
Trypanosomatid Richness in Wild and Synanthropic Small Mammals from a Biological Station in Rio de Janeiro, Brazil

 ,
, 

Abstract
1. Introduction
2. Results
2.1. Small Mammals and Their Sampling Areas
2.2. Infection Rates of Trypanosomatids
2.3. Parasitological and Molecular Diagnosis
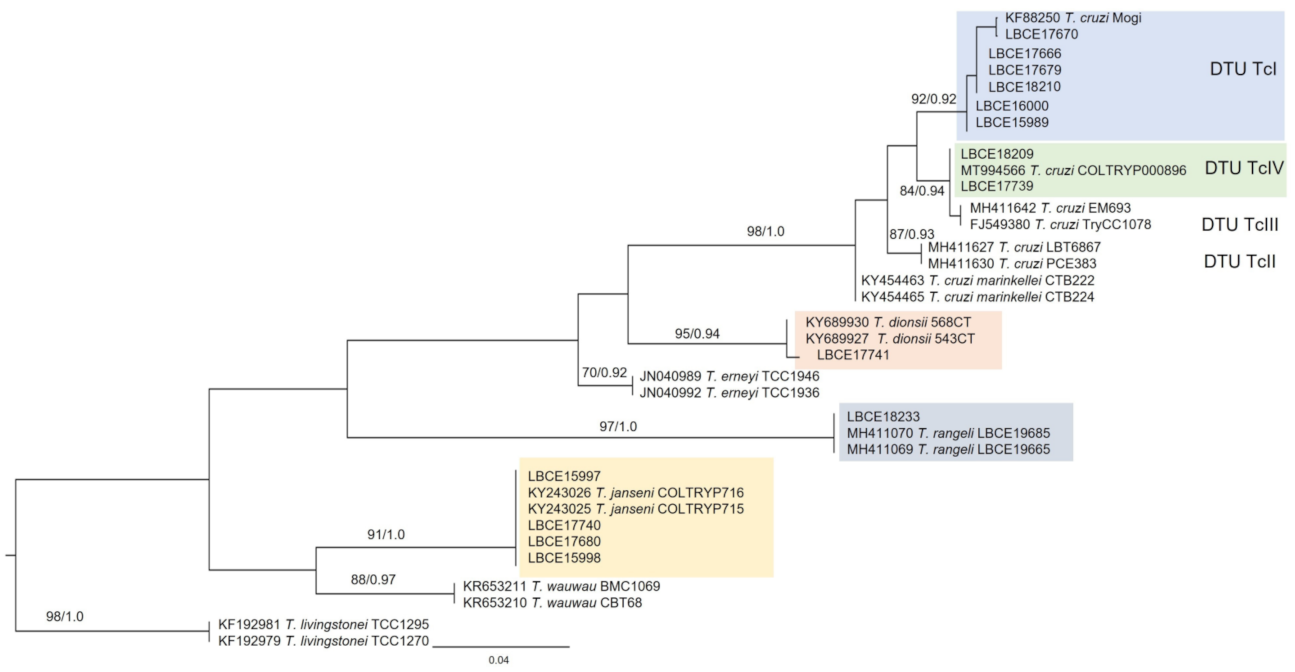
2.4. Phylogenetic Analysis of Trypanosomatids Characterized at the Species Level
2.5. Serological Diagnosis
2.6. Recaptures
- LBCE 17825: captured twice in A2. It was positive in serology for T. cruzi and Leishmania spp., and T. cruzi DTU TcI was isolated from the blood. Four months later, only serological infection was observed, with the same IFAT titers (1/40 T. cruzi; 1/80 Leishmania spp.);
- LBCE 17674: captured twice in A2. In the first capture, it was not positive for any of the diagnostic assays, and showed positivity in the fresh blood examination only in the second capture (nine months later);
- LBCE 18232: captured twice in A1 (three months interval). It was positive in serology, for T. cruzi and Leishmania spp. The IFAT titers were different for Leishmania spp. (1:40 and 1:80), while the IFAT titers for T. cruzi remained the same (1:160);
- LBCE 18255: first captured in A2, and four months later in A1 it was positive in serology for T. cruzi, showing different IFAT titers in the first (1:160) and second capture (1:80), while it was serologically positive for Leishmania spp. only in the first capture (1:40). In the second capture, T. janseni was isolated in the blood culture.
3. Discussion
- positive in the fresh blood examination in the second capture, probably showing an increase in parasitemia or infection by other trypanosomatid parasites;
- positive blood culture for T. cruzi DTU TcI only in the first capture, with the expected decrease in parasitemia in the late phase of infection, as this parasite was not detected in the second capture; and
- positive blood culture for T. janseni only in the second capture, probably because that host became infected after the first capture. The latter was recaptured after four months in a different area. This result indicates that individuals of D. aurita can move across different areas in the study site. This is not a surprising finding considering that D. aurita commonly covers long distances during its lifetime [13,44].
