
Biosorption and Bioaccumulation Capacity of Arthrospira platensis toward Yttrium Ions


 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Object of Study
2.3. Biosorption
2.4. Bioaccumulation
2.5. Biochemical Tests
2.6. Antioxidant Activity
2.7. Statistical Analysis
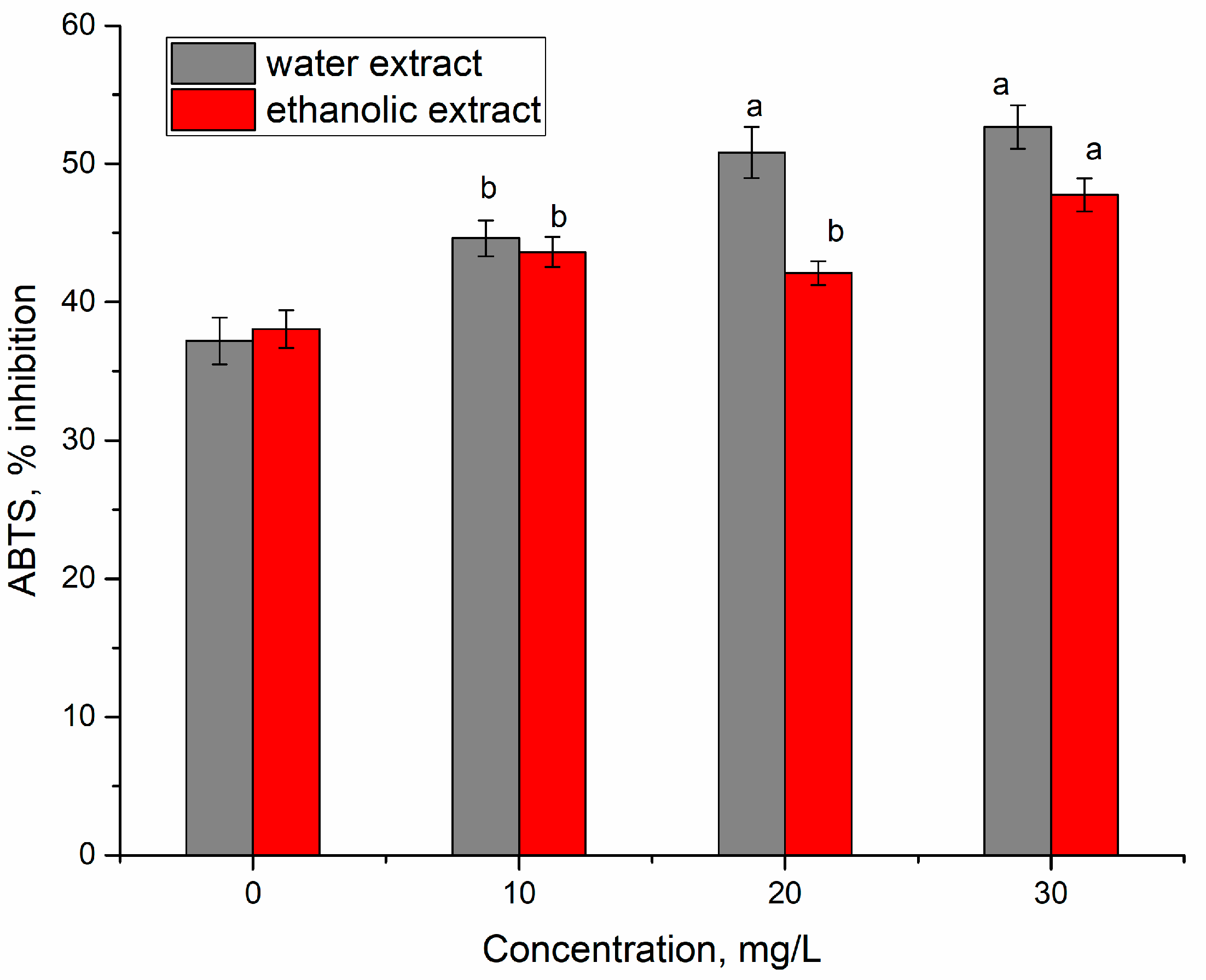
3. Results and Discussion
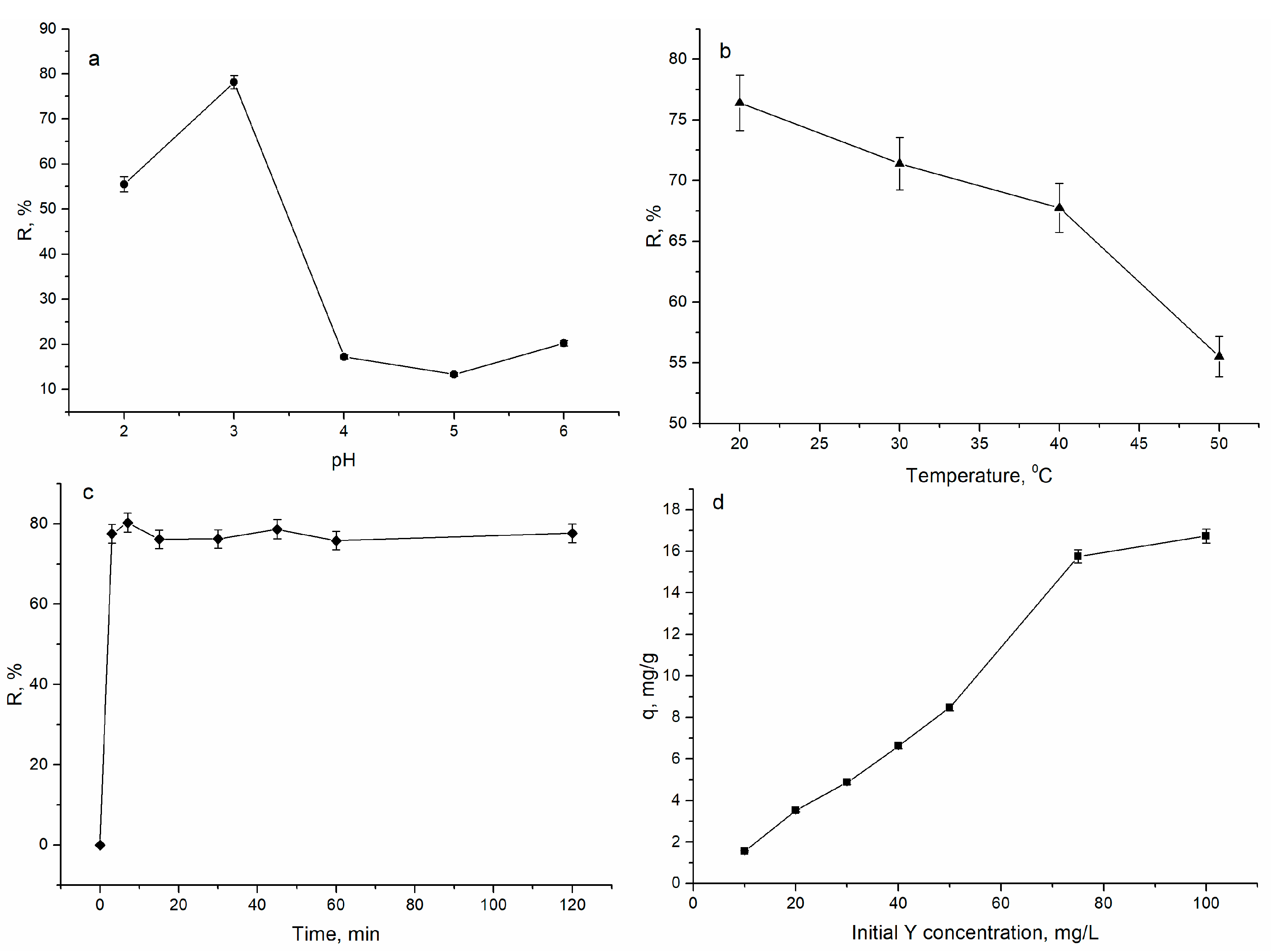
3.1. Yttrium Removal by Biosorption
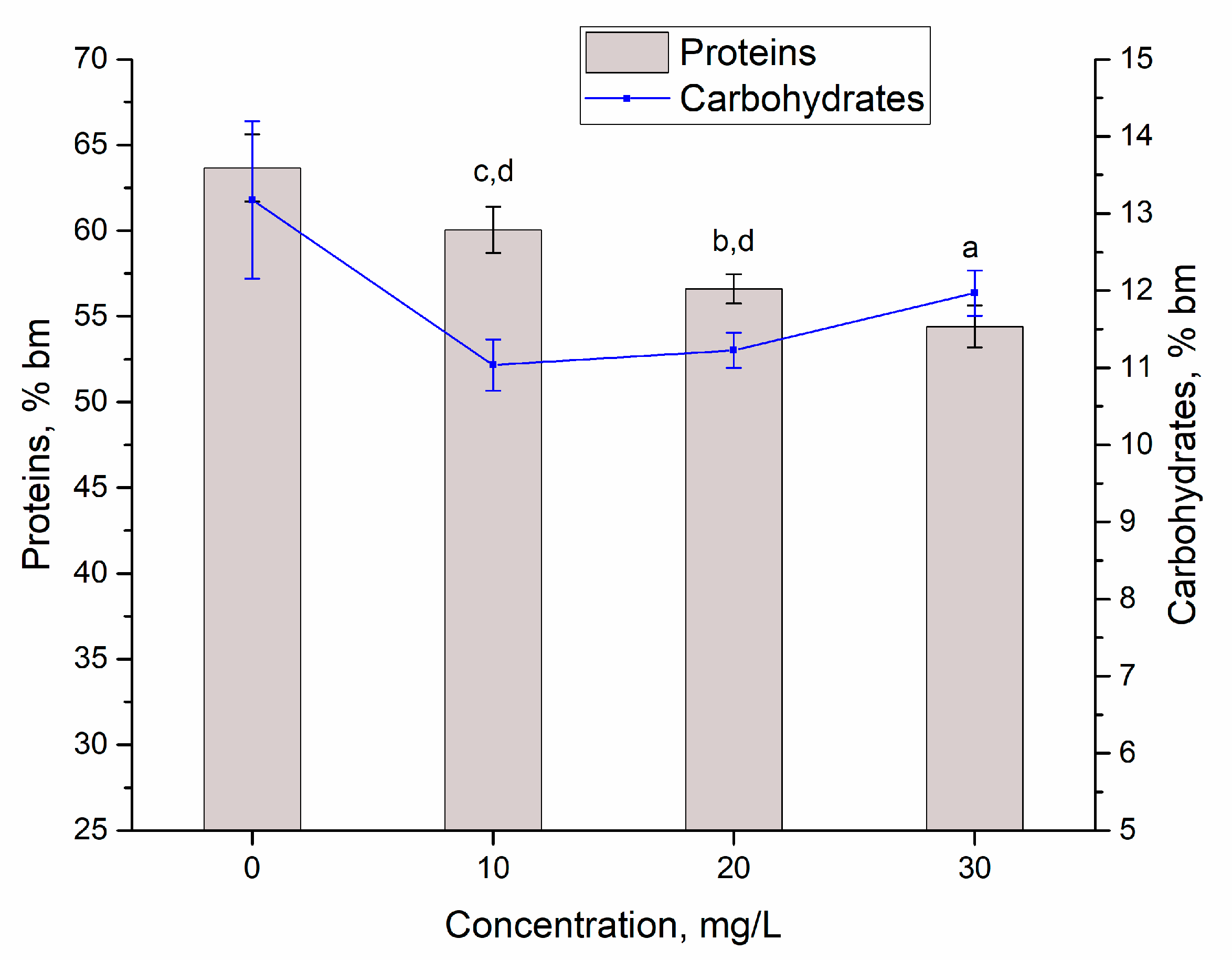
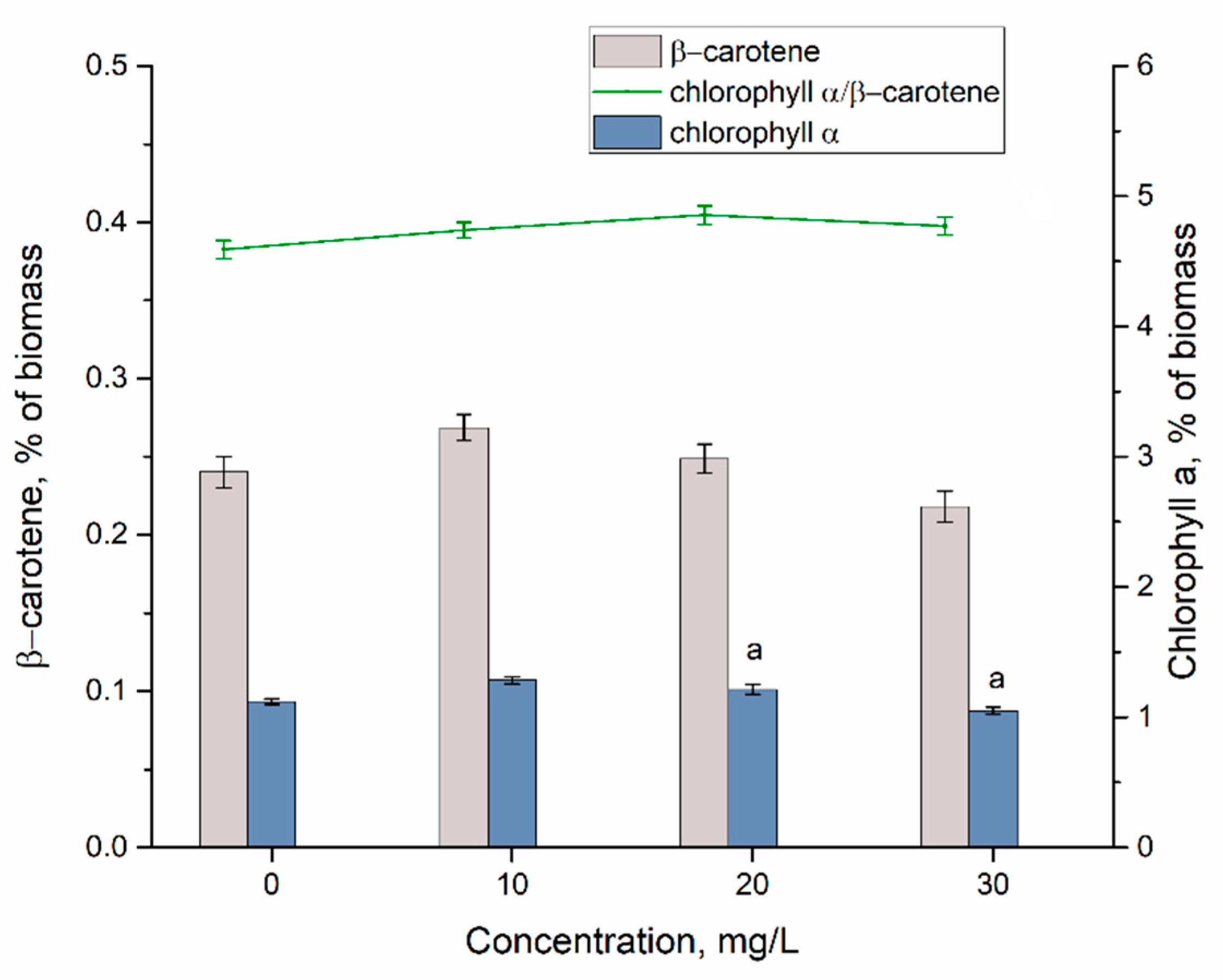
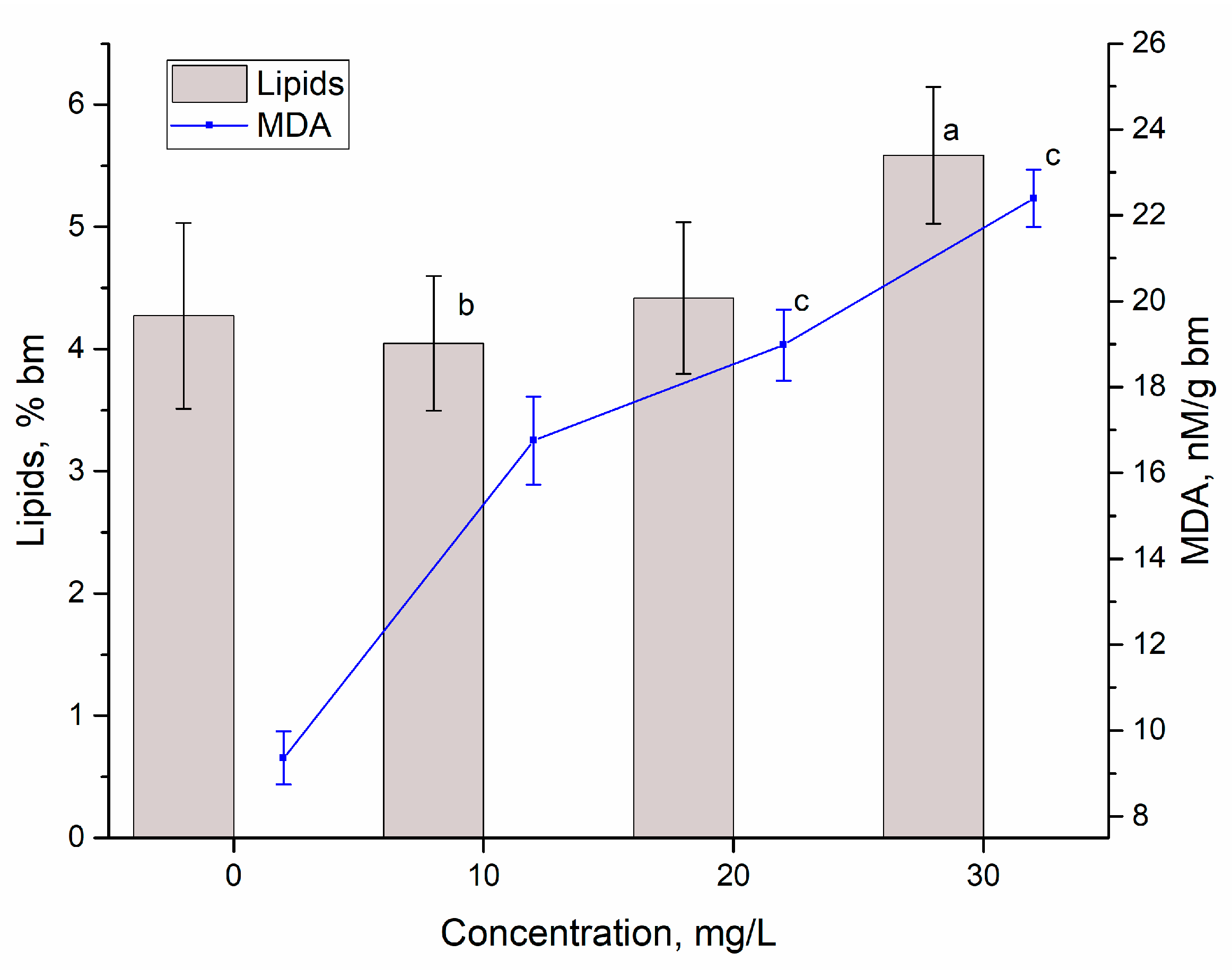
3.2. Yttrium Removal by Bioaccumulation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Wang, W.; Xu, C.; Jin, Y.; Zhang, Z.; Yan, R.; Zhu, D. The accumulation of rare-earth yttrium ions by Penicillium sp. ZD28. AMB Express 2020, 10, 25. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.L.; Shen, J.L. Removal of yttrium from rare-earth wastewater by Serratia marcescens: Biosorption optimization and mechanisms studies. Sci. Rep. 2022, 12, 4861. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Graedel, T.E. Global in-use stocks of the rare earth elements: A first estimate. Environ. Sci. Technol. 2011, 45, 4096–4101. [Google Scholar] [CrossRef] [PubMed]
- Dubey, S.S.; Grandhi, S. Sorption studies of yttrium(III) ions on surfaces of nano-thorium(IV) oxide and nano-zirconium(IV) oxide. Int. J. Environ. Sci. Technol. 2019, 16, 59–70. [Google Scholar] [CrossRef]
