; Habu, A. Reproductive disorders in boars infected experimentally with Japanese encephalitis virus. Jpn. J. Anim. Reprod. 1977, 23, 171–175. [Google Scholar] [CrossRef]
- Yamada, M.; Nakamura, K.; Yoshii, M.; Kaku, Y. Nonsuppurative encephalitis in piglets after experimental inoculation of Japanese encephalitis flavivirus isolated from pigs. Vet. Pathol. 2004, 41, 62–67. [Google Scholar] [CrossRef]
- Hurlbut, H.S.; Thomas, J.I. Observations on the experimental transmission of Japanese encephalitis by mosquitoes 1. Am. J. Trop. Med. Hyg. 1950, 1, 683–688. [Google Scholar] [CrossRef] [PubMed]
- Van den Hurk, A.F.; Ritchie, S.A.; Mackenzie, J.S. Ecology and geographical expansion of Japanese encephalitis virus. Annu. Rev. Entomol. 2009, 54, 17–35. [Google Scholar] [CrossRef]
- Ricklin, M.E.; García-Nicolás, O.; Brechbühl, D.; Python, S.; Zumkehr, B.; Nougairede, A.; Charrel, R.N.; Posthaus, H.; Oevermann, A.; Summerfield, A. Vector-free transmission and persistence of Japanese encephalitis virus in pigs. Nat. Commun. 2016, 7, 10832. [Google Scholar] [CrossRef]
- Di Francesco, J.; Choeung, R.; Peng, B.; Pring, L.; Pang, S.; Duboz, R.; Ong, S.; Sorn, S.; Tarantola, A.; Fontenille, D.; et al. Comparison of the dynamics of Japanese encephalitis virus circulation in sentinel pigs between a rural and a peri-urban setting in Cambodia. PLoS Negl. Trop. Dis. 2018, 12, e0006644. [Google Scholar] [CrossRef]
- Diallo, A.O.I.; Chevalier, V.; Cappelle, J.; Duong, V.; Fontenille, D.; Duboz, R. How much does direct transmission between pigs contribute to Japanese encephalitis virus circulation? A modelling approach in Cambodia. PLoS ONE 2018, 13, e0201209. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, O.; Karoji, Y.; Kuroda, A.; Karaki, T.; Takenokuma, K.; Maeda, O. Protection of pigs against mosquito-borne Japanese encephalitis virus by immunization with a live attenuated vaccine. Antivir. Res. 1982, 2, 355–360. [Google Scholar] [CrossRef]
- Khan, S.U.; Salje, H.; Hannan, A.; Islam, M.A.; Bhuyan, A.A.M.; Islam, M.A.; Rahman, M.Z.; Nahar, N.; Hossain, M.J.; Luby, S.P.; et al. Dynamics of Japanese encephalitis virus transmission among pigs in northwest Bangladesh and the potential impact of pig vaccination. PLoS Negl. Trop. Dis. 2014, 8, e3166. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.C.; Ooi, M.H.; Abdullah, A.R.; Wong, S.Y.; Krishnan, S.; Tio, P.H.; Pek, P.C.; Lai, B.F.; Mohan, A.; Muhi, J.; et al. A decade of Japanese encephalitis surveillance in Sarawak, Malaysia: 1997–2006. Trop. Med. Int. Health 2008, 13, 52–55. [Google Scholar] [CrossRef] [PubMed]
- Ting, S.H.; Tan, H.C.; Wong, W.K.; Ng, M.L.; Chan, S.H.; Ooi, E.E. Seroepidemiology of neutralizing antibodies to Japanese encephalitis virus in Singapore: Continued transmission despite abolishment of pig farming? Acta Trop. 2004, 92, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Mochizuki, H.; Sugawa, Y.; Okazaki, K.; Matumoto, M. Studies on Japanese encephalitis of cattle—1. Bovine encephalitis caused by natural infection with Japanese encephalitis virus. Natl. Inst. Anim. Health Q. 1951, 23, 111–118. [Google Scholar]
- Ilkal, M.A.; Dhanda, V.; Rao, B.U.; George, S.; Mishra, A.C.; Prasanna, Y.; Gopalkrishna, S.; Pavri, K.M. Absence of viraemia in cattle after experimental infection with Japanese encephalitis virus. Trans. R. Soc. Trop. Med. Hyg. 1988, 82, 628–631. [Google Scholar] [CrossRef]