4. Materials and Methods
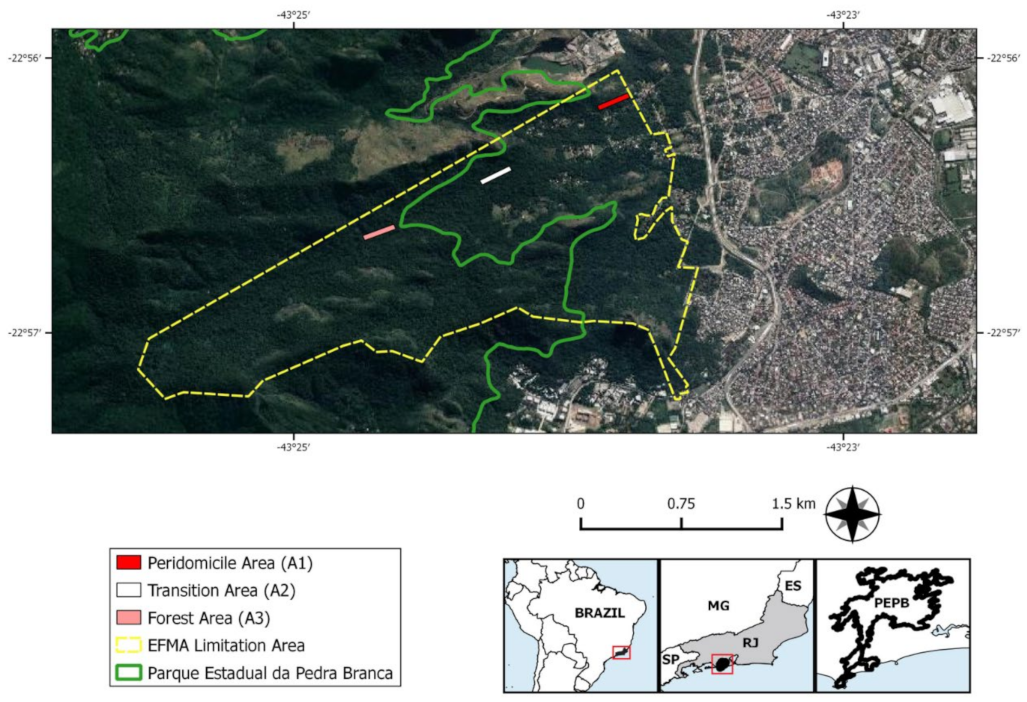
4.1. Study Area
- peridomicile (A1): representing areas adjacent to human dwellings;
- transition (A2): disturbed forest and reforestation areas between the peridomicile and the preserved forest; and
- preserved forest (A3): the most preserved and distant area from the human dwellings composed of a secondary forest of ombrophilous dense vegetation. See Gentile et al. (2018) [26] for a better description of the study areas.
4.2. Small Wild Mammal Capture and Identification
4.3. Field Procedures
- sterile saline (sodium chloride-NaCl at 58.44 g/mol), antibiotics, and antifungals (10 mg streptomycin, 25 µL amphotericin B, and 10,000 IU penicillin per mL, Sigma™, St, Louis, MO, USA commercial solution) for culture; and
- absolute ethanol that was stored in a freezer at −20 °C for subsequent molecular diagnosis.
4.4. Parasitological Procedures
4.5. Serological Diagnosis
4.6. Molecular Diagnosis
- kDNA target, used to detect Leishmania spp. Infections;
- HSP70 (234) target, employed in Leishmania spp. kDNA-positive samples;
- the 18S rDNA target for the detection of Trypanosomatidae, and for the characterization of all positive culture samples; and
- the 24S rDNA target for 18S-positive samples in which characterization was not possible due to the low quality of the DNA sequences obtained.
- a final volume of 25 µL was used, containing 13.5 µL of ultrapure water, 8.5 µL of Go Taq Master Mix (Promega, Madison, WI, USA), and 2 µL of DNA in both rounds;
- in the first round, 0.5 µL of the primers TRY R and F (16 pmol) (Eurofins Genomics™, Val Fleuri, Luxembourg, Luxembourg) were used, and in the second round, 0.5 µL of the primers SSU R and F (16 pmol) (Eurofins Genomics™, Val Fleuri, Luxembourg, Luxembourg) were used; and
- in the cycling condition, the initial denaturation occurred at 95 °C for 15 min, in a Swift™ Max Pro Thermal Cycler 16 thermal cycler (model SWT-MXP-BLC-1).