- Cardon, P.Y.; Triffault-Bouchet, G.; Caron, A.; Rosabal, M.; Fortin, C.; Amyot, M. Toxicity and Subcellular Fractionation of Yttrium in Three Freshwater Organisms: Daphnia magna, Chironomus riparius, and Oncorhynchus mykiss. ACS Omega 2019, 4, 13747–13755. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Tian, Y.; Zeng, Z.; Shuai, P.; Lan, H.; Zhu, X.; Zhong, Y.; Wu, L.; Fan, X. YCl3 promotes neuronal cell death by inducing apoptotic pathways in rats. Biomed Res. Int. 2017, 2017, 2183658. [Google Scholar] [CrossRef]
- Nakamura, Y.; Tsumura, Y.; Tonogai, Y.; Shibata, T.; Ito, Y. Differences in Behavior among the Chlorides of Seven Rare Earth Elements Administered Intravenously to Rats. Toxicol. Sci. 1997, 37, 106–116. [Google Scholar] [CrossRef]
- Hamdan, A.M.; Abd-El-Mageed, H.; Ghanem, N. Biological treatment of hazardous heavy metals by Streptomyces rochei ANH for sustainable water management in agriculture. Sci. Rep. 2021, 11, 9314. [Google Scholar] [CrossRef]
- Razzak, S.A.; Faruque, M.O.; Alsheikh, Z.; Alsheikhmohamad, L.; Alkuroud, D.; Alfayez, A.; Hossain, S.M.Z.; Hossain, M.M. A comprehensive review on conventional and biological-driven heavy metals removal from industrial wastewater. Environ. Adv. 2022, 7, 100168. [Google Scholar] [CrossRef]
- Yushin, N.; Zinicovscaia, I.; Cepoi, L.; Chiriac, T.; Rudi, L.; Grozdov, D. Biosorption and Bioaccumulation Capacity of Arthospiraplatensis toward Europium Ions. Water 2022, 14, 2128. [Google Scholar] [CrossRef]
- Cepoi, L. Environmental and Technological Stresses and Their Management in Cyanobacteria. In Cyanobacteria: From Basic Science to Applications; Elsevier: Amsterdam, The Netherlands, 2018; pp. 217–244. ISBN 9780128146682. [Google Scholar]
