
Mycobacterium avium Subsp. hominissuis Interactions with Macrophage Killing Mechanisms



Abstract
:1. Introduction
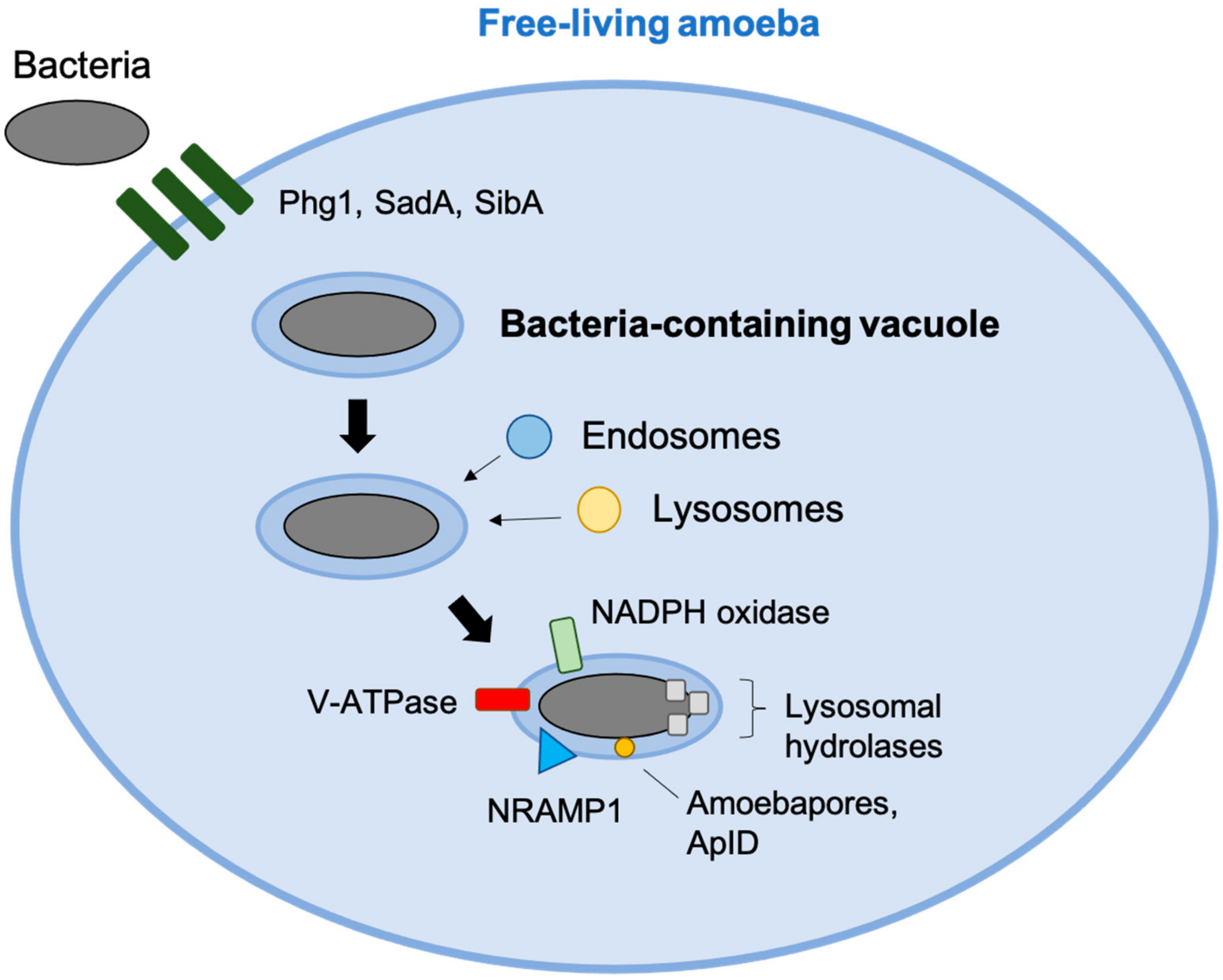
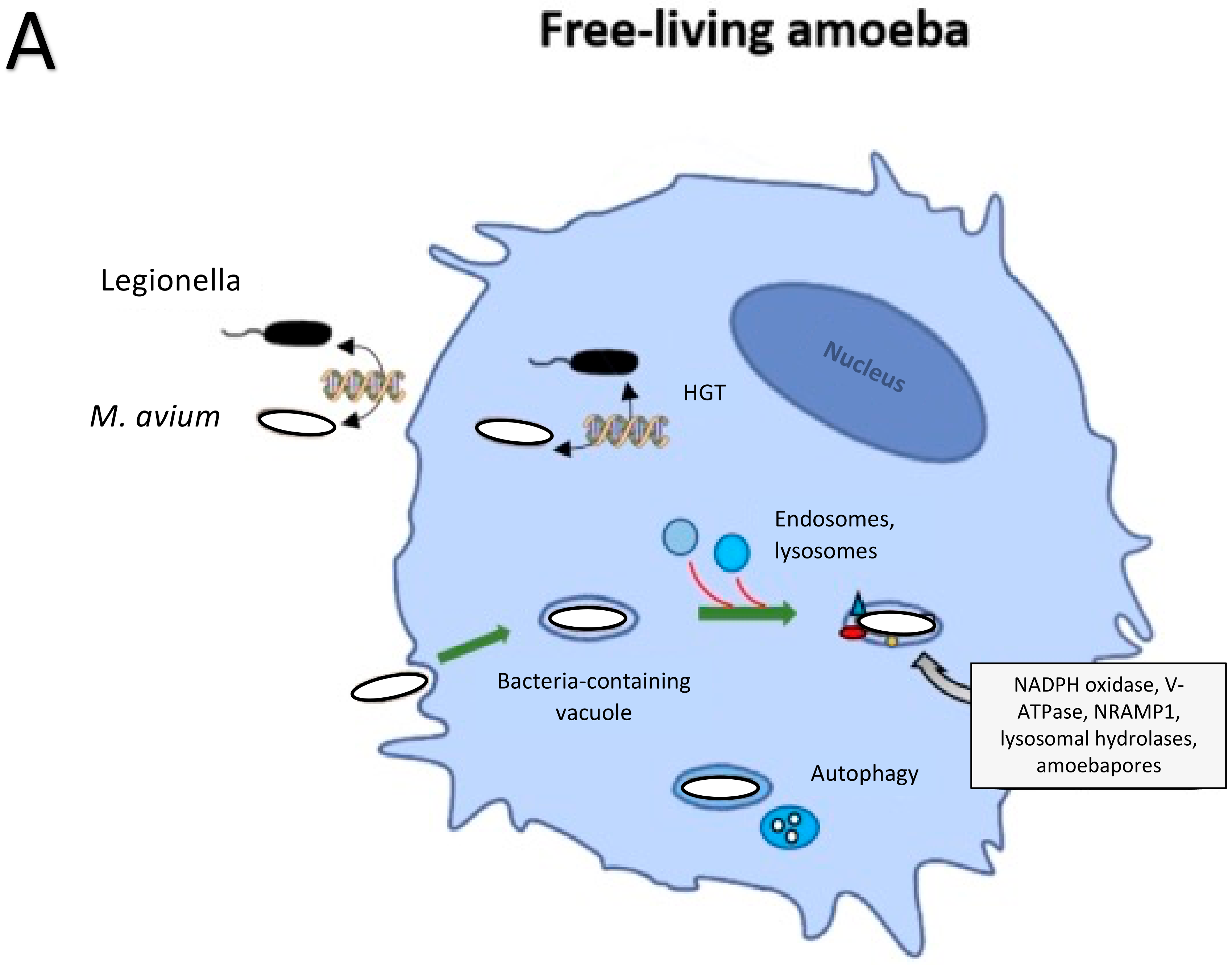
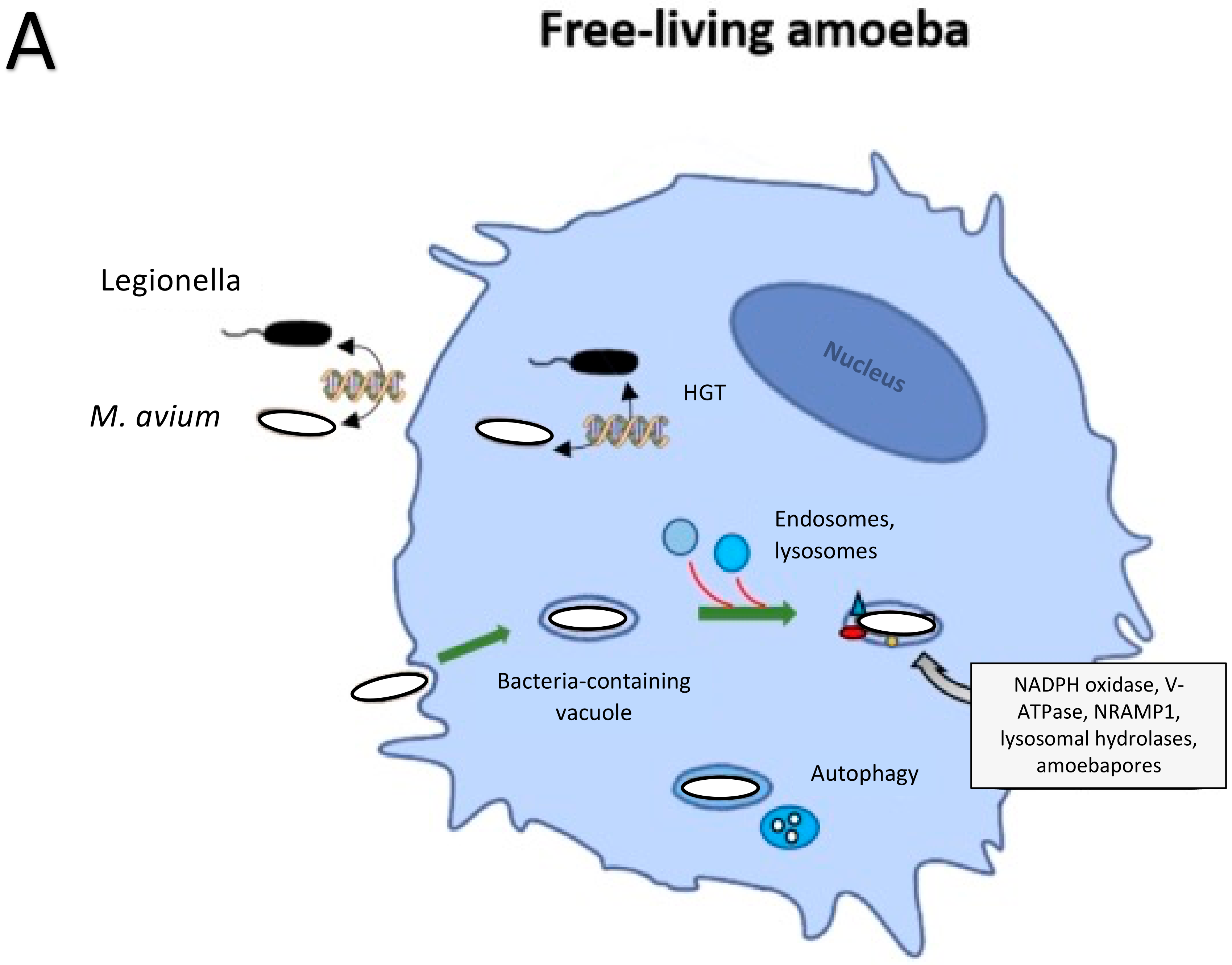
2. Bacteria Evolution in Protozoa and Other Inhabitants of the Soil
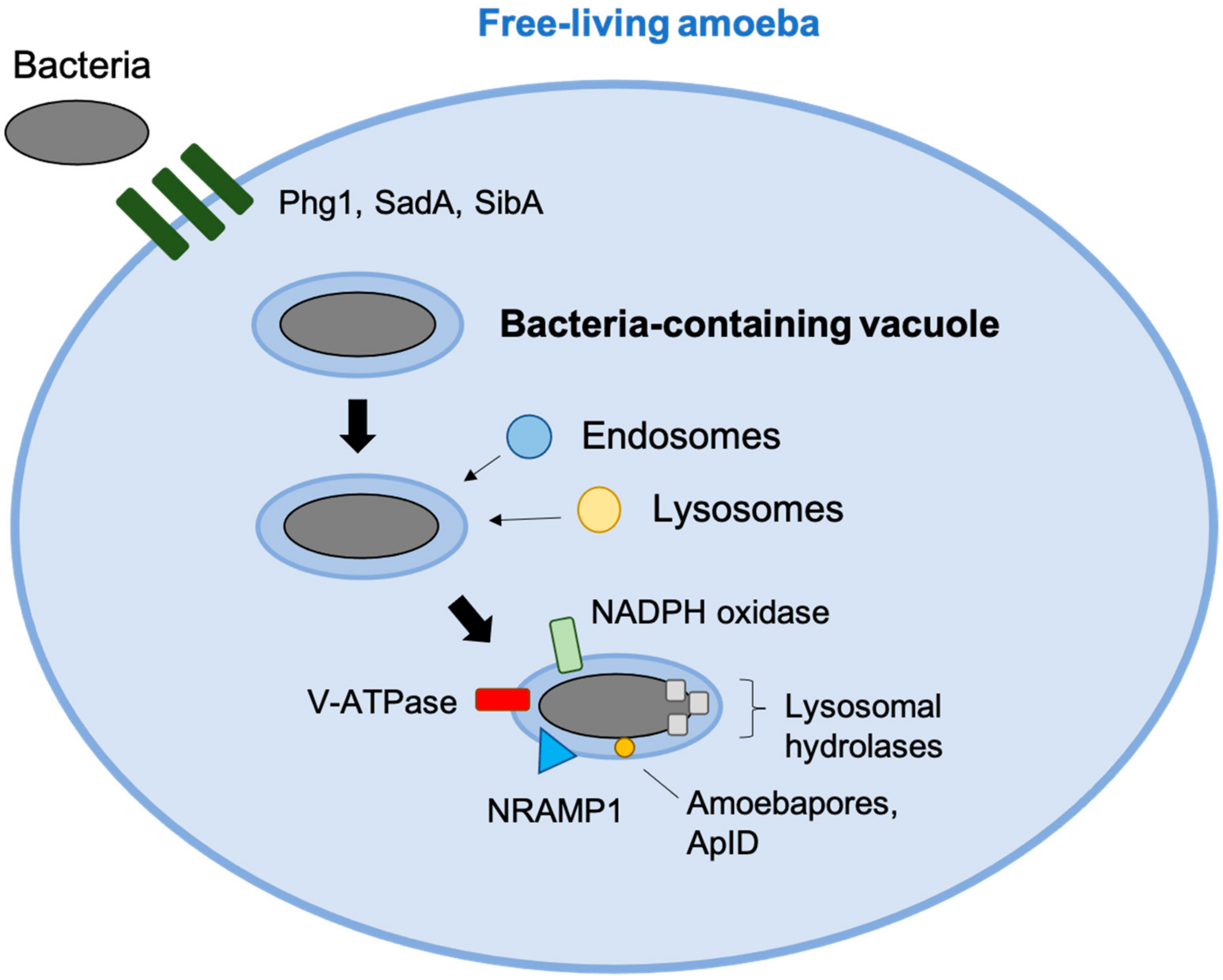
3. Environmental Protozoans Killing of Bacteria
4. M. avium and the Phagosome Environment
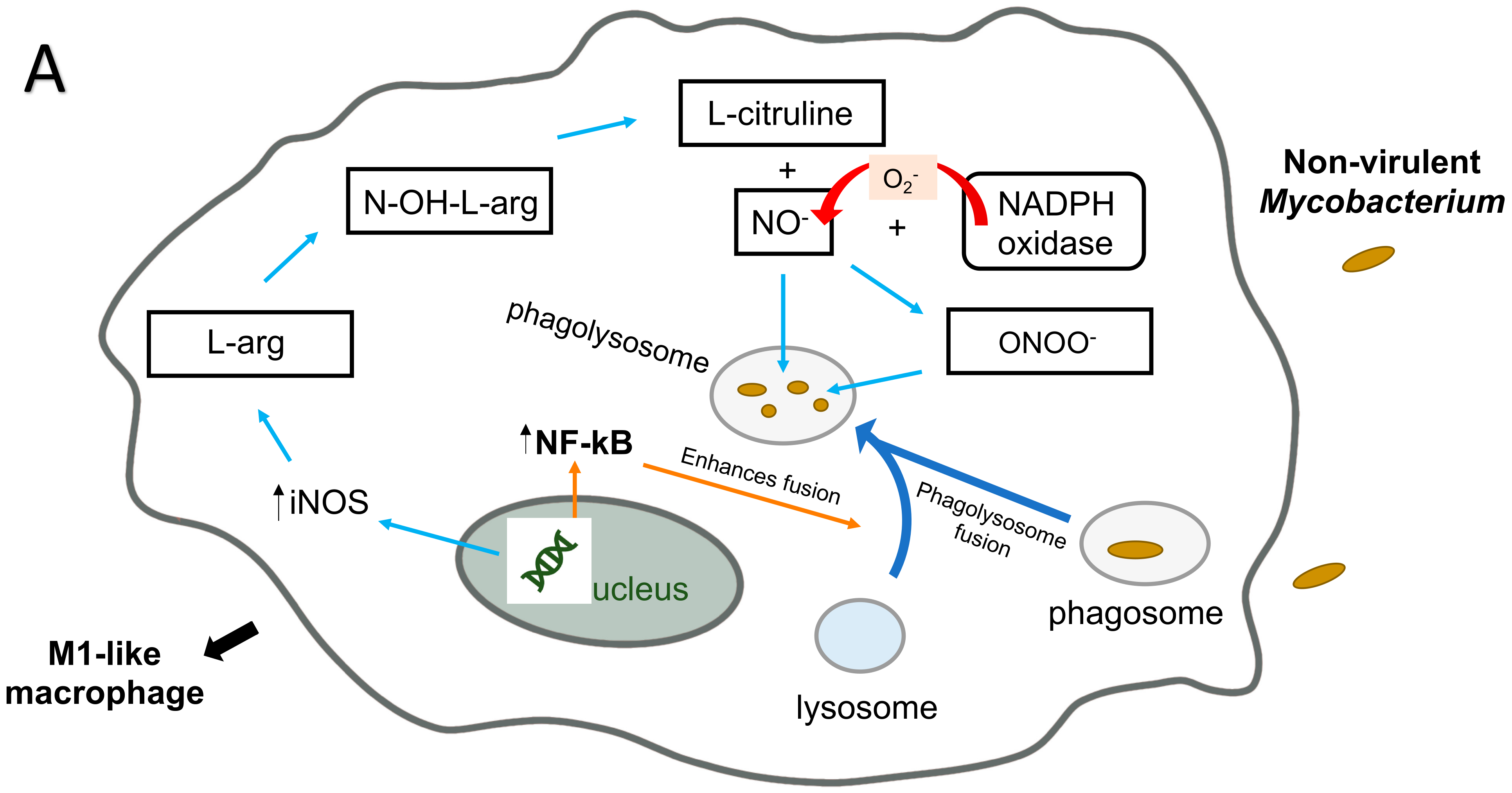
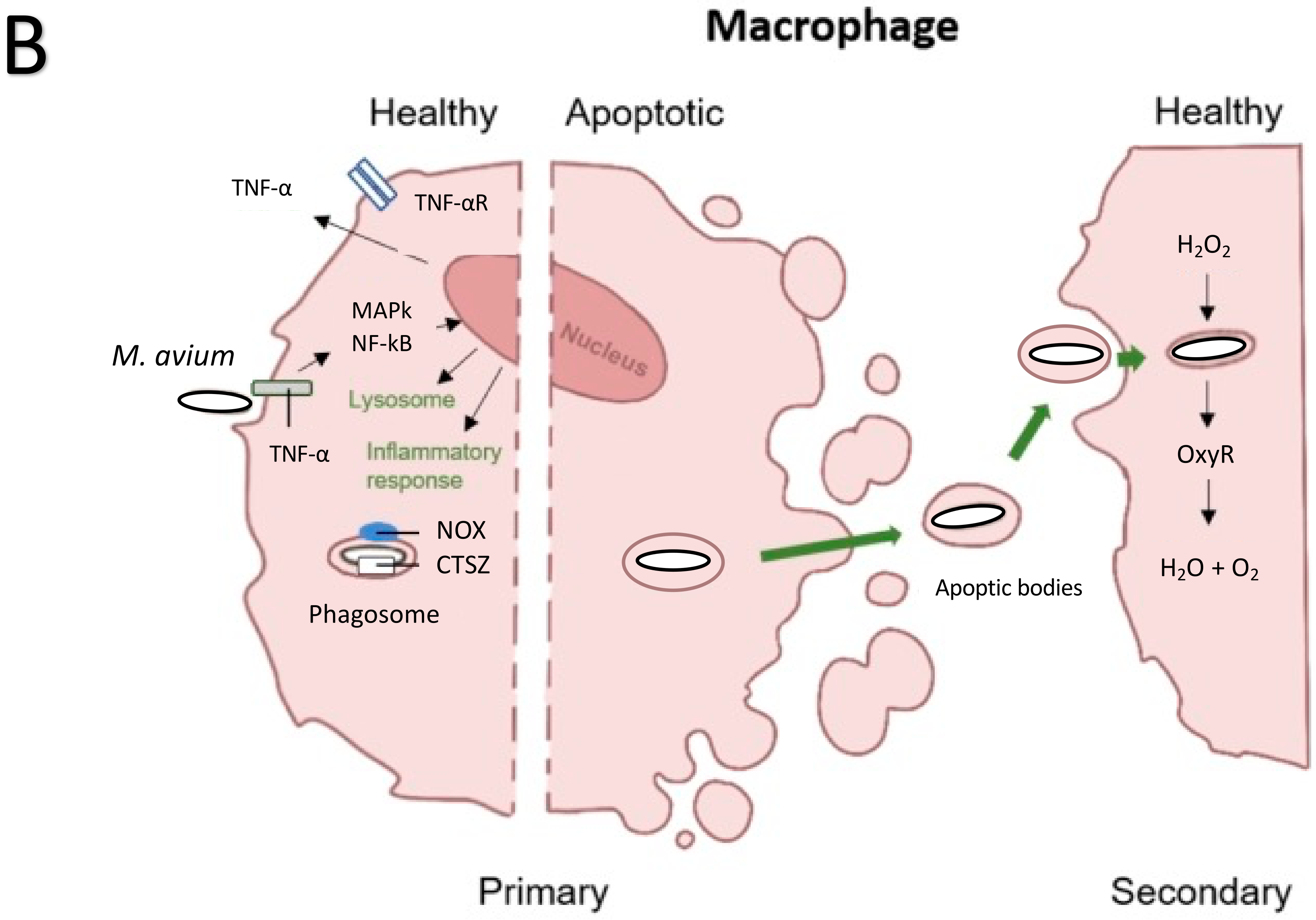
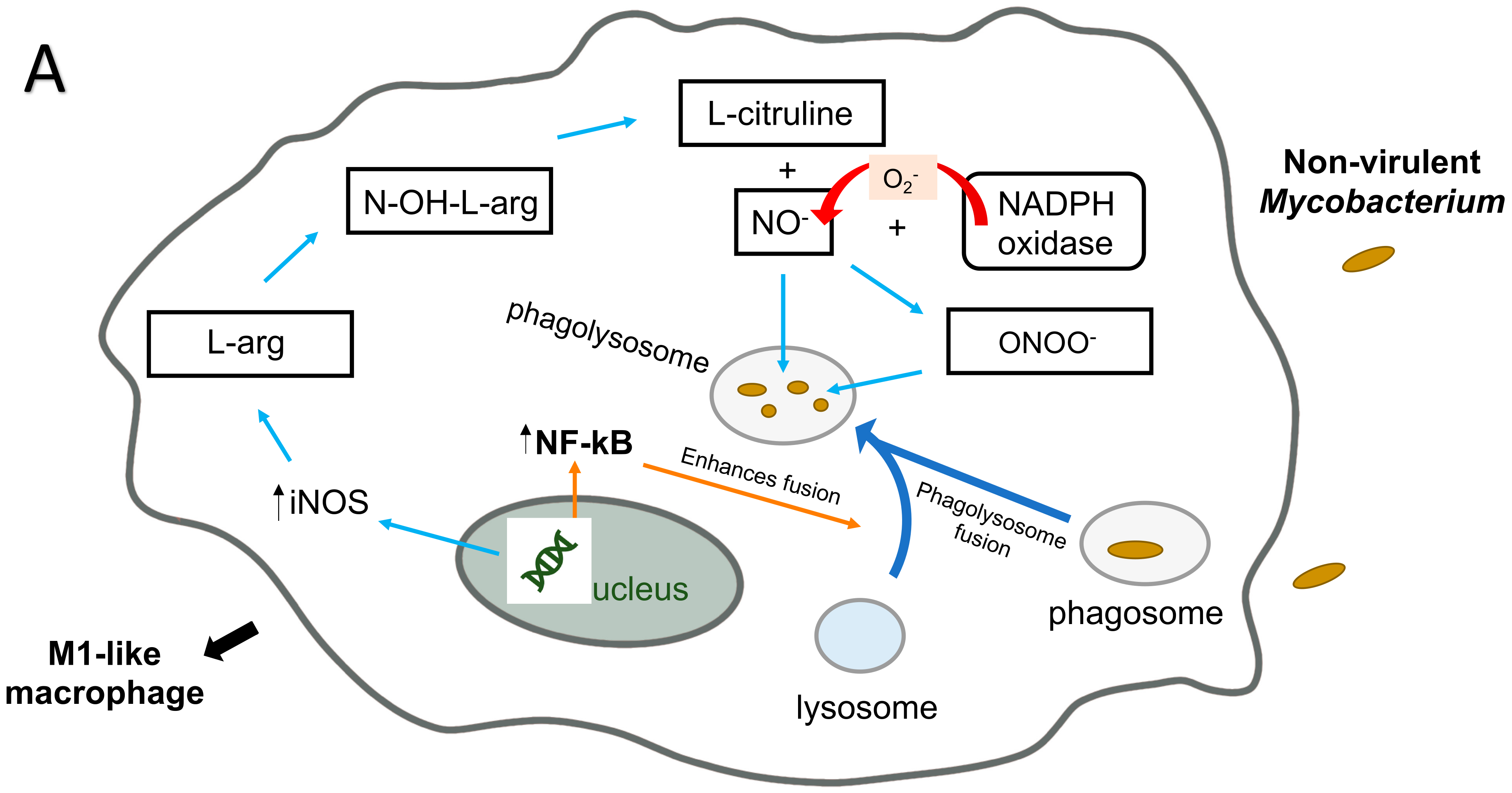
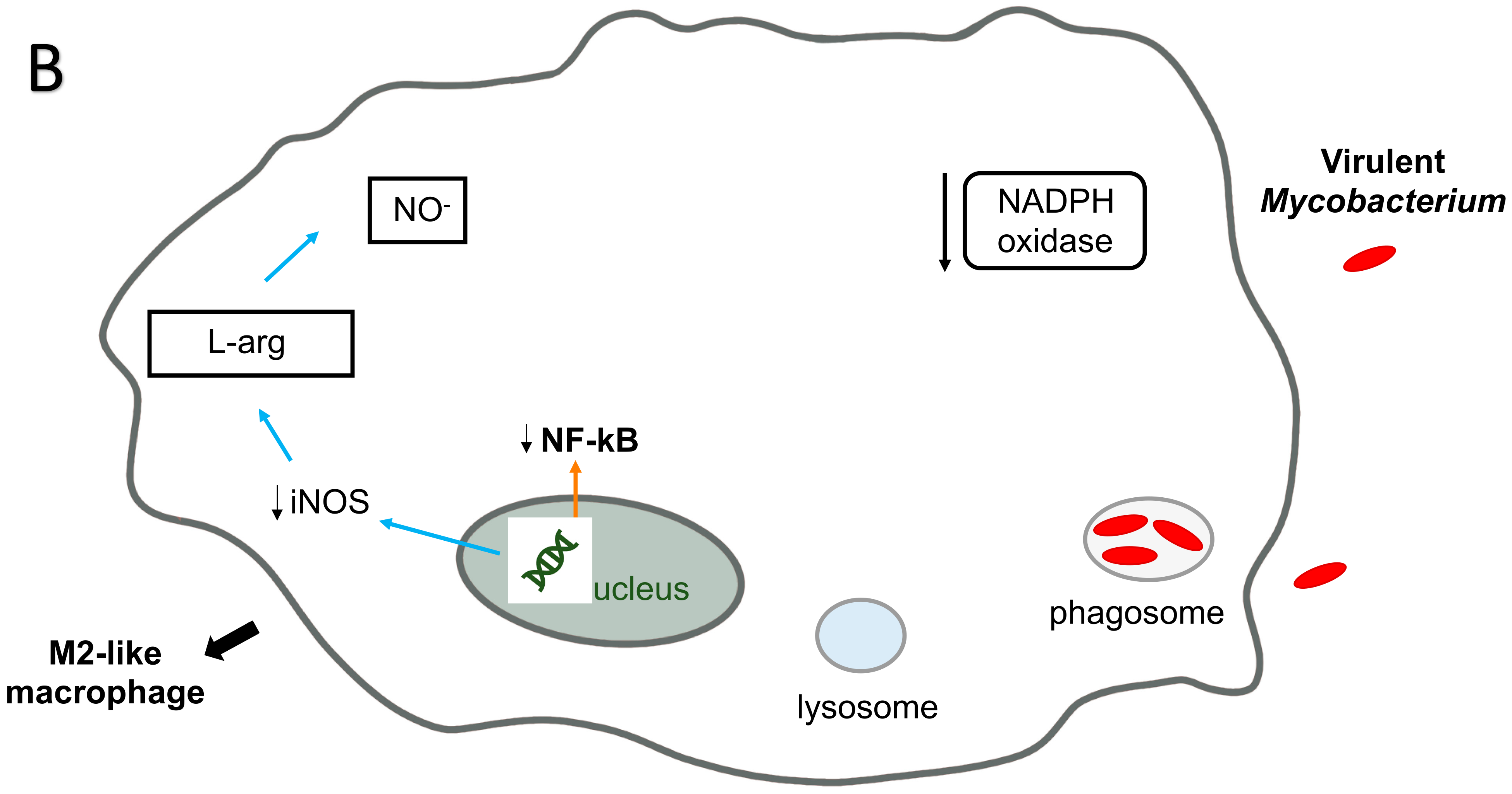
5. M. avium Infection of Macrophages: ROS Killing Mechanisms
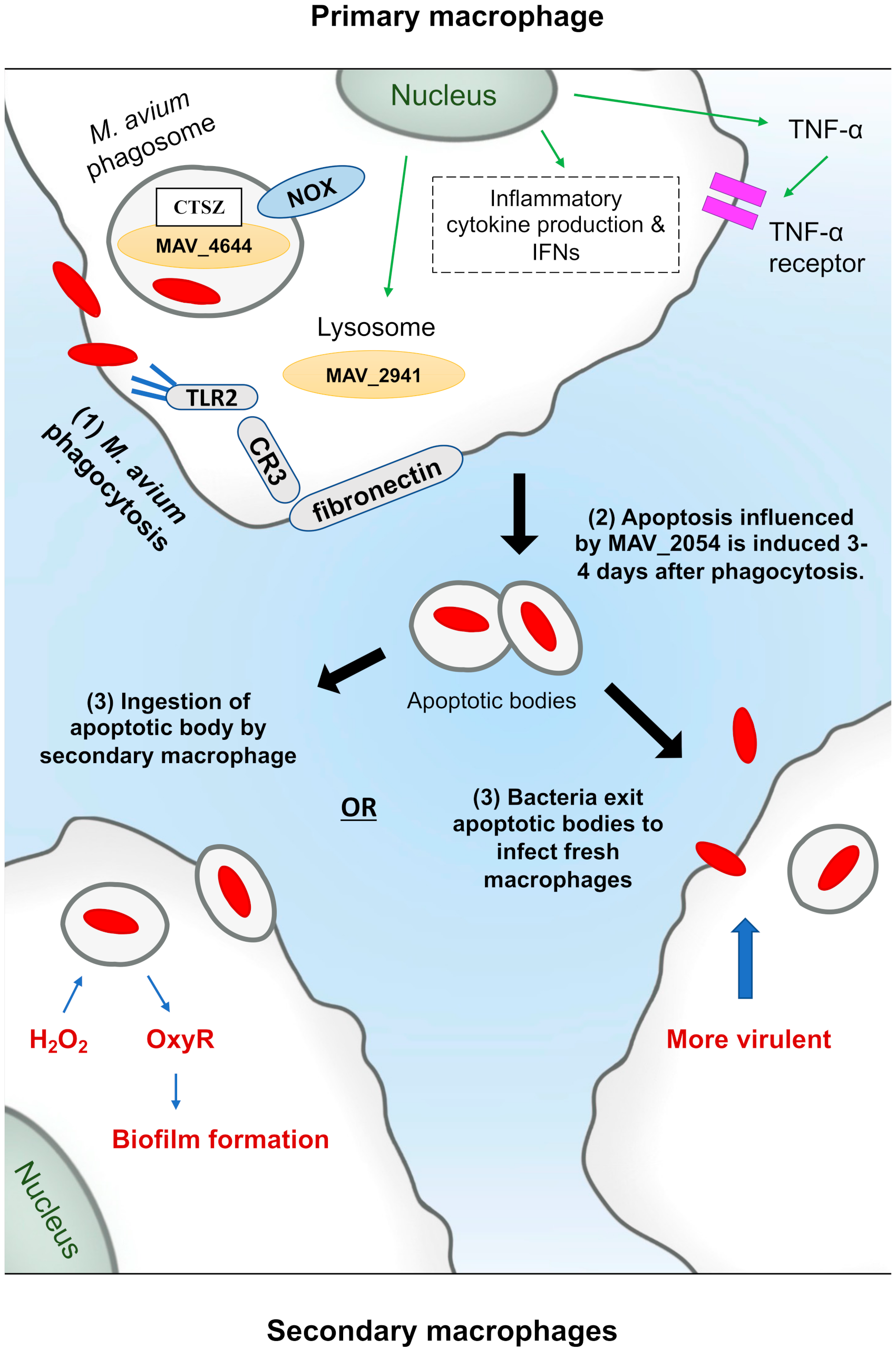
6. M. avium and Macrophages: RNI Killing Mechanism
M. avium and Methylation of DNA in Macrophages
7. Using the Current Understanding to Benefit Treatment
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Beer, V.; Bonifas, V. Mycobacteria. Schweiz. Med. Wochenschr. 1975, 105, 984–987. [Google Scholar]
- Thomas, V.; McDonnell, G. Relationship between mycobacteria and amoebae: Ecological and epidemiological concerns. Lett. Appl. Microbiol. 2007, 45, 349–357. [Google Scholar] [CrossRef]
- Drancourt, M. Looking in amoebae as a source of mycobacteria. Microb. Pathog. 2014, 77, 119–124. [Google Scholar] [CrossRef]
- Busatto, C.; Vianna, J.S.; da Silva, L.V.; Ramis, B.R.; da Silva, P.E.A. Mycobacterium avium: An overview. Tuberculosis 2019, 114, 127–134. [Google Scholar] [CrossRef]
- Mougari, F.; Guglielmetti, L.; Raskine, L.; Sermet-Gaudelus, I.; Veziris, N.; Cambau, E. Infections caused by Mycobacterium abscessus: Epidemiology, diagnostic tools and treatment. Expert Rev. Anti-Infect. Ther. 2016, 14, 1139–1154. [Google Scholar] [CrossRef]
- Griffith, D.E.; Aksamit, T.; Brown-Elliott, B.A.; Daley, G.; Gordin, F.; Holland, S.M.; Horsburgh, R.; Huitt, G.; Iademarco, M.F.; Iseman, M.; et al. An official ATS/IDSA statement: Diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. Am. J. Respir Crit. Care Med. 2007, 175, 367–416. [Google Scholar] [CrossRef]
- Henkle, E.; Winthrop, K.L. Nontuberculous mycobacteria infections in immunosuppressed hosts. Clin. Chest Med. 2015, 36, 91–99. [Google Scholar] [CrossRef] [Green Version]
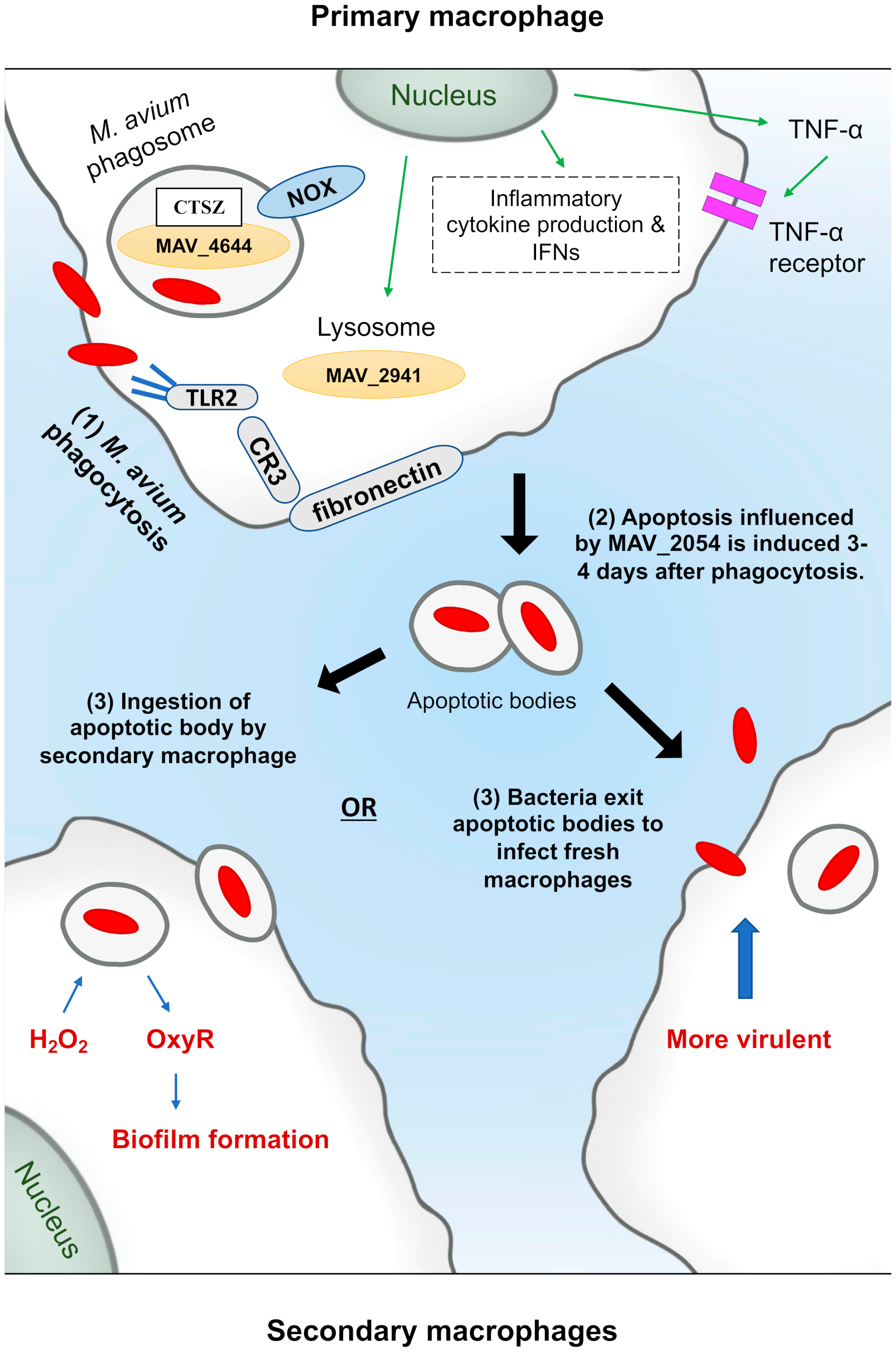
- Danelishvili, L.; Bermudez, L.E. Mycobacterium avium MAV_2941 mimics phosphoinositol-3-kinase to interfere with macrophage phagosome maturation. Microbes Infect. 2015, 17, 628–637. [Google Scholar] [CrossRef] [Green Version]
- Lewis, M.S.; Danelishvili, L.; Rose, S.J.; Bermudez, L.E. MAV_4644 Interaction with the Host Cathepsin Z Protects Mycobacterium avium subsp. hominissuis from Rapid Macrophage Killing. Microorganisms 2019, 7, 144. [Google Scholar] [CrossRef] [Green Version]
- Greub, G.; Raoult, D. Microorganisms Resistant to Free-Living Amoebae. Clin. Microbiol. Rev. 2004, 17, 413–433. [Google Scholar] [CrossRef] [Green Version]
- Danelishvili, L.; Wu, M.; Stang, B.; Harriff, M.; Cirillo, S.L.; Cirillo, J.D.; Bildfell, R.; Arbogast, B.; Bermudez, L.E. Identification of Mycobacterium avium pathogenicity island important for macrophage and amoeba infection. Proc. Natl. Acad. Sci. USA 2007, 104, 11038–11043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirillo, J.D.; Falkow, S.; Tompkins, L.S.; Bermudez, L.E. Interaction of Mycobacterium avium with environmental amoebae enhances virulence. Infect. Immun. 1997, 65, 3759–3767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermudez, L.E.; Rose, S.; Everman, J.; Ziaie, N. Establishment of a Mycobacterium avium subsp. hominissuis host-to-host transmission model in Caenorhabditis elegans and bacterial colonization associated genes. Front. Cell. Infect. Microbiol. 2018, 8, 123. [Google Scholar] [CrossRef] [PubMed]
- Samba-Louaka, A.; Delafont, V.; Rodier, M.H.; Cateau, E.; Héchard, Y. Free-living amoebae and squatters in the wild: Ecological and molecular features. FEMS Microbiol. Rev. 2019, 43, 415–434. [Google Scholar] [CrossRef] [PubMed]