4.7. Phylogenetic Analyses
4.8. Statistical Analysis
4.9. Ethics Statement
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- D’Avila-Levy, C.M.; Boucinha, C.; Kostygov, A.; Santos, H.L.C.; Morelli, K.A.; Grybchuk-Ieremenko, A.; Duval, L.; Votýpka, J.; Yurchenko, V.; Grellier, P.; et al. Exploring the environmental diversity of kinetoplastid flagellates in the high-throughput DNA sequencing era. Memórias Inst. Oswaldo Cruz 2015, 110, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Votýpka, J.; D’Avila-Levy, C.M.; Grellier, P.; Maslov, D.A.; Lukeš, J.; Yurchenko, V. New approaches to systematics of Trypanosomatidae: Criteria for taxonomic (re)description. Trends Parasitol. 2015, 31, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Kaufer, A.; Ellis, J.; Stark, D.; Barratt, J. The evolution of trypanosomatid taxonomy. Parasites Vectors 2017, 10, 287. [Google Scholar] [CrossRef] [PubMed]
- Maslov, D.A.; Opperdoes, F.R.; Kostygov, A.Y.; Hashimi, H.; Lukeš, J.; Yurchenko, V. Recent advances in trypanosomatid research: Genome organization, expression, metabolism, taxonomy and evolution. Parasitology 2018, 146, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Page, A.M.; Canning, E.U.; Barker, R.J.; Nicholas, J.P. A new species of Rhynchoidomonas Patton, 1910 (Kinetoplastida: Trypanosomatina) from Operophtera brumata (Lepidoptera: Geometridae). Syst. Parasitol. 1986, 8, 101–105. [Google Scholar] [CrossRef]
- Akhoundi, M.; Downing, T.; Votýpka, J.; Kuhls, K.; Luke, J.; Cannet, A.; Ravel, C.; Marty, P.; Delaunay, P.; Kasbari, M.; et al. Leishmania infections: Molecular targets and diagnosis. Mol. Asp. Med. 2017, 57, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Ereqat, S.; Nasereddin, A.; Al-Jawabreh, A.; Al-Jawabreh, H.; Al-Laham, N.; Abdeen, Z. Prevalence of Trypanosoma evansi in livestock in Palestine. Parasites Vectors 2020, 13, 21. [Google Scholar] [CrossRef]
- Roman, F.; Iñiguez, A.M.; Yeo, M.; Jansen, A.M. Multilocus sequence typing: Genetic diversity in Trypanosoma cruzi I (TcI) isolates from Brazilian didelphids. Parasites Vectors 2018, 11, 107. [Google Scholar] [CrossRef] [PubMed]
- Zingales, B. Trypanosoma cruzi: Um parasita, dois parasitas ou vários parasitas da doença de Chagas? Rev. Biol. 2011, 6, 44–48. [Google Scholar] [CrossRef]
- Jansen, A.M.; Xavier, S.C.D.C.; Roque, A.L.R. Trypanosoma cruzi transmission in the wild and its most important reservoir hosts in Brazil. Parasites Vectors 2018, 11, 502. [Google Scholar] [CrossRef]
- Patton, J.L.; Pardiñas, U.F.J.; D’elía, G. Mammals of South America: Rodents; The University of Chicago Press: Chicago, IL, USA, 2015; Volume 2, p. 1384. [Google Scholar]
- Cássia-Pires, R.; Boite, M.C.; D’Andrea, P.S.; Herrera, H.M.; Cupolillo, E.; Jansen, A.M.; Roque, A.L.R. Distinct Leishmania Species Infecting Wild Caviomorph Rodents (Rodentia: Hystricognathi) from Brazil. PLoS Negl. Trop. Dis. 2014, 8, e3389. Available online: www.plosntds.org (accessed on 15 October 2021). [CrossRef]
- Roque, A.L.R.; Jansen, A.M. Reservatórios do Trypanosoma cruzi e sua relação com os vetores. In Vetores da doença de chagas no Brasil, Curitiba: Sociedade Brasileira de Zoologia; Zoologia: Guias e Manuais de Identificação, Series; Galvão, C., Ed.; Sociedade Brasileira de Zoologia: Curitiba, Brazil, 2014; pp. 75–87. ISBN 978-85-98203-09-6. [Google Scholar]
- Caldart, E.T.; Freire, R.L.; Ferreira, F.P.; Ruffolo, B.B.; Sbeghen, M.R.; Mareze, M.; Garcia, J.L.; Mitsuka-Breganó, R.; Navarro, I.T. Leishmania in synanthropic rodents (Rattus rattus): New evidence for the urbanization of Leishmania (Leishmania) amazonensis. Braz. J. Vet. Parasitol. 2017, 26, 17–27. [Google Scholar] [CrossRef]
- Morales, E.A.; Mayor, P.; Bowler, M.; Aysanoa, E.; Pérez-Velez, E.S.; Pérez, J.; Ventocilla, J.A.; Baldeviano, C.C.; Lescano, A.G. Prevalence of Trypanosoma cruzi and Other Trypanosomatids in Frequently-Hunted Wild Mammals from the Peruvian Amazon. Am. J. Trop. Med. Hyg. 2017, 97, 1482–1485. [Google Scholar] [CrossRef]
- Rodrigues, M.S.; Lima, L.; Xavier, S.C.C.; Herrera, H.M.; Rocha, F.L.; Roque, A.L.R.; Teixeira, M.M.G.; Jansen, A.M. Uncovering Trypanosoma spp. diversity of wild mammals by the use of DNA from blood clots. Parasites Wildl. 2019, 8, 171–181. [Google Scholar] [CrossRef]