- Cepoi, L.; Zinicovscaia, I.; Rudi, L.; Chiriac, T.; Miscu, V.; Djur, S.; Strelkova, L.; Grozdov, D. Spirulina platensis as renewable accumulator for heavy metals accumulation from multi-element synthetic effluents. Environ. Sci. Pollut. Res. 2020, 27, 31793–31811. [Google Scholar] [CrossRef] [PubMed]
- Cepoi, L.; Zinicovscaia, I.; Rudi, L.; Chiriac, T.; Miscu, V.; Djur, S.; Strelkova, L.; Vergel, K.; Nekhoroshkov, P. Growth and heavy metals accumulation by Spirulina platensis biomass from multicomponent copper containing synthetic effluents during repeated cultivation cycles. Ecol. Eng. 2020, 142, 105637. [Google Scholar] [CrossRef]
- Gunasundari, E.; Kumar, P.S. Higher adsorption capacity of Spirulina platensis alga for Cr(VI) ions removal: Parameter optimisation, equilibrium, kinetic and thermodynamic predictions. IET Nanobiotechnol. 2017, 11, 317–328. [Google Scholar] [CrossRef]
- Zinicovscaia, I.; Safonov, A.; Ostalkevich, S.; Gundorina, S.; Nekhoroshkov, P.; Grozdov, D. Metal ions removal from different type of industrial effluents using Spirulina platensis biomass. Int. J. Phytoremediation 2019, 21, 1442–1448. [Google Scholar] [CrossRef] [PubMed]
- Almomani, F.; Bohsale, R.R. Bio-sorption of toxic metals from industrial wastewater by algae strains Spirulina platensis and Chlorella vulgaris: Application of isotherm, kinetic models and process optimization. Sci. Total Environ. 2021, 755, 142654. [Google Scholar] [CrossRef]