- Gulati, B.R.; Singha, H.; Singh, B.K.; Virmani, N.; Khurana, S.K.; Singh, R.K. Serosurveillance for Japanese encephalitis virus infection among equines in India. J. Vet. Sci. 2011, 12, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Ellis, P.; Daniels, P.W.; Banks, D.J. Emerging infectious diseases: Japanese encephalitis. Vet. Clin. N. Am. Equine Pract. 2000, 16, 565. [Google Scholar] [CrossRef]
- Carey, D.E.; Reuben, R.; Myers, R.M. Japanese encephalitis studies in Vellore. South India. V. Experimental infection and transmission. Indian J. Med. Res. 1969, 57, 283. [Google Scholar]
- Ladreyt, H.; Auerswald, H.; Tum, S.; Ken, S.; Heng, L.; In, S.; Lay, S.; Top, C.; Ly, S.; Duong, V.; et al. Comparison of Japanese encephalitis force of infection in pigs, poultry and dogs in cambodian villages. Pathogens 2020, 9, 719. [Google Scholar] [CrossRef]
- Auerswald, H.; Ruget, A.S.; Ladreyt, H.; In, S.; Mao, S.; Sorn, S.; Tum, S.; Duong, V.; Dussart, P.; Cappelle, J.; et al. Serological evidence for Japanese encephalitis and West Nile virus infections in domestic birds in Cambodia. Front. Vet. Sci. 2020, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- Kako, N.; Suzuki, S.; Sugie, N.; Kato, T.; Yanase, T.; Yamakawa, M.; Shirafuji, H. Japanese encephalitis in a 114-month-old cow: Pathological investigation of the affected cow and genetic characterization of Japanese encephalitis virus isolate. BMC Vet. Res. 2014, 10, 63. [Google Scholar] [CrossRef] [PubMed]
- Lam, K.H.K.; Ellis, T.M.; Williams, D.T.; Lunt, R.A.; Daniels, P.W.; Watkins, K.L.; Riggs, C.M. Japanese encephalitis in a racing thoroughbred gelding in Hong Kong. Vet. Rec. 2005, 157, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Peiris, J.S.M.; Amerasingje, F.P.; Arunagiri, C.K.; Perera, L.P.; Karunaratne, S.H.P.P.; Ratnayake, C.B.; Kulatilaka, T.A.; Abeysinghe, M.R.N. Japanese encephalitis in Sri Lanka: Comparison of vector and virus ecology in different agro-climatic areas. Trans. R. Soc. Trop. Med. Hyg. 1993, 87, 541–548. [Google Scholar] [CrossRef]
- Cui, J.; Counor, D.; Shen, D.; Sun, G.; He, H.; Deubel, V.; Zhang, S. Detection of Japanese encephalitis virus antibodies in bats in Southern China. Am. J. Trop. Med. Hyg. 2008, 78, 1007–10011. [Google Scholar] [CrossRef]
- Wang, J.L.; Pan, X.L.; Zhang, H.L.; Fu, S.H.; Wang, H.Y.; Tang, Q.; Wang, L.F.; Liang, G.D. Japanese encephalitis viruses from bats in Yunnan, China. Emerg. Infect. Dis. 2009, 15, 939–942. [Google Scholar] [CrossRef] [PubMed]
- Sulkin, S.E.; Allen, R.; Miura, T.; Toyokawa, K. Studies of arthropod-borne virus infections in chiroptera. VI. Isolation of Japanese B encephalitis virus from naturally infected bats. Am. J. Trop. Med. Hyg. 1970, 19, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Cross, J.H. Japanese encephalitis virus surveillance in Taiwan. II. Isolations from mosquitoes and bats in Taipei area 1969–1970. J. Formos. Med. Assoc. 1971, 70, 681–686. [Google Scholar]
- Liu, S.; Li, X.; Chen, Z.; Chen, Y.; Zhang, Q.; Liao, Y.; Zhou, J.; Ke, X.; Ma, L.; Xiao, J.; et al. Comparison of genomic and amino acid sequences of eight Japanese encephalitis virus isolates from bats. Arch. Virol. 2013, 158, 2543–2552. [Google Scholar] [CrossRef] [PubMed]
- Sulkin, S.E.; Allen, R. Virus Infections in Bats. In Monographs in Virology; Karger: Basel, Switzerland, 1974; Volume 8, pp. I–VIII. [Google Scholar]