- Lopes, C.M.T.; Barreto, R.F.S.M.; Pavan, M.G.; Pereira, C.S.; Roque, A.L.R. Trypanosoma janseni n. sp. (Trypanosomatida: Trypanosomatidae) isolated from Didelphis aurita (Mammalia: Didelphidae) in the Atlantic Rainforest of Rio de Janeiro, Brazil: Integrative taxonomy and phylogeography within the Trypanosoma cruzi clade. Memórias Inst. Oswaldo Cruz 2018, 113, 45–55. [Google Scholar] [CrossRef]
- Tahir, D.; Davousta, B.; Heuc, K.; Lamourb, T.; Demarc, M.; Mariéa, J.L.; Blanchet, D. Molecular and serological investigation of Trypanosoma cruzi infection in dogs in French Guiana. Vet. Parasitol. Reg. Stud. Rep. 2018, 12, 106–109. [Google Scholar] [CrossRef]
- Barros, J.H.S.; Toma, H.K.; Madeira, M.D.F. Molecular study of Trypanosoma caninum isolates based on different genetic markers. Parasitol. Res. 2015, 114, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.K.; Godfrey, S.S.; Thompson, R.C.A. Trypanosomes of Australian mammals: A review. Int. J. Parasitol. Parasites Wildl. 2014, 3, 57–66. [Google Scholar] [CrossRef]
- Jansen, A.M.; Xavier, S.C.; Roque, A.L. The multiple and complex and changeable scenarios of the Trypanosoma cruzi transmission cycle in the sylvatic environment. Acta Trop. 2015, 151, 1–15. [Google Scholar] [CrossRef] [PubMed]
- SOS Mata Atlântica. Available online: https://www.sosma.org.br/noticias/desmatamento-da-mata-atlantica-cresce-em-dez-estados/ (accessed on 22 September 2021).
- Rezende, C.L.; Scarano, F.R.; Assad, E.D.; Joly, C.A.; Metzgerf, J.P.; Strassburg, B.B.N.; Tabarelli, M.; Fonseca, G.A.; Mittermeier, R.A. From hotspot to hopespot: An opportunity for the Brazilian Atlantic Forest. Perspect. Ecol. Conserv. 2018, 16, 208–214. [Google Scholar] [CrossRef]
- INSTITUTO ESTADUAL DO AMBIENTE (INEA). BIODIVERSIDADE E ÁREAS PROTEGIDAS/Unidades de Conservação. Parque Estadual da Pedra Branca. Available online: http://www.inea.rj.gov.br/biodiversidade-territorio/conheca-as-unidades-de-conservacao/parque-estadual-da-pedra-branca/ (accessed on 22 September 2021).
- De Souza, N.A.; Da Silva, J.B.; Godoy, R.E.; De Souza, F.J.M.; De Andrade-Coelho, C.A.; Da Silva, V.C.; De Azevedo, A.C.R.; Rangel, E.F. Studies on Phlebotominae (Diptera: Psychodidae) in the Campus FIOCRUZ Mata Atlântica, Jacarepaguá, in the City of Rio de Janeiro, Brazil. Rev. Soc. Bras. Med. Trop. 2015, 48, 26–32. [Google Scholar] [CrossRef]
- Gentile, R.; Cardoso, T.S.; Costa-Neto, S.F.; Teixeira, B.R.; D’Andrea, P.S. Community structure and population dynamics of small mammals in an urban-sylvatic interface area in Rio de Janeiro, Brazil. Zoologia 2018, 35, e13465. [Google Scholar] [CrossRef]
- Rangel, D.A.; Lisboa, C.V.; Novaes, R.L.M.; Silva, B.A.; Souza, R.D.F.; Jansen, A.M.; Moratelli, R.; Roque, A.L.R. Isolation and characterization of trypanosomatids, including Crithidia mellificae, in bats from the Atlantic Forest of Rio de Janeiro, Brazil. PLoS Negl. Trop. Dis. 2019, 13, e0007527. [Google Scholar] [CrossRef]
- Costa-Neto, S.F.; Cardoso, T.S.; Boullosa, R.G.; Maldonado, A., Jr.; Gentile, R. Metacommunity structure of the helminths of the black-eared opossum Didelphis aurita in peri-urban, sylvatic and rural environments in south-eastern Brazil. J. Helminthol. 2019, 93, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Madeira, M.F.; Sousa, M.A.; Barros, J.H.S.; Figueiredo, F.B.; Fagundes, A.; Schubach, A.; De Paula, A.A.; Faissal, B.N.S.; Fonseca, T.S.; Thoma, H.K.; et al. Trypanosoma caninum n. sp. (Protozoa: Kinetoplastida) isolated from intact skin of domestic dog (Canis familiaris) captured in Rio de Janeiro, Brazil. Parasitology 2009, 136, 411–423. [Google Scholar] [CrossRef]
- Smith, A.; Clark, P.; Averis, S.; Lymbery, A.J.; Wayne, A.F.; Morris, K.D.; Thompsom, R.C.A. Trypanosomes in a declining species of threatened Australian marsupial, the brush-tailed bettong Bettongia penicillata (Marsupialia: Potoridae). Parasitology 2008, 135, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Brandão, E.M.V.; Xavier, S.C.C.; Carvalhaes, J.G.; D’Andrea, P.S.; Lemos, F.G.; Azevedo, F.C.; Cássia-Pires, R.; Jansen, A.M.; Roque, A.L.R. Trypanosomatids in Small Mammals of an Agroecosystem in Central Brazil: Another Piece in the Puzzle of Parasite Transmission in an Anthropogenic Landscape. Pathogens 2019, 8, 190. [Google Scholar] [CrossRef]