- Cepoi, L.; Zinicovscaia, I. Spirulina platensis as a model object for the environment bioremediation studies. In Handbook of Algal Science, Technology and Medicine; Academic Press: Cambridge, MA, USA, 2020; pp. 629–640. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Park, J.; Jeong, H.J.; Yoon, E.Y.; Moon, S.J. Easy and rapid quantification of lipid contents of marine dinoflagellates using the sulpho-phospho-vanillin method. Algae 2016, 31, 391–401. [Google Scholar] [CrossRef]
- Carra, P.O. Algal biliproteins. Biochem. J. 1970, 119, 2P–3P. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Fialkovsky, I.S.; Lutskiy, D.S.; Litvinova, T.E.; Oleijnik, O.I. Effect of anion composition on the extraction of cerium (III) and yttrium (III) by oleic acid. ARPN J. Eng. Appl. Sci. 2018, 13, 3152–3161. [Google Scholar]
- Kazak, E.S.; Kalitina, E.G.; Kharitonova, N.A.; Chelnokov, G.A.; Elovskii, E.V.; Bragin, I.V. Biosorption of Rare-Earth Elements and Yttrium by Heterotrophic Bacteria in an Aqueous Environment. Moscow Univ. Geol. Bull. 2018, 73, 287–294. [Google Scholar] [CrossRef]
- Karavaiko, G.I.; Kareva, A.S.; Avakian, Z.A.; Zakharova, V.I.; Korenevsky, A.A. Biosorption of scandium and yttrium from solutions. Biotechnol. Lett. 1996, 18, 1291–1296. [Google Scholar] [CrossRef]
- Umar, A.; Khan, M.S.; Alam, S.; Zekker, I.; Burlakovs, J.; dC Rubin, S.S.; Bhowmick, G.D.; Kallistova, A.; Pimenov, N.; Zahoor, M. Article synthesis and characterization of Pd-Ni bimetallic nanoparticles as efficient adsorbent for the removal of acid orange 8 present in wastewater. Water 2021, 13, 1095. [Google Scholar] [CrossRef]