- Wheat, W.H.; Casali, A.L.; Thomas, V.; Spencer, J.S.; Lahiri, R.; Williams, D.L.; McDonnell, G.E.; Gonzalez-Juarrero, M.; Brennan, P.J.; Jackson, M. Long-term survival and virulence of Mycobacterium leprae in amoebal cysts. PLoS Negl. Trop. Dis. 2014, 8, e3405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, H.; Suzuki, H.; Maruyama, F.; Iwamoto, T. The recombination-cold region as an epidemiological marker of recombinogenic opportunistic pathogen Mycobacterium avium. BMC Genom. 2019, 20, 752. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.; Shin, S.J. Genetic Involvement of Mycobacterium avium Complex in the Regulation and Manipulation of Innate Immune Functions of Host Cells. Int. J. Mol. Sci. 2021, 22, 3011. [Google Scholar] [CrossRef] [PubMed]
- Bannantine, J.P.; Zhang, Q.; Li, L.L.; Kapur, V. Genomic homogeneity between Mycobacterium avium subsp. avium and Mycobacterium avium subsp. paratuberculosis belies their divergent growth rates. BMC Microbiol. 2003, 3, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strassmann, J.E.; Shu, L. Ancient bacteria-amoeba relationships and pathogenic animal bacteria. PLoS Biol. 2017, 15, e2002460. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Edwards, M.; Falkinham, J.O.; Pruden, A. Molecular survey of the occurrence of Legionella spp., Mycobacterium spp., Pseudomonas aeruginosa, and amoeba hosts in two chloraminated drinking water distribution systems. Appl. Environ. Microbiol. 2012, 78, 6285–6294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiley, H.; Keegan, A.; Fallowfield, H.; Bentham, R. Detection of Legionella, L. pneumophila and Mycobacterium avium complex (MAC) along potable water distribution pipelines. Int. J. Environ. Res. Public Health 2014, 11, 7393–7405. [Google Scholar] [CrossRef] [Green Version]
- Whiley, H.; Giglio, S.; Bentham, R. Opportunistic Pathogens Mycobacterium Avium Complex (MAC) and Legionella spp. Colonise Model Shower. Pathogens 2015, 4, 590–598. [Google Scholar] [CrossRef]
- Steinert, M.; Birkness, K.; White, E.; Fields, B.; Quinn, F. Mycobacterium avium bacilli grow saprozoically in coculture with Acanthamoeba polyphaga and survive within cyst walls. Appl. Environ. Microbiol. 1998, 64, 2256–2261. [Google Scholar] [CrossRef] [Green Version]
- Lamrabet, O.; Merhej, V.; Pontarotti, P.; Raoult, D.; Drancourt, M. The genealogic tree of mycobacteria reveals a long-standing sympatric life into free-living protozoa. PLoS ONE 2012, 7, e34754. [Google Scholar] [CrossRef]
- Cosson, P.; Soldati, T. Eat, kill or die: When amoeba meets bacteria. Curr. Opin. Microbiol. 2008, 11, 271–276. [Google Scholar] [CrossRef]
- Gotthardt, D.; Warnatz, H.J.; Henschel, O.; Brückert, F.; Schleicher, M.; Soldati, T. High-resolution dissection of phagosome maturation reveals distinct membrane trafficking phases. Mol. Biol. Cell 2002, 13, 3508–3520. [Google Scholar] [CrossRef] [Green Version]
- German, N.; Doyscher, D.; Rensing, C. Bacterial killing in macrophages and amoeba: Do they all use a brass dagger. Future Microbiol. 2013, 8, 1257–1264. [Google Scholar] [CrossRef]
- Benghezal, M.; Fauvarque, M.O.; Tournebize, R.; Froquet, R.; Marchetti, A.; Bergeret, E.; Lardy, B.; Klein, G.; Sansonetti, P.; Charette, S.J.; et al. Specific host genes required for the killing of Klebsiella bacteria by phagocytes. Cell Microbiol. 2006, 8, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Andrä, J.; Herbst, R.; Leippe, M. Amoebapores, archaic effector peptides of protozoan origin, are discharged into phagosomes and kill bacteria by permeabilizing their membranes. Dev. Comp. Immunol. 2003, 27, 291–304. [Google Scholar] [CrossRef]
- Leippe, M.; Andrä, J.; Müller-Eberhard, H.J. Cytolytic and antibacterial activity of synthetic peptides derived from amoebapore, the pore-forming peptide of Entamoeba histolytica. Proc. Natl. Acad. Sci. USA 1994, 91, 2602–2606. [Google Scholar] [CrossRef] [Green Version]