- Sulkin, S.E.; Sims, R.; Allen, R. Studies of arthropod-borne virus infections in Chiroptera. II. Experiments with Japanese B and St. Louis encephalitis viruses in the gravid bat. Evidence of transplacental transmission. Am. J. Trop. Med. Hyg. 1964, 13, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Van den Hurk, A.F.; Smith, C.S.; Field, H.E.; Smith, I.L.; Northill, J.A.; Taylor, C.T.; Jansen, C.C.; Smith, G.A.; Mackenzie, J.S. Transmission of Japanese encephalitis virus from the black flying fox, Pteropus alecto, to Culex annulirostris mosquitoes, despite the absence of detectable viremia. Am. J. Trop. Med. Hyg. 2009, 81, 457–462. [Google Scholar] [CrossRef]
- Gage, K.L.; Burkot, T.R.; Eisen, R.J.; Hayes, E.B. Climate and vectorborne diseases. Am. J. Prev. Med. 2008, 35, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Borah, J.; Dutta, P.; Khan, S.A.; Mahanta, J. Association of weather and anthropogenic factors for transmission of Japanese encephalitis in an endemic area of India. EcoHealth 2013, 10, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Xu, Z.; Zhang, J.; Mao, D.; Luo, C.; He, Y.; Liang, G.; Lu, B.; Bisesi, M.S.; Sun, Q.; et al. Regional impact of climate on Japanese encephalitis in areas located near the three gorges dam. PLoS ONE. 2014, 9, e84326. [Google Scholar]
- Wang, L.; Hu, W.; Magalhaes, R.J.; Bi, P.; Ding, F.; Sun, H.; Li, S.; Yin, W.; Wei, L.; Liu, Q.; et al. The role of environmental factors in the spatial distribution of Japanese encephalitis in mainland China. Environ. Int. 2014, 73, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hunt, S.K.; Galatowitsch, M.L.; McIntosh, A.R. Interactive effects of land use, temperature, and predators determine native and invasive mosquito distributions. Freshw. Biol. 2017, 62, 1564–1577. [Google Scholar] [CrossRef]
- Sakamoto, R.; Tanimoto, T.; Takahashi, K.; Hamaki, T.; Kusumi, E.; Crump, A. Flourishing Japanese encephalitis, associated with global warming and urbanisation in Asia, demands widespread integrated vaccination programmes. Ann. Glob. Health 2019, 85, 111. [Google Scholar] [CrossRef] [PubMed]
- Ciota, A.T.; Matacchiero, A.C.; Kilpatrick, A.M.; Kramer, L.D. The effect of temperature on life history traits of Culex mosquitoes. J. Med. Entomol. 2014, 51, 55–62. [Google Scholar] [CrossRef]
- Samuel, G.H.; Adelman, Z.N.; Myles, K.M. Temperature-dependent effects on the replication and transmission of arthropod-borne viruses in their insect hosts. Curr. Opin. Insect Sci. 2016, 16, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Parida, M.; Dash, P.K. Impact of transmission cycles and vector competence on global expansion and emergence of arboviruses. Rev. Med. Virol. 2017, 27, e1941. [Google Scholar] [CrossRef]
- Samy, A.M.; Alkishe, A.A.; Thomas, S.M.; Wang, L.; Zhang, W. Mapping the potential distributions of etiological agent, vectors, and reservoirs of Japanese encephalitis in Asia and Australia. Acta Trop. 2018, 188, 108–117. [Google Scholar] [CrossRef]
- Schuh, A.J.; Ward, M.J.; Leigh Brown, A.J.; Barrett, A.D. Phylogeography of Japanese encephalitis virus: Genotype is associated with climate. PLoS Negl. Trop. Dis. 2013, 7, e2411. [Google Scholar] [CrossRef] [PubMed]
- Huestis, D.L.; Dao, A.; Diallo, M.; Sanogo, Z.L.; Samake, D.; Yaro, A.S.; Ousman, Y.; Linton, Y.M.; Krishna, A.; Very, L.; et al. Windborne long-distance migration of malaria mosquitoes in the Sahel. Nature 2019, 574, 404–408. [Google Scholar] [CrossRef]
- Kay, B.H.; Farrow, R.A. Mosquito (Diptera: Culicidae) dispersal: Implications for the epidemiology of Japanese and Murray Valley encephalitis viruses in Australia. J. Med. Entomol. 2000, 37, 797–801. [Google Scholar] [CrossRef]
- Sellers, R.F.; Maarouf, A.R. Possible introduction of epizootic hemorrhagic disease of deer virus (serotype 2) and bluetongue virus (serotype 11) into British Columbia in 1987 and 1988 by infected Culicoides carried on the wind. Can. J. Vet. Res. 1991, 55, 367–370. [Google Scholar] [PubMed]