- Rahman, N.U.; Ullah, I.; Alam, S.; Khan, M.S.; Shah, L.A.; Zekker, I.; Burlakovs, J.; Kallistova, A.; Pimenov, N.; Vincevica-Gaile, Z.; et al. Activated ailanthus altissima sawdust as adsorbent for removal of acid yellow 29 from wastewater: Kinetics approach. Water 2021, 13, 2136. [Google Scholar] [CrossRef]
- Treto-Suárez, M.A.; Prieto-García, J.O.; Mollineda-Trujillo, Á.; Lamazares, E.; Hidalgo-Rosa, Y.; Mena-Ulecia, K. Kinetic study of removal heavy metal from aqueous solution using the synthetic aluminum silicate. Sci. Rep. 2020, 10, 10836. [Google Scholar] [CrossRef]
- Nithya, R.; Gomathi, T.; Sudha, P.N.; Venkatesan, J.; Anil, S.; Kim, S.K. Removal of Cr(VI) from aqueous solution using chitosan-g-poly(butyl acrylate)/silica gel nanocomposite. Int. J. Biol. Macromol. 2016, 87, 545–554. [Google Scholar] [CrossRef]
- Zinicovscaia, I.; Yushin, N.; Abdusamadzoda, D.; Grozdov, D.; Shvetsova, M. Efficient Removal of Metals from Synthetic and Real Galvanic Zinc–Containing Effluents by Brewer’s Yeast Saccharomyces cerevisiae. Materials 2020, 13, 3624. [Google Scholar] [CrossRef]
- Liu, J.; Zeng, L.; Liao, S.; Liao, X.; Liu, J.; Mao, J.; Chen, Y.; Qiu, T.; Ren, S. Highly efficient enrichment and adsorption of rare earth ions (yttrium(III)) by recyclable magnetic nitrogen functionalized mesoporous expanded perlite. Chin. Chem. Lett. 2020, 31, 2849–2853. [Google Scholar] [CrossRef]
- Sakr, A.K.; Cheira, M.F.; Hassanin, M.A.; Mira, H.I.; Mohamed, S.A.; Khandaker, M.U.; Osman, H.; Eed, E.M.; Sayyed, M.I.; Hanfi, M.Y. Adsorption of yttrium ions on 3-amino-5-hydroxypyrazole impregnated bleaching clay, a novel sorbent material. Appl. Sci. 2021, 11, 10320. [Google Scholar] [CrossRef]