- Dario, M.A.; Rodrigues, M.S.; Barros, J.H.; Xavier, S.C.D.C.; D’andrea P., S.; Roque, A.L.; Jansen, A.M. Ecological scenario and Trypanosoma cruzi DTU characterization of a fatal acute Chagas disease case transmitted orally (Espírito Santo state, Brazil). Parasites Vectors 2016, 9, 477. [Google Scholar] [CrossRef] [PubMed]
- Dario, M.A.; Lisboa, C.V.; Costa, L.M.; Moratelli, R.; Nascimento, M.P.; Costa, L.P.; Leite, Y.L.R.; Llewellyn, M.S.; Xavier, S.C.D.C.; Roque, A.L.R.; et al. High Trypanosoma spp. diversity is maintained by bats and triatomines in Espírito Santo state, Brazil. PLoS ONE 2017, 12, e0188412. [Google Scholar] [CrossRef]
- Dario, M.A.; Moratelli, R.; Schwabl, P.; Jansen, A.M.; Llewellyn, M.S. Small subunit ribosomal metabarcoding reveals extraordinary trypanosomatid diversity in Brazilian bats. PLoS Negl. Trop. Dis. 2017, 11, e0005790. [Google Scholar] [CrossRef]
- Malavazi, P.F.N.S.; Daudt, C.; Melchior, L.A.K.; Meneguetti, D.U.O.; Xavier, S.C.C.; Jansen, A.M.; Souza, S.F.; Roque, A.R.L. Trypanosomes of vectors and domestic dogs in Trypanosoma cruzi transmission areas from Brazilian southwestern amazon: New mammalian host for Trypanosoma janseni. Acta Trop. 2020, 210, 105504. [Google Scholar] [CrossRef]
- Da Silva, F.M.; Junqueira, A.C.V.; Campaner, M.; Rodrigues, A.C.; Crisante, G.; Ramirez, L.E.; Caballero, Z.C.E.; Monteiro, F.A.; Coura, J.R.; Añez, N.; et al. Comparative phylogeography of Trypanosoma rangeli and Rhodnius (Hemiptera: Reduviidae) supports a long coexistence of parasite lineages and their sympatric vectors. Mol. Ecol. 2007, 16, 3361–3373. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, F.M.; Marcili, A.; Lima, L.; Cavazzana, M.J.R.; Ortiz, P.A.; Campaner, M.; Takeda, G.F.; Paiva, F.; Nunes, V.L.B.; Camargo, E.P.; et al. Trypanosoma rangeli isolates of bats from Central Brazil: Genotyping and phylogenetic analysis enable description of a new lineage using spliced-leader gene sequences. Acta Trop. 2009, 109, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Dario, M.A.; Pavan, M.G.; Rodrigues, M.S.; Lisboa, C.V.; Kluyber, D.; Desbiez, A.L.J.; Herrera, H.M.; Roque, A.L.R.; Lima, L.; Teixeira, M.M.G.; et al. Trypanosoma rangeli genetic, mammalian hosts and geographical diversity from five Brazilian biomes. Pathogens 2021, 10, 736. [Google Scholar] [CrossRef]
- Álvarez, O.E.; Ortiz, P.A.; Lima, L.; Costa-Martins, A.G.; Serrano, M.G.; Herder, S.; Buck, G.A.; Camargo, E.P.; Hamilton, P.B.; Stevens, J.R.; et al. Trypanosoma rangeli is phylogenetically closer to Old World trypanosomes than to Trypanosoma cruzi. Int. J. Parasitol. 2018, 48, 569–584. [Google Scholar] [CrossRef]
- Ramirez, L.E.; Machado, M.I.; Maywald, P.G.; Matos, A.; Chiari, E.; Silva, E.L. Primeira evidência de Trypanosoma rangeli no sudeste do Brasil, região endêmica para doença de Chagas. Rev. Soc. Bras. Med. Trop. 1998, 31, 99–102. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brandão, E.M.V.; Xavier, S.C.C.; Rocha, F.L.; Lima, C.F.M.; Candeias, Í.Z.; Lemos, F.G.; Azevedo, F.C.; Jansen, A.M.; Roque, A.L.R. Wild and Domestic Canids and Their Interactions in the Transmission Cycles of Trypanosoma Cruzi and Leishmania spp. in an Area of the Brazilian Cerrado. Pathogens 2020, 9, 818. [Google Scholar] [CrossRef]
- Cássia-Pires, R.; De Melo, M.D.F.A.D.; Barbosa, R.D.H.; Roque, A.L.R. Multiplex PCR as a tool for the diagnosis of Leishmania spp. kDNA and the gapdh housekeeping gene of mammal hosts. PLoS ONE 2017, 12, e0173922. [Google Scholar] [CrossRef]
- Folgueira, C.; Quijada, L.; Soto, M.; Abanades, D.R.; Alonso, C.; Requena, J.M. The Translational Efficiencies of the Two Leishmania infantum HSP70 mRNAs, Differing in Their 3-Untranslated Regions, Are Affected by Shifts in the Temperature of Growth through Different Mechanisms. J. Biol. Chem. 2005, 280, 35172–35183. [Google Scholar] [CrossRef]
- Gentile, R.; Cerqueira, R. Movement patterns of five species of small mammals in a Brazilian restinga. J. Trop. Ecol. 1995, 11, 671–677. [Google Scholar] [CrossRef]
- Agência FIOCRUZ de Notícias. Fiocruz inaugura primeira estação biológica do município do Rio. 11 October 2016. Available online: https://portal.fiocruz.br/noticia/fiocruz-inaugura-primeira-estacao-biologica-do-municipio-do-rio (accessed on 22 September 2021).