- Ducheyne, E.; De Deken, R.; Bécu, S.; Codina, B.; Nomikou, K.; Mangana-Vougiaki, O.; Georgiev, G.; Purse, B.V.; Hendrickx, G. Quantifying the wind dispersal of Culicoides species in Greece and Bulgaria. Geospat. Health 2007, 1, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Melville, L.; Weir, R.; Hunt, N.; Davis, S.; Walsh, S. Forty years continuous monitoring for bluetongue virus at an Australian site of high arbovirus activity. What have we achieved. Vet. Ital. 2015, 51, 263–267. [Google Scholar]
- Jacquet, S.; Huber, K.; Pagès, N.; Talavera, S.; Burgin, L.E.; Carpenter, S.; Sanders, C.; Dicko, A.H.; Djerbal, M.; Goffredo, M.; et al. Range expansion of the Bluetongue vector, Culicoides imicola, in continental France likely due to rare wind-transport events. Sci. Rep. 2016, 6, 27247. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Carrión, E.; Ivorra, B.; Ramos, Á.M.; Martinez-Lopez, B.; Aguilar-Vega, C.; Sánchez-Vizcaíno, J.M. An advection-deposition-survival model to assess the risk of introduction of vector-borne diseases through the wind: Application to bluetongue outbreaks in Spain. PLoS ONE 2018, 13, e0194573. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Yang, M.S.; Rhim, H.; Han, J.I.; Oem, J.K.; Kim, Y.H.; Lee, K.K.; Lim, C.W.; Kim, B. Analysis of five arboviruses and Culicoides distribution on cattle farms in Jeollabuk-do, Korea. Korean J. Parasitol. 2018, 56, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.A.; Rochester, W. Wind-blown mosquitoes and introduction of Japanese encephalitis into Australia. Emerg. Infect. Dis. 2001, 7, 900–903. [Google Scholar] [CrossRef]
- Metelka, J.; Robertson, C.; Stephen, C. Japanese encephalitis: Estimating future trends in Asia. AIMS Public Health 2015, 2, 601–615. [Google Scholar] [CrossRef] [PubMed]
- Sabin, A. Epidemic encephalitis in military personnel; isolation of Japanese B virus on Okinawa in 1945, serologic diagnosis, clinical manifestations, epidemiologic aspects and use of mouse brain vaccine. J. Am. Med. Assoc. 1947, 133, 281–293. [Google Scholar] [CrossRef]
- Hsu, T.C.; Chow, L.P.; Wei, H.Y.; Chen, C.L.; Hsu, S.T. A controlled field trial for an evaluation of effectiveness of mouse-brain Japanese encephalitis vaccine. J. Formos. Med. Assoc. 1971, 70, 55–62. [Google Scholar]
- Rao Bhau, L.; Singh, G.; Gowal, D.; Saxena, S.; Kobayashi, M.; Oya, A.; Yoshioka, I. Safety and efficacy of Japanese encephalitis vaccine produced in India. Indian J. Med. Res. 1988, 88, 301–307. [Google Scholar]
- Kanamitsu, M.; Hashimoto, N.; Urasawa, S.; Katsurada, M.; Kimura, H. A field trial with an improved Japanese encephalitis vaccine in a nonendemic area of the disease. Biken J. 1970, 13, 313–328. [Google Scholar] [PubMed]
- Fukunaga, T.; Rojanasuphot, S.; Wungkobkiat, S.; Thammanichanon, A.; Kan, T. Japanese encephalitis vaccination in Thailand. Biken J. 1974, 17, 21–31. [Google Scholar] [PubMed]
- Poland, J.D.; Bruce Cropp, C.; Craven, R.B.; Monath, T.P. Evaluation of the Potency and Safety of Inactivated Japanese Encephalitis Vaccine in US Inhabitants. J. Infect. Dis. 1990, 161, 878–882. [Google Scholar] [CrossRef] [PubMed]
- Plesner, A.-M.; Rønne, T.; Wachmann, H. Case-control study of allergic reactions to Japanese encephalitis vaccine. Vaccine 2000, 18, 1830–1836. [Google Scholar] [CrossRef]