- Li, Y.; Xia, B.; Zhao, Q.; Liu, F.; Zhang, P.; Du, Q.; Wang, D.; Li, D.; Wang, Z.; Xia, Y. Removal of copper ions from aqueous solution by calcium alginate immobilized kaolin. J. Environ. Sci. 2011, 23, 404–411. [Google Scholar] [CrossRef]
- He, C.; Salih, K.A.M.; Wei, Y.; Mira, H.; Abdel-Rahman, A.A.H.; Elwakeel, K.Z.; Hamza, M.F.; Guibal, E. Efficient recovery of rare earth elements (Pr(iii) and tm(iii)) from mining residues using a new phosphorylated hydrogel (algal biomass/pei). Metals 2021, 11, 294. [Google Scholar] [CrossRef]
- Moldoveanu, G.A.; Papangelakis, V.G. Recovery of rare earth elements adsorbed on clay minerals: I. Desorption mechanism. Hydrometallurgy 2012, 117–118, 71–78. [Google Scholar] [CrossRef]
- Pinto, J.; Henriques, B.; Soares, J.; Costa, M.; Dias, M.; Fabre, E.; Lopes, C.B.; Vale, C.; Pinheiro-Torres, J.; Pereira, E. A green method based on living macroalgae for the removal of rare-earth elements from contaminated waters. J. Environ. Manag. 2020, 263, 110376. [Google Scholar] [CrossRef]
- Zinicovscaia, I.; Cepoi, L.; Rudi, L.; Chiriac, T.; Grozdov, D.; Pavlov, S.; Djur, S. Accumulation of dysprosium, samarium, terbium, lanthanum, neodymium and ytterbium by Arthrospira platensis and their effects on biomass biochemical composition. J. Rare Earths 2021, 39, 1133–1143. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Luo, X.; Ren, Y.; Gao, E.; Gao, H. Effects of yttrium and phosphorus on growth and physiological characteristics of Microcystis aeruginosa. J. Rare Earths 2018, 36, 781–788. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Y.; Du, J.; Wang, Z.; Wu, Q. Effects of yttrium under lead stress on growth and physiological characteristics of Microcystis aeruginosa. J. Rare Earths 2016, 34, 747–756. [Google Scholar] [CrossRef]