- Agência FIOCRUZ de Notícias. Fundação promove seminários sobre biodiversidade e saúde. 11 July 2017. Available online: https://agencia.fiocruz.br/fundacao-promove-seminarios-sobre-biodiversidade-e-saude (accessed on 22 September 2021).
- Neves, D.P. Parasitologia Humana, 11th ed.; Atheneu: São Paulo, Brazil, 2004; p. 494. [Google Scholar]
- Gomes, Y.M. Diagnóstico Etiológico. In Doença de Chagas; Malta, J. (Org), Editora Savier: São Paulo, Brazil, 1996; pp. 119–132. [Google Scholar]
- Rocha, F.L.; Roque, A.L.R.; De Lima, J.S.; Cheida, C.C.; Lemos, F.G.; Azevedo, F.C.; Arrais, R.C.; Bilac, D.; Herrera, H.M.; Mourão, G.; et al. Trypanosoma cruzi Infection in Neotropical Wild Carnivores (Mammalia: Carnivora): At the Top of the T. cruzi Transmission Chain. PLoS ONE 2013, 8, e67463. [Google Scholar] [CrossRef]
- Camargo, M.E. Fluorescent antibody test for the serodiagnosis of American trypanosomiasis. Technical modification employing preserved culture forms of Trypanosoma cruzi in a slide test. Rev. Inst. Med. Trop. São Paulo 1966, 8, 227–235. [Google Scholar] [PubMed]
- Deane, M.P.; Sousa, M.A.; Pereira, N.M.; Gonçalves, A.; Momem, H.; Morel, C. Trypanosoma cruzi: Inoculation schedules and re-isolation methods demonstrated by schizodeme and zymodeme analyses. J. Protozool. 1984, 31, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Jansen, A.M.; Moriearty, P.L.; Castro, B.G.; Deane, M.P. Trypanosoma cruzi in the opossum Didelphis marsupialis: An indirect fluorescent antibody test for the diagnosis and follow-up of natural and experimental infections. Trans. R. Soc. Trop. Med. Hyg. 1985, 79, 474–477. [Google Scholar] [CrossRef]
- Roque, A.L.R.; Xavier, S.C.; da Rocha, M.G.; Duarte, A.C.; D’Andrea, P.S.; Jansen, A.M. Trypanosoma cruzi transmission cycle among wild and domestic mammals in three areas of orally transmitted Chagas disease outbreaks. Am. J. Trop. Med. Hyg. 2008, 79, 742–749. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989; p. 1626. [Google Scholar]
- Degrave, W.; Fernandes, O.; Campbell, D.; Bozza, M.; Lopes, U. Use of molecular probes and PCR for detection and typing of Leishmania—A mini-review. Memórias Inst. Oswaldo Cruz 1994, 89, 463–469. [Google Scholar] [CrossRef]
- Da Graça, G.C.; Volpini, A.C.; Romero, G.A.S.; Oliveira Neto, M.P.; Hueb, M.; Porrozzi, R.; Boité, M.C.; Cupolillo, E. Development and validation of PCR-based assays for diagnosis of American cutaneous leishmaniasis and identification of the parasite species. Memórias Inst. Oswaldo Cruz 2012, 107, 664–674. [Google Scholar] [CrossRef]
- Arruda, M.V.; Reinach, F.C.; Colli, W.; Zingales, B. Sequence of the 24Set ribosomal RNA gene and characterization of a corresponding pseudogene from Trypanos. Cruzi. Mol. Biochem. Parasitol. 1990, 40, 35–42. [Google Scholar] [CrossRef]
- LASERGENE. User’s Guide: A Manual for the Lasergene System; [s.l.] Biocomputing Software for Windows; DNASTAR, Inc: Madison, WI, USA, 1994. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MRBAYES 3: Bayesian Phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure forphylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]



{kind=link}
{kind=link}
| Order (n) | Species | Infected/Total | A1 | A2 | A3 |
|---|---|---|---|---|---|
| Akodon cursor | 4/7 (57.5%, CI: 18.4–90.1) | 4/7 (57.5%) | - | - | |