- Yu, Y. Phenotypic and genotypic characteristics of Japanese encephalitis attenuated live vaccine virus SA14-14-2 and their stabilities. Vaccine 2010, 28, 3635–3641. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, S.; Strom, B.L.; Bilker, W.B.; Zhengle, L.; Chao-Min, W.; Hui-Lian, L.; Tai-Xiang, W.; Hong-Ji, Y.; Qi-Mau, L.; Tsai, T.F.; et al. Effectiveness of live-attenuated Japanese encephalitis vaccine (SA14-14-2): A case-control study. Lancet 1996, 347, 1583–1586. [Google Scholar] [CrossRef]
- Ohrr, H.; Tandan, J.B.; Sohn, Y.M.; Shin, S.H.; Pradhan, D.P.; Halstead, S.B. Effect of single dose of SA 14-14-2 vaccine 1 year after immunisation in Nepalese children with Japanese encephalitis: A case-control study. Lancet 2005, 366, 1375–1378. [Google Scholar] [CrossRef]
- Wu, W.; Liu, D.; Li, K.; Nuorti, J.P.; Nohynek, H.M.; Xu, D.; Ye, J.; Zheng, J.; Wang, H. Post-marketing safety surveillance for inactivated and live-attenuated Japanese encephalitis vaccines in China, 2008–2013. Vaccine 2017, 35, 3666–3671. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Japanese encephalitis vaccines: WHO position paper—February 2015. Wkly. Epidemiol. Rec. 2015, 90, 69–88. [Google Scholar]
- Chokephaibulkit, K.; Houillon, G.; Feroldi, E.; Bouckenooghe, A. Safety and immunogenicity of a live attenuated Japanese encephalitis chimeric virus vaccine (IMOJEV®) in children. Expert Rev. Vaccines 2016, 15, 153–166. [Google Scholar] [CrossRef]
- Cao, L.; Fu, S.; Gao, X.; Li, M.; Cui, S.; Li, X.; Cao, Y.; Lei, W.; Lu, Z.; He, Y.; et al. Low protective efficacy of the current Japanese encephalitis vaccine against the emerging genotype 5 Japanese encephalitis virus. PLoS Negl. Trop. Dis. 2016, 10, e0004686. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Cha, G.-W.; Jeong, Y.E.; Lee, W.-G.; Chang, K.S.; Roh, J.Y.; Yang, S.C.; Park, M.Y.; Park, C.; Shin, E.H. Detection of Japanese encephalitis virus genotype V in Culex orientalis and Culex pipiens (Diptera: Culicidae) in Korea. PLoS ONE 2015, 10, e0116547. [Google Scholar] [CrossRef]
- Li, M.-H.; Fu, S.-H.; Chen, W.-X.; Wang, H.-Y.; Guo, Y.-H.; Liu, Q.-Y.; Li, Y.-X.; Luo, H.-M.; Da, W.; Duo Ji, D.Z.; et al. Genotype V Japanese encephalitis virus is emerging. PLoS Negl. Trop. Dis. 2011, 5, e1231. [Google Scholar] [CrossRef] [PubMed]
- Erra, E.O.; Askling, H.H.; Yoksan, S.; Rombo, L.; Riutta, J.; Vene, S.; Lindquist, L.; Vapalahti, O.; Kantele, A. Cross-protective capacity of Japanese encephalitis (JE) vaccines against circulating heterologous JE virus genotypes. Clin. Infect. Dis. 2013, 56, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.C.; Chen, J.M.; Chiu, H.C.; Chen, Y.Y.; Lin, J.W.; Shih, C.C.; Chen, C.M.; Chang, C.C.; Chang, G.J.; Chiou, S.S. Partially neutralizing potency against emerging genotype I virus among children received formalin-inactivated Japanese encephalitis virus vaccine. PLoS Negl. Trop. Dis. 2012, 6, e1834. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.C.; Chen, J.M.; Chen, Y.Y.; Lin, J.W.; Chiou, S.S. Reduced neutralizing antibody titer against genotype I virus in swine immunized with a live-attenuated genotype III Japanese encephalitis virus vaccine. Vet. Microbiol. 2013, 163, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.K.; Hwang, J.M.; Moon, H.; Han, S.Y.; Kim, J.M.; Yang, D.K.; Park, B.K.; Song, D. Comparison of the antigenic relationship between Japanese encephalitis virus genotypes 1 and 3. Clin. Exp. Vaccine Res. 2016, 5, 26–30. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Summary of WHO Position Papers—Recommendations for Routine Immunization. 2020. Available online: http://www.who.int/immunization/policy/Immunization_routine_table1.pdf?ua=1 (accessed on 1 August 2021).