- Jin, X.; Chu, Z.; Yan, F.; Zeng, Q. Effects of lanthanum(III) and EDTA on the growth and competition of Microcystis aeruginosa and Scenedesmus quadricauda. Limnologica 2009, 39, 86–93. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.; Lü, Y.; Jin, H.; Deng, S.; Zeng, Y. Effects of cerium on growth and physiological characteristics of Anabaena flosaquae. J. Rare Earths 2012, 30, 1287–1292. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, H.; Deng, S.; Chen, Y.; Yu, Y. Effects of neodymium on growth and physiological characteristics of Microcystis aeruginosa. J. Rare Earths 2011, 29, 388–395. [Google Scholar] [CrossRef]
- Sekar, S.; Chandramohan, M. Phycobiliproteins as a commodity: Trends in applied research, patents and commercialization. J. Appl. Phycol. 2008, 20, 113–136. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kinetics | Model | Parameters | Y(III) | Isotherms | Model | Parameters | Y(III) |
| Pseudo-First-Order | qe | 1.53 | Langmuir | qm | 719.8 | ||
| k1 | 1.79 | b | 0.0002 | ||||
| R2 | 0.99 | ||||||
| SSE | 0.006 | R2 | 0.97 | ||||
| χ2 | 0.003 | ||||||
| Pseudo-Second-Order | qe | 1.5 | Freundlich | KF | 0.17 | ||
| k2 | –2.54 | ||||||
| R2 | 0.99 | n | 0.99 | ||||
| SSE | 0.006 | ||||||
| χ2 | 0.003 | R2 | 0.97 | ||||
| Elovich | α | 1.15·1043 | Temkin | bT | 0.53 | ||
| β | 69.4 | ||||||
| R2 | 0.99 | aT | 0.2 | ||||
| SSE | 32286 | ||||||