- Leippe, M.; Andrä, J.; Nickel, R.; Tannich, E.; Müller-Eberhard, H.J. Amoebapores, a family of membranolytic peptides from cytoplasmic granules of Entamoeba histolytica: Isolation, primary structure, and pore formation in bacterial cytoplasmic membranes. Mol. Microbiol. 1994, 14, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Dhakshinamoorthy, R.; Bitzhenner, M.; Cosson, P.; Soldati, T.; Leippe, M. The Saposin-Like Protein AplD Displays Pore-Forming Activity and Participates in Defense Against Bacterial Infection During a Multicellular Stage of Dictyostelium discoideum. Front. Cell. Infect. Microbiol. 2018, 8, 73. [Google Scholar] [CrossRef]
- Alderwick, L.J.; Harrison, J.; Lloyd, G.S.; Birch, H.L. The Mycobacterial Cell Wall-Peptidoglycan and Arabinogalactan. Cold Spring Harb. Perspect. Med. 2015, 5, a021113. [Google Scholar] [CrossRef] [Green Version]
- Clarke, M.; Lohan, A.J.; Liu, B.; Lagkouvardos, I.; Roy, S.; Zafar, N.; Bertelli, C.; Schilde, C.; Kianianmomeni, A.; Bürglin, T.R.; et al. Genome of Acanthamoeba castellanii highlights extensive lateral gene transfer and early evolution of tyrosine kinase signaling. Genome Biol. 2013, 14, R11. [Google Scholar] [CrossRef] [Green Version]
- Peracino, B.; Balest, A.; Bozzaro, S. Phosphoinositides differentially regulate bacterial uptake and Nramp1-induced resistance to Legionella infection in Dictyostelium. J. Cell Sci. 2010, 123 Pt 23, 4039–5401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motamedi, N.; Danelishvili, L.; Bermudez, L.E. Identification of Mycobacterium avium genes associated with resistance to host antimicrobial peptides. J. Med. Microbiol. 2014, 63 Pt 7, 923–930. [Google Scholar] [CrossRef]
- Allen, M.; Bailey, C.; Cahatol, I.; Dodge, L.; Yim, J.; Kassissa, C.; Luong, J.; Kasko, S.; Pandya, S.; Venketaraman, V. Mechanisms of Control of Mycobacterium tuberculosis by NK Cells: Role of Glutathione. Front. Immunol. 2015, 6, 508. [Google Scholar] [CrossRef] [Green Version]
- Mesquita, A.; Cardenal-Muñoz, E.; Dominguez, E.; Muñoz-Braceras, S.; Nuñez-Corcuera, B.; Phillips, B.A.; Tábara, L.C.; Xiong, Q.; Coria, R.; Eichinger, L.; et al. Autophagy in Dictyostelium: Mechanisms, regulation and disease in a simple biomedical model. Autophagy 2017, 13, 24–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhary, A.; Miller, S.I. Xenophagy: Pathogen-Containing Vacuoles Are Hard to Digest. Curr. Biol. 2019, 29, R1086–R1088. [Google Scholar] [CrossRef]
- Fais, S.; Overholtzer, M. Cell-in-cell phenomena, cannibalism, and autophagy: Is there a relationship. Cell Death Dis. 2018, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Rolando, M.; Escoll, P.; Nora, T.; Botti, J.; Boitez, V.; Bedia, C.; Daniels, C.; Abraham, G.; Stogios, P.J.; Skarina, T.; et al. Legionella pneumophila S1P-lyase targets host sphingolipid metabolism and restrains autophagy. Proc. Natl. Acad. Sci. USA 2016, 113, 1901–1906. [Google Scholar] [CrossRef] [Green Version]
- Franco, L.H.; Vidhya, R.N.; Scharn, C.R.; Xavier, R.J.; Torrealba, J.R.; Shiloh, M.U.; Levine, B. The ubiquitin ligase smurf1 functions in selective autophagy of Mycobacterium tuberculosis and anti-tuberculosis host defense. Cell Host Microbe 2017, 21, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, T.H.; Hsiao, Y.S.; Jo, J.; Chou, C.Y.; Dietrich, L.E.; Walz, T.; Tong, L. Structure and function of a single-chain, multi-domain long-chain acyl-CoA carboxylase. Nature 2015, 518, 120–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ignatov, D.; Kondratieva, E.; Azhikina, T.; Apt, A. Mycobacterium avium-triggered diseases: Pathogenomics. Cell. Microbiol. 2012, 14, 808–818. [Google Scholar] [CrossRef] [PubMed]
- Parker, H.; Dragunow, M.; Hampton, M.B.; Kettle, A.J.; Winterbourn, C.C. Requirements for NADPH oxidase and myeloperoxidase in neutrophil extracellular trap formation differ depending on the stimulus. J. Leukoc. Biol. 2012, 92, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Hampson, S.J.; Portaels, F.; Thompson, J.; Green, E.P.; Moss, M.T.; Hermon-Taylor, J.; McFadden, J.J. DNA probes demonstrate a single highly conserved strain of Mycobacterium avium infecting AIDS patients. Lancet 1989, 1, 65–68. [Google Scholar] [CrossRef]