- Department of Health Services Ministry of Health & Population, Nepal. National Immunization Program, Comprehensive Multi-Year Plan 2068–2072 (2011–2016). 2011. Available online: https://extranet.who.int/countryplanningcycles/sites/default/files/planning_cycle_repository/nepal/nepal_cmyp_2011-2016.pdf (accessed on 1 August 2021).
- Dumre, S.P.; Shakya, G.; Na-Bangchang, K.; Eursitthichai, V.; Rudi Grams, H.; Upreti, S.R.; Ghimire, P.; Kc, K.; Nisalak, A.; Gibbons, R.V.; et al. Dengue virus and Japanese encephalitis virus epidemiological shifts in Nepal: A case of opposing trends. Am. J. Trop. Med. Hyg. 2013, 88, 677–680. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Japanese Encephalitis Vaccination Campaign Achievement. 2018. Available online: http://www.searo.who.int/myanmar/areas/JEcampaignachievement/en/ (accessed on 1 August 2021).
- Suraratdecha, C.; Levin, C.; Jacobson, J.; La Force, M. Demand-driven and affordable next generation vaccines for preventing Japanese encephalitis in Asia and meningococcal meningitis in Sub-Saharan Africa. In Proceedings of the Sixth International Health Economics Association World Congress; Explorations in Health Economics, Copenhagen, Denmark, 8–11 July 2007. [Google Scholar]
- World Health Organization (WHO). Japanese Encephalitis: Vaccine Preventable Diseases, Surveillance Standards. 2018. Available online: https://www.who.int/publications/m/item/vaccine-preventable-diseases-surveillance-standards-je (accessed on 1 August 2021).
- Centers for Disease Control and Prevention (CDC). Expanding poliomyelitis and measles surveillance networks to establish surveillance for acute meningitis and encephalitis syndromes--Bangladesh, China, and India, 2006–2008. Morb. Mortal. Wkly. Rep. 2012, 61, 1008–10011. [Google Scholar]
- Heffelfinger, J.; Xi, L.; Batmunkh, N.; Grabovac, V.; Diorditsa, S.; Liyanage, J.; Pattamadilok, S.; Bahl, S.; Vannice, K.; Hyde, T.; et al. Japanese encephalitis surveillance and immunization—Asia and Western Pacific Regions, 2016. Morb. Mortal. Wkly. Rep. 2017, 66, 579–583. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulvey, P.; Duong, V.; Boyer, S.; Burgess, G.; Williams, D.T.; Dussart, P.; Horwood, P.F. The Ecology and Evolution of Japanese Encephalitis Virus. Pathogens 2021, 10, 1534. https://doi.org/10.3390/pathogens10121534
Mulvey P, Duong V, Boyer S, Burgess G, Williams DT, Dussart P, Horwood PF. The Ecology and Evolution of Japanese Encephalitis Virus. Pathogens. 2021; 10(12):1534. https://doi.org/10.3390/pathogens10121534
Chicago/Turabian StyleMulvey, Peter, Veasna Duong, Sebastien Boyer, Graham Burgess, David T. Williams, Philippe Dussart, and Paul F. Horwood. 2021. "The Ecology and Evolution of Japanese Encephalitis Virus" Pathogens 10, no. 12: 1534. https://doi.org/10.3390/pathogens10121534
APA StyleMulvey, P., Duong, V., Boyer, S., Burgess, G., Williams, D. T., Dussart, P., & Horwood, P. F. (2021). The Ecology and Evolution of Japanese Encephalitis Virus. Pathogens, 10(12), 1534. https://doi.org/10.3390/pathogens10121534