| χ2 | 21054 | R2 | 0.75 |
| Sorbent | pH | qmax, mg/g | Reference |
| A. platensis | 3 | 719.8 | Present study |
| Serratia marcescens | 5.5 | 123.65 | [2] |
| Nano-thorium (IV) Oxide | 6.9 | 10.5 | [4] |
| Nano-zirconium (IV) Oxide | 6.9 | 18 | [4] |
| 3-Amino-5-Hydroxypyrazole Impregnated Bleaching Clay | 6.0 | 172.41 | [32] |
| Magnetic Nitrogen Functionalized Mesoporous Expanded Perlite | 5.5 | 383.2 | [31] |
| Temperature, K | ∆G°, kJ/mol | ∆H°, kJ/mol | ∆S°, J/mol·K |
|---|---|---|---|
| 293 | −12.1 | −2.1 | 34.1 |
| 303 | −12.5 | ||
| 313 | −12.8 | ||
| 323 | −13.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yushin, N.; Zinicovscaia, I.; Cepoi, L.; Chiriac, T.; Rudi, L.; Grozdov, D. Biosorption and Bioaccumulation Capacity of Arthrospira platensis toward Yttrium Ions. Metals 2022, 12, 1465. https://doi.org/10.3390/met12091465
Yushin N, Zinicovscaia I, Cepoi L, Chiriac T, Rudi L, Grozdov D. Biosorption and Bioaccumulation Capacity of Arthrospira platensis toward Yttrium Ions. Metals. 2022; 12(9):1465. https://doi.org/10.3390/met12091465
Chicago/Turabian StyleYushin, Nikita, Inga Zinicovscaia, Liliana Cepoi, Tatiana Chiriac, Ludmila Rudi, and Dmitrii Grozdov. 2022. "Biosorption and Bioaccumulation Capacity of Arthrospira platensis toward Yttrium Ions" Metals 12, no. 9: 1465. https://doi.org/10.3390/met12091465
APA StyleYushin, N., Zinicovscaia, I., Cepoi, L., Chiriac, T., Rudi, L., & Grozdov, D. (2022). Biosorption and Bioaccumulation Capacity of Arthrospira platensis toward Yttrium Ions. Metals, 12(9), 1465. https://doi.org/10.3390/met12091465