- de Chastellier, C.; Thilo, L. Pathogenic Mycobacterium avium remodels the phagosome membrane in macrophages within days after infection. Eur. J. Cell Biol. 2002, 81, 17–25. [Google Scholar] [CrossRef]
- Schaible, U.E.; Sturgill-Koszycki, S.; Schlesinger, P.H.; Russell, D.G. Cytokine activation leads to acidification and increases maturation of Mycobacterium avium-containing phagosomes in murine macrophages. J. Immunol. 1998, 160, 1290–1296. [Google Scholar]
- Robinson, N.; Wolke, M.; Ernestus, K.; Plum, G. A mycobacterial gene involved in synthesis of an outer cell envelope lipid is a key factor in prevention of phagosome maturation. Infect. Immun. 2007, 75, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Kissing, S.; Hermsen, C.; Repnik, U.; Nesset, C.K.; von Bargen, K.; Griffiths, G.; Ichihara, A.; Lee, B.S.; Schwake, M.; De Brabander, J.; et al. Vacuolar ATPase in phagosome-lysosome fusion. J. Biol. Chem. 2015, 290, 14166–14180. [Google Scholar] [CrossRef] [Green Version]
- Early, J.; Fischer, K.; Bermudez, L.E. Mycobacterium avium uses apoptotic macrophages as tools for spreading. Microb. Pathog. 2011, 50, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Miltner, E.; Wu, M.; Petrofsky, M.; Bermudez, L.E. A Mycobacterium avium PPE gene is associated with the ability of the bacterium to grow in macrophages and virulence in mice. Cell. Microbiol. 2005, 7, 539–548. [Google Scholar] [CrossRef]
- Danelishvili, L.; Poort, M.J.; Bermudez, L.E. Identification of Mycobacterium avium genes up-regulated in cultured macrophages and in mice. FEMS Microbiol. Lett. 2004, 239, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Jha, S.S.; Danelishvili, L.; Wagner, D.; Maser, J.; Li, Y.J.; Moric, I.; Vogt, S.; Yamazaki, Y.; Lai, B.; Bermudez, L.E. Virulence-related Mycobacterium avium subsp hominissuis MAV_2928 gene is associated with vacuole remodeling in macrophages. BMC Microbiol. 2010, 10, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinison, J.J.; Danelishvili, L.; Gupta, R.; Rose, S.J.; Babrak, L.M.; Bermudez, L.E. Identification of Mycobacterium avium subsp. hominissuis secreted proteins using an in vitro system mimicking the phagosomal environment. BMC Microbiol. 2016, 16, 270. [Google Scholar] [CrossRef] [Green Version]
- Pfalzer, A.C.; Choi, S.W.; Tammen, S.A.; Park, L.K.; Bottiglieri, T.; Parnell, L.D.; Lamon-Fava, S. S-adenosylmethionine mediates inhibition of inflammatory response and changes in DNA methylation in human macrophages. Physiol. Genom. 2014, 46, 617–623. [Google Scholar] [CrossRef] [PubMed]
- McNamara, M.; Tzeng, S.C.; Maier, C.; Zhang, L.; Bermudez, L.E. Surface proteome of “Mycobacterium avium subsp. hominissuis” during the early stages of macrophage infection. Infect. Immun. 2012, 80, 1868–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNamara, M.; Tzeng, S.C.; Maier, C.; Wu, M.; Bermudez, L.E. Surface-exposed proteins of pathogenic mycobacteria and the role of cu-zn superoxide dismutase in macrophages and neutrophil survival. Proteome Sci. 2013, 11, 45. [Google Scholar] [CrossRef] [Green Version]
- McNamara, M.; Danelishvili, L.; Bermudez, L.E. The Mycobacterium avium ESX-5 PPE protein, PPE25-MAV, interacts with an ESAT-6 family Protein, MAV_2921, and localizes to the bacterial surface. Microb. Pathog. 2012, 52, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Gomes, M.S.; Appelberg, R. Evidence for a link between iron metabolism and Nramp1 gene function in innate resistance against Mycobacterium avium. Immunology 1998, 95, 165–168. [Google Scholar] [CrossRef]