| Rodentia (16) | Oligoryzomys nigripes | 0/2 (0%, CI: 00.0–84.2) | 0/1 (0%) | 0/1 (0%) | - |
| Rattus rattus | 7/7 (100%, CI: 100–100) | 5/5 (100%) | 2/2 (100%) | - | |
| Didelphis aurita | 52/70 (74.3%, CI: 62.4–83.9) | 26/36 (72.2%) | 20/26 (76.9%) | 6/8 (75%) | |
| Didelphimorphia (75) | Marmosa paraguayana | 4/4 (100%, CI: 100–100) | 1/1 (100%) | - | 3/3 (100%) |
| Metachirus myosurus | 1/1 (100%) | - | 1/1 (100%) | - | |
| 91 | 6 | 68/91 (74.7%) | 36/50 (72%) | 23/30 (76.7%) | 9/11 (81.2%) |
| Collection Environment | Sample ID | Species | Molecular Analysis of the Trypanosomatids Detected in Cultures | Trypanosomatids Detected in Molecular Diagnosis Directly in Tissues | GenBank Access Number | COLTRYP Number |
|---|---|---|---|---|---|---|
| A1 | LBCE 15991 | Akodon cursor | Negative | Trypanosomatidae (Sp)/T. cruzi DTU TcI **** (S) | MZ221936 (S) | - |
| LBCE 15994 | Akodon cursor | Negative | T. cruzi DTU TcI *** (Sp) | MZ229972 (Sp) | - | |
| LBCE 15995 | Rattus rattus | Negative | T. cruzi DTU TcI *** (L) | MZ229973 (L) | - | |
| LBCE 15997 | Didelphis aurita | T. janseni (B) * | Negative | MZ541906 (B) | - | |
| LBCE 15998 | Didelphis aurita | T. janseni (B) * | Negative | MZ541913 (B) | - | |
| LBCE 17670 | Didelphis aurita | Negative | T. cruzi DTU TcI **** (L) | MZ221942 (L) | - | |
| LBCE 17677 | Didelphis aurita | T. cruzi DTU TcI (B) | Negative | MZ541920 (B) | C00495 | |
| LBCE 17680 | Didelphis aurita | T. janseni (B, Sp) | Negative | MZ541926 (B)/MZ541914 (Sp) | C00494 (B) | |
| LBCE 17683 | Didelphis aurita | T. cruzi DTU TcI (B) | Negative | MZ541921 (B) | C00501 | |
| LBCE 17733 | Didelphis aurita | T. janseni (B) * | Negative | MZ541910 (B) | - | |
| LBCE 17735 | Didelphis aurita | T. cruzi DTU TcI (B) | Negative | MZ541900 (B) | C00538 | |
| LBCE 17736 | Akodon cursor | T. janseni (B) * | Negative | MZ541911 (B) | - | |
| LBCE 17737 | Didelphis aurita | T. janseni (B) * | Trypanosomatidae (Sp) | MZ541909 (B) | - | |
| LBCE 17739 | Marmosa paraguayana | T. cruzi DTU TcIV (B) | Negative | MZ541899 (B) | C00561 | |
| LBCE 17742 | Didelphis aurita | T. cruzi DTU TcI (B) | Negative | MZ541923 (B) | C00564 | |
| LBCE 18203 | Didelphis aurita | T. cruzi DTU TcI (B) | Negative | MZ541922 (B) | C00521 | |
| LBCE 18233 | Didelphis aurita | T. rangeli A (B) * | Negative | MZ541903 (B) | - | |
| LBCE 18241 | Didelphis aurita | T. janseni (B) * | Negative | MZ541925 (B) | - | |
| LBCE 18243 | Didelphis aurita | T. janseni (Sp) | Negative | MZ541916 (B) | C00911 | |
| LBCE 18245 | Didelphis aurita | T. janseni (B) * | Negative | MZ541912 (B) | - | |
| A2 | LBCE 15989 | Didelphis aurita | T. cruzi DTU TcI (B) * | Negative | MZ541904 (B) | - |
| LBCE 15990 | Didelphis aurita | T. cruzi DTU TcI (B) * | Negative | MZ541905 (B) | - | |
| LBCE 15999 | Metachirus myosurus | Negative | Trypanosomatidae (Sp, S) | - | - | |
| LBCE 16000 | Didelphis aurita | T. cruzi DTU TcI (B) * | T. cruzi DTU TcI **** (Sp, S) | MZ541907(B)/MZ221937 (Sp)/MZ221941 (S) | - | |
| LBCE 17665 | Didelphis aurita | T. janseni (Sp, L) ** | Negative | KY243025 (Sp)/KY243026 (L) | - | |
| LBCE 17666 | Didelphis aurita | Negative | T. cruzi DTU TcI (Sp) | MZ221938 (Sp) | - | |
| LBCE 17675 | Didelphis aurita | Negative | Trypanosomatidae (S) | - | - | |