- Gerrick, E.R.; Barbier, T.; Chase, M.R.; Xu, R.; François, J.; Lin, V.H.; Szucs, M.J.; Rock, J.M.; Ahmad, R.; Tjaden, B.; et al. Small RNA profiling in Mycobacterium tuberculosis identifies MrsI as necessary for an anticipatory iron sparing response. Proc. Natl. Acad. Sci. USA 2018, 115, 6464–6469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alter-Koltunoff, M.; Goren, S.; Nousbeck, J.; Feng, C.G.; Sher, A.; Ozato, K.; Azriel, A.; Levi, B.Z. Innate immunity to intraphagosomal pathogens is mediated by interferon regulatory factor 8 (IRF-8) that stimulates the expression of macrophage-specific Nramp1 through antagonizing repression by c-Myc. J. Biol. Chem. 2008, 283, 2724–2733. [Google Scholar] [CrossRef] [Green Version]
- Goswami, T.; Bhattacharjee, A.; Babal, P.; Searle, S.; Moore, E.; Li, M.; Blackwell, J.M. Natural-resistance-associated macrophage protein 1 is an H+/bivalent cation antiporter. Biochem. J. 2001, 354 Pt 3, 511–519. [Google Scholar] [CrossRef]
- Wyllie, S.; Seu, P.; Goss, J.A. The natural resistance-associated macrophage protein 1 Slc11a1 (formerly Nramp1) and iron metabolism in macrophages. Microbes Infect. 2002, 4, 351–359. [Google Scholar] [CrossRef]
- Lee, K.-I.; Choi, H.-G.; Son, Y.-J.; Whang, J.; Kim, K.; Jeon, H.S.; Park, H.-S.; Back, Y.W.; Choi, S.; Kim, S.-W.; et al. Mycobacterium avium MAV2052 protein induces apoptosis in murine macrophage cells through Toll-like receptor 4. Apoptosis 2016, 21, 459–472. [Google Scholar] [CrossRef]
- Danelishvili, L.; Rojony, R.; Carson, K.L.; Palmer, A.L.; Rose, S.J.; Bermudez, L.E. Mycobacterium avium subsp. hominissuis effector MAVA5_06970 promotes rapid apoptosis in secondary-infected macrophages during cell-to-cell spread. Virulence 2018, 9, 1287–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balcewicz-Sablinska, M.K.; Gan, H.; Remold, H.G. Interleukin 10 produced by macrophages inoculated with Mycobacterium avium attenuates mycobacteria-induced apoptosis by reduction of TNF-alpha activity. J. Infect. Dis. 1999, 180, 1230–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, W.J. Nontuberculous Mycobacteria-Overview. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Emmerich, K.; Fabri, M. Nontuberculous mycobacteria. Hautarzt 2017, 68, 403–411. [Google Scholar] [CrossRef]
- Agdestein, A.; Jones, A.; Flatberg, A.; Johansen, T.B.; Heffernan, I.A.; Djønne, B.; Bosco, A.; Olsen, I. Intracellular growth of Mycobacterium avium subspecies and global transcriptional responses in human macrophages after infection. BMC Genom. 2014, 15, 58. [Google Scholar] [CrossRef] [Green Version]
- Bermudez, L.E.; Goodman, J.; Petrofsky, M. Role of complement receptors in uptake of Mycobacterium avium by macrophages in vivo: Evidence from studies using CD18-deficient mice. Infect. Immun. 1999, 67, 4912–4916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermudez, L.E.; Young, L.S.; Enkel, H. Interaction of Mycobacterium avium complex with human macrophages: Roles of membrane receptors and serum proteins. Infect. Immun. 1991, 59, 1697–1702. [Google Scholar] [CrossRef] [Green Version]
- Koh, W.J.; Moon, S.M.; Kim, S.Y.; Woo, M.A.; Kim, S.; Jhun, B.W.; Park, H.Y.; Jeon, K.; Huh, H.J.; Ki, C.S.; et al. Outcomes of Mycobacterium avium complex lung disease based on clinical phenotype. Eur. Respir. J. 2017, 50, 1602503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C. Nitric oxide synthase in innate and adaptive immunity: An update. Trends Immunol. 2015, 36, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C.; Shiloh, M.U. Reactive oxygen and nitrogen intermediates in the relationship between mammalian hosts and microbial pathogens. Proc. Natl. Acad. Sci. USA 2000, 97, 8841–8848. [Google Scholar] [CrossRef] [Green Version]
- Gomes, M.S.; Flórido, M.; Pais, T.F.; Appelberg, R. Improved clearance of Mycobacterium avium upon disruption of the inducible nitric oxide synthase gene. J. Immunol. 1999, 162, 6734–6739. [Google Scholar]
- Flannagan, R.S.; Cosío, G.; Grinstein, S. Antimicrobial mechanisms of phagocytes and bacterial evasion strategies. Nat. Rev. Microbiol. 2009, 7, 355–366. [Google Scholar] [CrossRef]
- Cambier, C.J.; Takaki, K.K.; Larson, R.P.; Hernandez, R.E.; Tobin, D.M.; Urdahl, K.B.; Cosma, C.L.; Ramakrishnan, L. Mycobacteria manipulate macrophage recruitment through coordinated use of membrane lipids. Nature 2014, 505, 218–222. [Google Scholar] [CrossRef] [Green Version]
- Segal, B.H.; Doherty, T.M.; Wynn, T.A.; Cheever, A.W.; Sher, A.; Holland, S.M. The p47(phox-/-) mouse model of chronic granulomatous disease has normal granuloma formation and cytokine responses to Mycobacterium avium and Schistosoma mansoni eggs. Infect. Immun. 1999, 67, 1659–1665. [Google Scholar] [CrossRef]
- Mayer, B.K.; Falkinham, J.O. Superoxide dismutase activity of Mycobacterium avium, M. intracellulare, and M. scrofulaceum. Infect. Immun. 1986, 53, 631–635. [Google Scholar] [CrossRef] [Green Version]
- Escuyer, V.; Haddad, N.; Frehel, C.; Berche, P. Molecular characterization of a surface-exposed superoxide dismutase of Mycobacterium avium. Microb. Pathog. 1996, 20, 41–55. [Google Scholar] [CrossRef]
- Storz, G.; Imlay, J.A. Oxidative stress. Curr. Opin. Microbiol. 1999, 2, 188–194. [Google Scholar] [CrossRef]
- Cole, S.T.; Eiglmeier, K.; Parkhill, J.; James, K.D.; Thomson, N.R.; Wheeler, P.R.; Honoré, N.; Garnier, T.; Churcher, C.; Harris, D.; et al. Massive gene decay in the leprosy bacillus. Nature 2001, 409, 1007–1011. [Google Scholar] [CrossRef]
- Christman, M.F.; Storz, G.; Ames, B.N. OxyR, a positive regulator of hydrogen peroxide-inducible genes in Escherichia coli and Salmonella typhimurium, is homologous to a family of bacterial regulatory proteins. Proc. Natl. Acad. Sci. USA 1989, 86, 3484–3488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geier, H.; Mostowy, S.; Cangelosi, G.A.; Behr, M.A.; Ford, T.E. Autoinducer-2 triggers the oxidative stress response in Mycobacterium avium, leading to biofilm formation. Appl. Environ. Microbiol. 2008, 74, 1798–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reingewertz, T.H.; Meyer, T.; McIntosh, F.; Sullivan, J.; Meir, M.; Chang, Y.F.; Behr, M.A.; Barkan, D. Differential Sensitivity of Mycobacteria to Isoniazid Is Related to Differences in KatG-Mediated Enzymatic Activation of the Drug. Antimicrob. Agents Chemother. 2020, 64, e01899-19. [Google Scholar] [CrossRef]
- Ng, V.H.; Cox, J.S.; Sousa, A.O.; MacMicking, J.D.; McKinney, J.D. Role of KatG catalase-peroxidase in mycobacterial pathogenesis: Countering the phagocyte oxidative burst. Mol. Microbiol. 2004, 52, 1291–1302. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; De Buck, J. Localization of proteins in the cell wall of Mycobacterium avium subsp. paratuberculosis K10 by proteomic analysis. Proteome Sci. 2010, 8, 21. [Google Scholar] [CrossRef] [Green Version]
- Seyler, R.W.; Olson, J.W.; Maier, R.J. Superoxide dismutase-deficient mutants of Helicobacter pylori are hypersensitive to oxidative stress and defective in host colonization. Infect. Immun. 2001, 69, 4034–4040. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.C.; DeGroote, M.A.; Foster, J.W.; Bäumler, A.J.; Ochsner, U.; Testerman, T.; Bearson, S.; Giárd, J.C.; Xu, Y.; Campbell, G.; et al. Virulent Salmonella typhimurium has two periplasmic Cu, Zn-superoxide dismutases. Proc. Natl. Acad. Sci. USA 1999, 96, 7502–7507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igwe, E.I.; Rüssmann, H.; Roggenkamp, A.; Noll, A.; Autenrieth, I.B.; Heesemann, J. Rational live oral carrier vaccine design by mutating virulence-associated genes of Yersinia enterocolitica. Infect. Immun. 1999, 67, 5500–5507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, G.M.; Freeman, B.A. Detection of superoxide generated by endothelial cells. Proc. Natl. Acad. Sci. USA 1984, 81, 7269–7273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrt, S.; Schnappinger, D. Mycobacterial survival strategies in the phagosome: Defense against host stresses. Cell. Microbiol. 2009, 11, 1170–1178. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.H.; Tsai-Wu, J.J.; Huang, Y.T.; Lin, C.Y.; Lioua, G.G.; Lee, F.J. Identification and subcellular localization of a novel Cu,Zn superoxide dismutase of Mycobacterium tuberculosis. FEBS Lett. 1998, 439, 192–196. [Google Scholar] [CrossRef] [Green Version]