| LBCE 17729 | Didelphis aurita | T. cruzi DTU TcI (B) * | Negative | -2 | - | |
| LBCE 17740 | Didelphis aurita | T. janseni (Sp) | Negative | MZ541915 (B) | - | |
| LBCE 17741 | Didelphis aurita | T. dionisii (B) * | Negative | MZ541908 (B) | - | |
| LBCE 17823 | Didelphis aurita | T. cruzi DTU TcI (B) | Negative | MZ541919 (B) | C00493 | |
| LBCE 18209 1 | Didelphis aurita | T. cruzi DTU TcIV (B) * | Negative | MZ541901 (B) | - | |
| LBCE 18210 | Didelphis aurita | T. cruzi DTU TcI (B) | Negative | MZ541902 (B) | C00520 | |
| LBCE 18251 | Didelphis aurita | T. janseni (Sp) * | Negative | MZ541917 (Sp) | - | |
| LBCE 18255 | Didelphis aurita | T. janseni (B) * | Negative | MZ541918 (B) | - | |
| A3 | LBCE 17678 | Marmosa paraguayana | Negative | Trypanosomatidae (Sp,S) | - | - |
| LBCE 17679 | Marmosa paraguayana | Negative | T. cruzi DTU TcI (Sp)/Trypanosomatidae (S) | MZ221939 (Sp) | - | |
| LBCE 17685 | Marmosa paraguayana | Negative | T. cruzi DTU TcI (Sp)/Trypanosomatidae (L) | MZ221940 (Sp) | - | |
| LBCE 17743 | Didelphis aurita | T. cruzi DTU TcI (B) | Negative | MZ541924 (B) | C00565 | |
| A1 = 20 A2 = 15 A3 = 4 | N = 39 | 5 species | T. cruzi DTU TcI (12) T. cruzi DTU TcIV (2) T. janseni (15) T. dionisii (1) T. rangeli A (1) | Trypanosomatidae (9) T. cruzi DTU TcI (9) | Sequences (39) | Isolates (11) |
| Infected Species (n; %) | T.cruzi (n; %) IFAT Titer Range | Leishmania spp. (n; %) IFAT Titer Range | Mixed Infection * n (%) |
|---|---|---|---|
| Akodon cursor (1; 14.3%) | (1; 100%) 1/10 | (1; 100%) 1/20 | 1 (100%) |
| Rattus rattus (7; 100%) | (5; 71.4%) 1/10–1/40 | (5; 71.4%) 1/10–1/20 | 3 (42.8%) |
| Didelphis aurita (42; 60%) | (29; 69.4%) 1/40–1/160 | (31; 73.8%) 1/40–1/160 | 18 (42.8%) |
| Marmosa paraguayana (3; 75%) | (3; 100%) 1/40–1/160 | (1; 33.3%) 1/80 | 1 (33.3%) |
| 53/88 (60.2%) | 38/53 (71.6%) | 38/53 (71.6%) | 23/53 (43.3%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berbigier, A.P.; Barros, J.H.d.S.; Pontes, E.S.; Lisboa, C.V.; Gentile, R.; Xavier, S.C.d.C.; Jansen, A.M.; Roque, A.L.R. Trypanosomatid Richness in Wild and Synanthropic Small Mammals from a Biological Station in Rio de Janeiro, Brazil. Pathogens 2021, 10, 1442. https://doi.org/10.3390/pathogens10111442
Berbigier AP, Barros JHdS, Pontes ES, Lisboa CV, Gentile R, Xavier SCdC, Jansen AM, Roque ALR. Trypanosomatid Richness in Wild and Synanthropic Small Mammals from a Biological Station in Rio de Janeiro, Brazil. Pathogens. 2021; 10(11):1442. https://doi.org/10.3390/pathogens10111442
Chicago/Turabian StyleBerbigier, Alice Pereira, Juliana Helena da Silva Barros, Edilene Sousa Pontes, Cristiane Varella Lisboa, Rosana Gentile, Samanta Cristina das Chagas Xavier, Ana Maria Jansen, and André Luiz Rodrigues Roque. 2021. "Trypanosomatid Richness in Wild and Synanthropic Small Mammals from a Biological Station in Rio de Janeiro, Brazil" Pathogens 10, no. 11: 1442. https://doi.org/10.3390/pathogens10111442
APA StyleBerbigier, A. P., Barros, J. H. d. S., Pontes, E. S., Lisboa, C. V., Gentile, R., Xavier, S. C. d. C., Jansen, A. M., & Roque, A. L. R. (2021). Trypanosomatid Richness in Wild and Synanthropic Small Mammals from a Biological Station in Rio de Janeiro, Brazil. Pathogens, 10(11), 1442. https://doi.org/10.3390/pathogens10111442