- Piddington, D.L.; Fang, F.C.; Laessig, T.; Cooper, A.M.; Orme, I.M.; Buchmeier, N.A. Cu,Zn superoxide dismutase of Mycobacterium tuberculosis contributes to survival in activated macrophages that are generating an oxidative burst. Infect. Immun. 2001, 69, 4980–4987. [Google Scholar] [CrossRef] [Green Version]
- Sassetti, C.M.; Rubin, E.J. Genetic requirements for mycobacterial survival during infection. Proc. Natl. Acad. Sci. USA 2003, 100, 12989–12994. [Google Scholar] [CrossRef] [Green Version]
- Bryk, R.; Lima, C.D.; Erdjument-Bromage, H.; Tempst, P.; Nathan, C. Metabolic enzymes of mycobacteria linked to antioxidant defense by a thioredoxin-like protein. Science 2002, 295, 1073–1077. [Google Scholar] [CrossRef]
- Doi, T.; Ando, M.; Akaike, T.; Suga, M.; Sato, K.; Maeda, H. Resistance to nitric oxide in Mycobacterium avium complex and its implication in pathogenesis. Infect. Immun. 1993, 61, 1980–1989. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.C. Antimicrobial reactive oxygen and nitrogen species: Concepts and controversies. Nat. Rev. Microbiol. 2004, 2, 820–832. [Google Scholar] [CrossRef] [PubMed]
- Doherty, T.M.; Sher, A. Defects in cell-mediated immunity affect chronic, but not innate, resistance of mice to Mycobacterium avium infection. J. Immunol. 1997, 158, 4822–4831. [Google Scholar] [PubMed]
- Ehlers, S.; Kutsch, S.; Benini, J.; Cooper, A.; Hahn, C.; Gerdes, J.; Orme, I.; Martin, C.; Rietschel, E.T. NOS2-derived nitric oxide regulates the size, quantity and quality of granuloma formation in Mycobacterium avium-infected mice without affecting bacterial loads. Immunology 1999, 98, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Orme, I.M.; Ordway, D.J. Host response to nontuberculous mycobacterial infections of current clinical importance. Infect. Immun. 2014, 82, 3516–3522. [Google Scholar] [CrossRef] [Green Version]
- Jamaati, H.; Mortaz, E.; Pajouhi, Z.; Folkerts, G.; Movassaghi, M.; Moloudizargari, M.; Adcock, I.M.; Garssen, J. Nitric Oxide in the Pathogenesis and Treatment of Tuberculosis. Front. Microbiol. 2017, 8, 2008. [Google Scholar] [CrossRef] [Green Version]
- Voskuil, M.I.; Schnappinger, D.; Visconti, K.C.; Harrell, M.I.; Dolganov, G.M.; Sherman, D.R.; Schoolnik, G.K. Inhibition of respiration by nitric oxide induces a Mycobacterium tuberculosis dormancy program. J. Exp. Med. 2003, 198, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Archuleta, R.J.; Yvonne Hoppes, P.; Primm, T.P. Mycobacterium avium enters a state of metabolic dormancy in response to starvation. Tuberculosis 2005, 85, 147–158. [Google Scholar] [CrossRef]
- Tan, M.P.; Sequeira, P.; Lin, W.W.; Phong, W.Y.; Cliff, P.; Ng, S.H.; Lee, B.H.; Camacho, L.; Schnappinger, D.; Ehrt, S.; et al. Nitrate respiration protects hypoxic Mycobacterium tuberculosis against acid- and reactive nitrogen species stresses. PLoS ONE 2010, 5, e13356. [Google Scholar] [CrossRef] [Green Version]
- Cunningham-Bussel, A.; Zhang, T.; Nathan, C.F. Nitrite produced by Mycobacterium tuberculosis in human macrophages in physiologic oxygen impacts bacterial ATP consumption and gene expression. Proc. Natl. Acad. Sci. USA 2013, 110, E4256–E4265. [Google Scholar] [CrossRef] [Green Version]
- Butala, N.S.; Falkinham, J.O. Nitrate and nitrite reductase activities of Mycobacterium avium. Int. J. Mycobacteriol. 2018, 7, 328–331. [Google Scholar] [CrossRef]
- Pathania, R.; Navani, N.K.; Gardner, A.M.; Gardner, P.R.; Dikshit, K.L. Nitric oxide scavenging and detoxification by the Mycobacterium tuberculosis haemoglobin, HbN in Escherichia coli. Mol. Microbiol. 2002, 45, 1303–1314. [Google Scholar] [CrossRef]
- Wittenberg, J.B.; Bolognesi, M.; Wittenberg, B.A.; Guertin, M. Truncated hemoglobins: A new family of hemoglobins widely distributed in bacteria, unicellular eukaryotes, and plants. J. Biol. Chem. 2002, 277, 871–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conus, S.; Simon, H.U. Cathepsins: Key modulators of cell death and inflammatory responses. Biochem. Pharmacol. 2008, 76, 1374–1382. [Google Scholar] [CrossRef] [PubMed]
- Garin, J.; Diez, R.; Kieffer, S.; Dermine, J.F.; Duclos, S.; Gagnon, E.; Sadoul, R.; Rondeau, C.; Desjardins, M. The phagosome proteome: Insight into phagosome functions. J. Cell Biol. 2001, 152, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Danelishvili, L.; Wagner, D.; Petrofsky, M.; Bermudez, L.E. Identification of virulence determinants of Mycobacterium avium that impact on the ability to resist host killing mechanisms. J. Med. Microbiol. 2010, 59 Pt 1, 8–16. [Google Scholar] [CrossRef]
- Thiriot, J.D.; Martinez-Martinez, Y.B.; Endsley, J.J.; Torres, A.G. Hacking the host: Exploitation of macrophage polarization by intracellular bacterial pathogens. Pathog. Dis. 2020, 78, ftaa009. [Google Scholar] [CrossRef] [Green Version]
- Tapels, D.; Hill, P.; Westermann, A.J.; Fisher, R.A.; Thurston, T.L.; Saliba, A.E.; Blommestein, I.; Vogel, J.; Helaine, S. Salmonella persisters undermine host immune defenses during antibiotic treatment. Science 2018, 362, 1156–1160. [Google Scholar] [CrossRef] [Green Version]
- Danelishvili, L.; Armstrong, E.; Miyasako, E.; Jeffrey, B.; Bermudez, L.E. Exposure of Mycobacterium avium subsp. homonissuis to Metal Concentrations of the Phagosome Environment Enhances the Selection of Persistent Subpopulation to Antibiotic Treatment. Antibiotics 2020, 9, 927. [Google Scholar] [CrossRef]
- Winthrop, K.L.; Chang, E.; Yamashita, S.; Iademarco, M.F.; LoBue, P.A. Nontuberculous mycobacteria infection and tumor necrosis factor-alpha antagonists. Emerg. Infect. Dis. 2009, 15, 1700–1701. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Description | Function in M. avium | Virulent Mechanisms | Reference |
|---|---|---|---|---|
| MAV_3616 | Long-chain acyl-CoA dehydrogenase | Catabolism of fatty acid and amino acids | Antimicrobial peptides resistance. | [36] |
| MAV_2941 | Hypothetical protein | Hijacking host binding protein | Inhibition of phagosome-lysosome maturation by mimicking the binding site of host vesicle trafficking proteins. | [8] |
| MAV_2928 | PPE25_MAV | Secreted protein, type VII secretion system | Inhibition of phagosome-lysosome maturation. | [52,56] |
| MAV_2839 | Alkylhydroperoxide reductase | Catalyzes peroxide reduction | Resistance to oxidative stress, such as reactive oxygen intermediates. | [57] |
| MAV_4682 | isocitrate lyase | Metabolism, glyoxylate shut | Resistance to oxidative stress. | [57] |
| MAV_2043 | Cu-Zn-SOD | Catalysis, superoxide dismutase | Resistance to oxidative stress phagosome acidification. | [58] |
| MAV_4264 | Hypothetical protein | Unknown function, homology with bacterial regulatory protein TetR | Resistance to both oxidative stress and phagosome acidification. | [59] |
| MAV_4644 | ADP-ribosyltransferase | Interfering with host cathepsin Z protein | Resistance to oxidative stress, such as nitric oxide. Resistance to phagosome acidification | [9] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abukhalid, N.; Islam, S.; Ndzeidze, R.; Bermudez, L.E. Mycobacterium avium Subsp. hominissuis Interactions with Macrophage Killing Mechanisms. Pathogens 2021, 10, 1365. https://doi.org/10.3390/pathogens10111365
Abukhalid N, Islam S, Ndzeidze R, Bermudez LE. Mycobacterium avium Subsp. hominissuis Interactions with Macrophage Killing Mechanisms. Pathogens. 2021; 10(11):1365. https://doi.org/10.3390/pathogens10111365
Chicago/Turabian StyleAbukhalid, Norah, Sabrina Islam, Robert Ndzeidze, and Luiz E. Bermudez. 2021. "Mycobacterium avium Subsp. hominissuis Interactions with Macrophage Killing Mechanisms" Pathogens 10, no. 11: 1365. https://doi.org/10.3390/pathogens10111365
APA StyleAbukhalid, N., Islam, S., Ndzeidze, R., & Bermudez, L. E. (2021). Mycobacterium avium Subsp. hominissuis Interactions with Macrophage Killing Mechanisms. Pathogens, 10(11), 1365. https://doi.org/10.3390/pathogens10111365